Нейрон ограничен липо-протеиновой(жиро-белковой) мембраной. Мембранный потенциал нейрона оказывает влияние на процессы трансмембранного обмена веществ. Мембранный потенциал покоящейся клетки называется потенциалом покоя.
Скорость диффузии веществ через мембрану зависит главным образом от размера молекул и их относительной растворимости в жирах.
В мембранах клеток существуют белки-транслоказы. Взаимодействуя со специфическим лигандом, они обеспечивают его диффузию (транспорт из области большей концентрации в область меньшей) через мембрану.
Специальные молекулярные рецепторы, находящиеся на мембране, могут соединяться с сигнальными (управляющими) веществами, вследствие чего может меняться состояние мембраны и всей клетки. Молекулярные рецепторы запускают биохимические реакции в ответ на соединение с ними лагандов (управляющих веществ).
каталитическая ферментативная активность. Ферменты могут быть встроены в мембрану или связаны с её поверхностью (как внутри, так и снаружи клетки), и там они осуществляют свою ферментативную деятельность.
Как видим, основное свойство мембраны – это её проницаемость по отношению к различным веществам.

Легче и лучше всего через мембрану пассивно проникают жирорастворимые неполярные мелкие молекулы. Так, легче всего с помощью простой диффузией проходят через мембрану малые неполярные молекулы, такие как О2, стероиды, тиреоидные гормоны, а также жирные кислоты. Несколько медленнее диффундируют через липидный слой малые полярные незаряженные молекулы: СО2, NH3, Н2О, этанол, мочевина. Диффузия глицерола идёт уже значительно медленнее, а глюкоза практически не способна самостоятельно пройти через мембрану. Для всех заряженных молекул, независимо от размера, липидная мембрана практически непроницаема. Таким образом, свободно проникать сквозь мембрану в клетку и обратно могут только жирорастворимые вещества, способные растворяться в жировом (липидном) слое мембраны. Транспорт других веществ через мембрану требует особых механизмов. Какие же вещества необходимо протаскивать через мембрану «насильно»? Это все полярные молекулы, не растворимые в жирах: молекулы воды, ионы (электролиты), а также более крупные молекулы питательных веществ, таких как глюкоза и аминокислоты.
Для транспорта в клетку веществ, слабо способных к диффузии через липидный слой мембраны, необходимы специальные транспортные структуры.
Виды транспортных структур мембраны:
1. Ионные каналы – специальные поры (дырочки) в мембране, образованные канальными белками, позволяющие ионам проходить через мембрану в обоих направлениях: как внутрь, так и наружу.
2. Транслоказы – специальные мембранные белки, облегчающие переход вещества через мембрану за счёт своего временного связывания с диффундирующим веществом. Не требуют энергии, работают в обоих направлениях в зависимости от концентрации переносимого вещества.
3. Транспортёры – белковые структуры, насильно протаскивающие определённые вещества сквозь клеточную мембрану в определённом направлении с затратами энергии. Ионные насосы – это транспортёры ионов. По способу использования энергии для своей работы транспортёры можно разделить на «симпортные» и «антипортные». Симпортные транспортёры используют совместный транспорт в одном направлении двух веществ: одно из них должно иметь большую потенциальную энергию для движения через мембрану. Например, симпорт в клетку с помощью ионов натрия глюкозы, или симпорт ионов кальция с помощью ионов натрия. Антипортные транспортёры (обменники) используют встречный транспорт двух веществ с разной потенциальной энергией диффузии. Так работает, например, натрий-калиевый ионный насос.
Итак, перенос веществ через клеточную мембрану происходит различными путями.
Механизмы транспорта веществ через мембрану:
1. Простая диффузия жирорастворимых (гидрофобных) веществ через жировой слой мембраны. Это пассивный процесс под действием градиента (перепада) концентрации вещества по разные стороны мембраны. (Смотрите видео: пассивный транспорт через мембрану).
2. Неуправляемая диффузия (неуправляемый пассивный перенос) водорастворимых веществ через постоянно открытые ионные каналы мембраны.
3. Управляемая диффузия (управляемый пассивный перенос) водорастворимых веществ через управляемые ионные каналы мембраны.
4. Активный транспорт водорастворимых веществ с помощью специальных белковых транспортных структур (транспортёров) за счёт использования энергии расщепления АТФ.
Одна из самых главных транспортных структур мембраны – это фермент АТФаза. АТФазы разных видов транспортируют через мембрану ионы. Они переносят их как внутрь клетки, так и, наоборот, наружу.
Название АТФаза означает, что это фермент, нацеленный на расщепление АТФ (аденозинтрифосфатаза)
6. Потенциал действия нейрона. Изменения возбудимости при возбуждении.
Потенциал действия - это электрофизиологический процесс, выражающийся в быстром колебании мембранного потенциала покоя вследствие перемещения ионов в клетку и из клетки и способный распространяться без затухания. ПД обеспечивает передачу сигналов между нервными клетками, между нервными центрами и рабочими органами.
Потенциал действия — это тот электрический импульс, которой доставляет информацию от рецепторов к мозгу, и с помощью которого мозг управляет телом. Для его существования необходимы особо устроенные клетки, нейроны, мембраны которых облеплены с обеих сторон положительно и отрицательно заряженными ионами, которые только и ждут сигнала, чтобы этот электрический импульс пронести по всей длине клетки. Основным местом существования потенциала действия является аксон нейрона. Дендриты некоторых типов нейронов также способны проводить электрический импульс.
Изменение возбудимости при возбуждении. При возбуждении возбудимость изменяется по фазам.
1) фаза первичной экзальтации - возбудимость выше нормы, реакция на порошковый и подпороговый раздражитель
2) фаза абсолютной рефрактерности - ответная реакция на раздражитель отсутствует, что обусловлено инактивацией натриевых каналов
3) фаза относительной рефрактерности - возбудимость восстанавливается, и ответная реакция становится возможной только при действии раздражителя надпороговой силы, что обусловлено выходящим калиевым током
4) фаза вторичной экзальтации - ответная реакция на подпороговый раздражитель
5) фаза субнормальной возбудимости - возбудимость ниже нормы, ответ возможен на действие надпороговой силы.
Восстановлением исходного положения «натрий снаружи, калий внутри» занимается натрий-калиевый насос, выводящий из клетки три Na+ и одновременно проносящий внутрь два К+.
Как видно, всё очень просто, главное – открыть нужные ионные каналы. Стимул-управляемые ионные каналы открываются раздражителем (стимулом). Хемо-управляемые ионные каналы открываются медиатором (возбуждающим или тормозным). Точнее, в зависимости от того, на какие каналы (натриевые, калиевые или хлорные) будет действовать медиатор, таков будет и локальный потенциал – возбуждающий или тормозный. А медиатор как для возбуждающих локальных потенциалов, так и для тормозных, может быть одним и тем же, тут важно, какие ионные каналы будут связываться с ним своими молекулярными рецепторами – натриевые, калиевые или хлорные.
7. Нервные волокна, их классификация. Особенности проведения возбуждения по безмиелиновым и миелинизированным волокнам.

Нервные волокна – это отростки нервных клеток, окруженные оболочками из нейроглиальных клеток.
По наличию (или отсутствию) миелиновой оболочки нервные волокна делят на миелиновые и безмиелиновые.
Классификация нервных волокон по Ллойду – Ханту учитывает диаметр волокна и скорость проведения импульса, но применяется только для чувствительных волокон.
Классификация нервных волокон по Эрлангеру – Гассеру основана на исследовании составного потенциала действия смешанного периферического нерва (например, седалищного).
Особенности распространения возбуждения по безмиелиновым волокнам:
1. Возбуждение распространяется непрерывно и все волокно сразу охватывается возбуждением.
2. Возбуждение распространяется с небольшой скоростью.
3. Возбуждение распространяется с декриментом (уменьшение силы тока к концу нервного волокна).
По безмиелиновым волокнам возбуждение проводится к внутренним органам от нервных центров.
Особенности распространения возбуждения по миелиновым волокнам:
1. Распространение ПД в миэлинизированных нервных волокнах осуществляется сальтаторно - скачкообразно от перехвата к перехвату, т.е. возбуждение (ПД) как бы «перепрыгивает» через участки нервного волокна, покрытые миелином, от одного перехвата к другому и все волокно сразу не охватывается возбуждением.
2. Возбуждение распространяется с большой скоростью.
По миелиновым волокнам возбуждение распространяется от анализаторов к ЦНС, к скелетным мышцам, т.е. там, где требуется высокая скорость ответной реакции.
ГЛАВНОЕ ОТЛИЧИЕ В ТОМ, ЧТО ПО МИЕЛИНОВЫМ ВОЛОКНАМ ПРОВОДИТСЯ НАМНОГО БЫСТРЕЕ.
Безмиелиновые нервные волокна - один слой швановских клеток, между ними - щелевидные пространства. Клеточная мембрана на всем протяжении контактирует с окружающей средой. При нанесении раздражения возбуждение возникает в месте действия раздражителя. Безмиелиновые нервные волокна обладают электрогенными свойствами (способностью генерировать нервные импульсы) на всем протяжении.
Миелиновые нервные волокна - покрыты слоями шванновских клеток, которые местами образуют перехваты Ранвье (участки без миелина) через каждые 1 мм. Продолжительность перехвата Ранвье 1 мкм. Миелиновая оболочка выполняет трофическую и изолирующую функции (высокое сопротивление). Участки, покрытые миелином не обладают электрогенными свойствами. Ими обладают перехваты Ранвье. Возбуждение возникает в ближайшем к месту действия раздражителя перехвата Ранвье. В перехватах Ранвье высокая плотность Nа-каналов, поэтому в каждом перехвате Ранвье происходит усиление нервных импульсов.
Перехваты Ранвье выполняют функцию ретрансляторов (генерируют и усиливают нервные импульсы).
8. Законы проведения возбуждения по нерву.
1)Возбуждение возможно в любом направлении
2) Проведения возбуждения возможно только по целому нерву (не поврежденному)
Закон изолированного проведения возбуждения по нервному волокну. В составе нерва возбуждение по нервному волокну распространяется изолированно, без перехода на другие волокна, имеющиеся в составе нерва.
Закон двустороннего проведения возбуждения по нервному волокну.Нервы обладают двусторонней проводимостью, т.е. возбуждение может распространяться в любом направлении от возбужденного участка (места его возникновения), т. е., центростремительно и центробежно. Это можно доказать, если на нервное волокно наложить регистрирующие электроды на некотором расстоянии друг от друга, а между ними нанести раздражение. Возбуждение зафиксируют электроды по обе стороны от места раздражения. Естественным направлением распространения возбуждения является: в афферентных проводниках - от рецептора к клетке, в эфферентных - от клетки к рабочему органу.
Закон анатомической и физиологической целостности нервного волокна.Проведение возбуждения по нервному волокну возможно лишь в том случае, если сохранена его анатомическая и физиологическая целостность, т.е. передача возбуждения возможна только по структурно и функционально не измененному, неповрежденному нерву (законы анатомической и физиологической целостности).
Закон изолированного проведения возбуждения по нервному волокну.В составе нерва возбуждение по нервному волокну распространяется изолированно, без перехода на другие волокна, имеющиеся в составе нерва. Изолированное проведение возбуждения обусловлено тем, что сопротивление жидкости, заполняющей межклеточные пространства, значительно ниже сопротивления мембраны нервных волокон. Поэтому основная часть тока, возникающего между возбужденным и невозбужденным участками нервного волокна, проходит по межклеточным щелям, не действуя на рядом расположенные нервные волокна. Изолированное проведение возбуждения имеет важное значение. Нерв содержит большое количество нервных волокон (чувствительных, двигательных, вегетативных), которые иннервируют различные по структуре и функциям эффекторы (клетки; ткани, органы). Если бы возбуждение внутри нерва распространялось с одного нервного волокна на другое, то нормальное функционирование органов было бы невозможно.
9. Синапсы. Центральные и периферические синапсы.
Си́напс - место контакта между двумя нейронами или между нейроном и получающей сигнал эффекторной клеткой. Периферические синапсы: нервно-мышечные
нейросекреторные (аксо-вазальные)
рецепторно-нейрональные
Центральные синапсы
аксо-дендритические — с дендритами, в том числе
аксо-шипиковые — с дендритными шипиками, выростами на дендритах;
аксо-соматические — с телами нейронов;
аксо-аксональные — между аксонами;
дендро-дендритические — между дендритами;
Центральные синапсы
В нервной системе синапсы образуются между отростками разных нейронов, а также между отростками и телами клеток. Соответственно их называют:
- Аксо-аксональными,
- Аксо-дендритными,
- Аксо-соматическими,
- Дендро-соматическими,
- Дендро-дендритными.
Количество синапсов на нейроне очень большое и достигает нескольких тысяч.
 Рис.3.4. Структура аксосоматического синапса.
Рис.3.4. Структура аксосоматического синапса.
В качестве примера может быть рассмотрен аксо-соматический синапс (между аксоном одной нервной клетки и телом другой), структура которого показана на рис.3.4. Аксон, подходя к телу другого нейрона, образует расширение, называемое пресинаптическим окончанием или терминалью. Мембрана такого окончания называется пресинаптической. Под ней располагается синаптическая щель, ширина которой составляет 10-50 мкм. За синаптической щелью лежит мембрана тела нейрона, называемая в области синапса постсинаптической.
1 — аксон,
2 — синаптическая пуговка,
3 — пресинаптическая мембрана,
4 — лостсинаптическая мембрана,
5 — рецепторы постсинаптической мембраны,
6 — синаптические пузырьки с медиатором,
7 — кванты медиатора в синаптической щели,
8 — митохондрии.
СаСБ — кальцийсвязывэющий белок.
Периферические синапсы
Периферические синапсы образованы между окончаниями эфферентного нерва и мембраной эффектора. Для соматических рефлексов это синапсы между двигательным нервом и скелетной мышцей, поэтому их еще называют нервно-мышечные синапсы (рис.3.7). Благодаря форме синаптического образования они получили название концевых пластинок. Строение и основные свойства концевых пластинок в общих чертах подобны центральным синапсам. Медиатором в нервно-мышечных синапсах служит ацетилхолин, выделяемый квантами. Разрушение медиатора в синаптической шели происходит за счет фермента холинэстеразы. Рецепторы постсинаптической мембраны относятся к никотиночувствительному типу (н-холинорепепторы), конкурентно блокируются ядом кураре, что прекращает нервно-мышечную передачу. Образуемый на постсинаптической мембране медиатор-рецепторный комплекс активирует хемо-чувствительные рецепторуправляемые Na-каналы, вызывает натриевый ток внутрь клетки, деполяризацию и формирование потенциала, называемого потенциалом концевой пластинки (ПКП), являющегося аналогом ВПСП центральных синапсов.
Важнейшим отличительным свойством нервно-мышечных синапсов от центральных является высокая амплитуда ПКП, всегда превышающая критический уровень деполяризации мембраны, поэтому ПКП не требует суммации как ВПСП, а каждый ПКП ведет к генерации потенциала действия, возбуждению и сокращению мышечной клетки.
 Рис.3.7. Структура нервно-мышечного синапса.
Рис.3.7. Структура нервно-мышечного синапса.
Возможно это связано с тем, что постсинаптическая мембрана имеет многочисленные складки, существенно увеличивающие поверхность взаимодействия медиатора с рецепторами (рис.3.7).
1 — пресинаптическая концевая пластинка,
2 — синаптические пузырьки с медиатором,
3 — митохондрии,
4 — синапгическая щель с квантами медиатора,
5 — складчатая постсинаптическая мембрана,
6 — рецепторы постсинаптической мембраны,
7 — саркоплазматический ретикулум,
8 — сократительный аппарат (миофибриллы) мышечной клетки.
10. Синапсы с электрическим механизмом передачи сигнала. Свойства.
Электрический синапс, или эфапс — электрический щелевой контакт между двумя примыкающими нейронами или иными возбудимыми клетками, которая образуется в виде узкой щели, содержащей характерные только для этого типа контактов элементы
Что такое электрический синапс? Электрические синапсы – это межклеточные образования, с помощью которых обеспечивается передача импульса возбуждения. Данный процесс происходит за счет появления электрического тока между двумя отделами под названием пресинаптический и постсинаптический.
Важная работа синапсов электрического типа Свойства электрических синапсов заключаются в следующем:
- быстрое действие (гораздо превосходит активность в синапсах химического типа); ----слабые следовые эффекты (суммации от последовательных импульсов практически нет);
-надежная передача возбуждения;
-высокая пластичность;
-передача в одну и в обе стороны.
Особенности структуры.

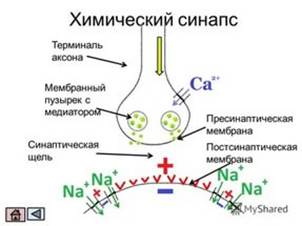
11. Синапсы с химическим механизмом передачи сигнала. Свойства.
Химический синапс — особый тип межклеточного контакта между нейроном и клеткой-мишенью. У данного типа синапса роль посредника (медиатора) передачи выполняет химическое вещество.
Синаптические контакты могут быть между аксоном и дендритом (аксодендритические), аксоном и сомой клетки (аксосоматические), аксонами (аксоаксональные), дендритами (дендродендритические), дендритами и сомой клетки.
. В покое медиатор попадает в синаптическую щель постоянно, но в малом количестве. Под влиянием пришедшего возбуждения количество медиатора резко возрастает. Затем медиатор перемещается к постсинаптической мембране, действует на специфические для него рецепторы и образует на мембране комплекс медиатор—рецептор. Данный комплекс изменяет проницаемость мембраны для ионов К+ и Na+, в результате чего изменяется ее потенциал покоя.