Коллоквиум: Филогенез систем органов. Регенерация. Гомеостаз.
Регенерация. Физиологическая регенерация, ее значение.
Важнейшая проблема медицины – восстановление поврежденных тканей и органов и возвращение им их функций. Проблема медицинская, но основа ее биологическая.
Регенерация – процесс вторичного развития органа или ткани, вызванный повреждениями какого – либо рода.
Первичное развитие – онтогенез.
Вторичное развитие – развитие, связанное не с естественным размножением, а с внешними воздействиями, но организм. Внешнее воздействие вовлекает дефинитивные органы и ткани в процесс развития. Дарвин подчеркивал, что половое размножение, бесполое размножение и регенерация – проявление одного и того же свойства организма.
Регенерация происходит на всех уровнях материи.
В процессе жизнедеятельности изменяется структура ДНК – молекулярная регенерация.
Регенерация может происходить внутри органоидов – внутриорганоидная регенерация. Восстанавливаются кристы митохондрий, цистерны комплекса Гольджи, части ЭПР и др. Например, гепатоцит человека, злоупотребляющего алкоголем.
Возможна регенерация целых органелл - органоидная. Восстанавливается число митохондрий, лизосом и других органоидов – гиперплазия.
Все вместе эти 3 уровня регенерации составляют внутриклеточную регенерацию.
Клеточная регенерация – увеличение количества клеток.
По способности к регенерации выделяют 3 группы тканей и органов:
1. Регенераторная реакция в форме новообразования клеток: эпителий кожи, костный мозг, костная ткань, эпителий тонкой кишки, лимфатическая система.
2. Промежуточная форма. Происходит деление клеток и внутриклеточная регенерация. Печень, легкие, почки, надпочечники, скелетная мускулатура.
3. Преобладает внутриклеточная регенерация. Клетки центральной нервной системы, миокарда.
Регенерация присуща всем организмам. С потерей или отсутствием способности к бесполому размножению теряется способность к соматической регенерации (из участка тела организм не образуется, но регенеративная функция отдельных частей организма сохраняется).
Регенерация может быть физиологической и репаративной. В свою очередь репаративная регенерация бывает нескольких видов:
- возмещающая;
- посттравматическая;
- восстановительная;
- патологическая.
По степени восстановления репаративная репарация может быть типической (полной) – гомоморфоз, морфолаксис и атипическая - неполная, гетероморфоз.
Физиологическая регенерация – восстановление частей организма, износившихся в процессе жизнедеятельности. Действует на протяжении всего онтогенеза, поддерживает постоянство структур, несмотря на гибель клеток. Интенсивные процессы физиологической регенерации при восстановлении клеток крови, эпидермиса, слизистых оболочек. Примерами могут быть линька птиц, рост зубов у грызунов. Физиологическая регенерация происходит не только в тканях с интенсивно делящимися клетками, но и там, где клетки делятся незначительно. 25 гепатоцитов из 1000 погибают и столько же восстанавливаются. Физиологическая регенерация – динамический процесс, который включает в себя клеточное деление и другие процессы. Обеспечение функций лежит в основе нормального функционирования организма.
Виды адаптации.
Все адаптации делят на аккомодации и эволюционные адаптации. Аккомодации представляют собой обратимый процесс. Они возникают при резком изменении условий среды. Например, при переселении животные попадают в новую для них обстановку, но постепенно привыкают к ней. Например, человек, переселившийся из средней полосы в тропики или на Крайний Север, некоторое время испытывает дискомфорт, но со временем привыкает к новым условиям. Эволюционная адаптация необратима и возникшие изменения генетически закрепляются. Сюда относят все приспособления, на которые действует естественный отбор. Например, покровительственная окраска или быстрый бег. Приспособления также делят на организменные и видовые. Организменные адаптации в свою очередь подразделяются на морфологические, физиологические, биохимические и этологические. Морфологические адаптации проявляются в преимуществах строения, покровительственной окраске, предостерегающей окраске, мимикрии, маскировке, приспособительном поведении. Преимущества строения – это оптимальные пропорции тела, расположение и густота волосяного или перьевого покрова и т.п. Хорошо известен облик водного млекопитающего – дельфина.
У животных, ведущих скрытный, затаивающийся образ жизни, полезным оказываются приспособления, придающие им сходство с предметами окружающей среды.
Покровительственная окраска позволяет быть незаметным среди окружающего фона.
Мимикрия (подражание) – это результат гомологичных (одинаковых) мутаций у разных видов, которые помогают выжить незащищённым животным.
Кроме защитной окраски, у животных и растений наблюдаются и другие средства защиты. У растений нередко образуются иглы и колючки, защищающие их от поедания травоядными животными (кактусы, шиповник, боярышник, облепиха и др.). Такую же роль играют ядовитые вещества, обжигающие волоски, например у крапивы.
Маскировка – приспособления, при которых форма тела и окраска животных сливаются с окружающими предметами. Например, в тропических лесах многие змеи неразличимы среди лиан, лохматый морской конёк похож на водоросль, насекомые на коре деревьев похожи на лишайники (жуки, усачи, пауки, бабочки).
Приспособительное поведение – принятие определённых поз покоя (гусеницы некоторых насекомых в неподвижном состоянии очень похожи на сучок дерева; бабочка каллима со сложенными крыльями удивительно напоминает сухой лист дерева), либо, наоборот, демонстративное поведение, отпугивающее хищников.
Физиологические адаптации - приобретение специфических особенностей обмена веществ в разных условиях среды. Они обеспечивают функциональные преимущества организма. Их условно разделяют на статические (постоянные физиологические параметры — температура, водно-солевой баланс, концентрация сахара и т. п.) и динамические (адаптации к колебаниям действия фактора — изменение температуры, влажности, освещенности, магнитного поля и т. п.).
Сравнительный обзор скелета позвоночных животных. Скелет головы. Осевой скелет. Скелет конечностей. Основные тенденции прогрессивной эволюции. Онтофилогенетически обусловленные пороки развития скелета у человека.
Скелет
У хордовых скелет внутренний. По строению и функциям подразделяется на осевой, скелет конечностей и головы.
Осевой скелет
В подтипе Бесчерепные имеется только осевой скелет в виде хорды. Она построена из сильно вакуолизированных клеток, плотно прилегающих друг к другу и покрытых снаружи общими эластической и волокнистой оболочками. Упругость хорде придают тургорное давление ее клеток и прочность оболочек. Хорда закладывается в онтогенезе всех хордовых и выполняет у более высокоорганизованных животных не столько опорную, сколько морфогенетическую функцию, являясь органом, осуществляющим эмбриональную индукцию.
На протяжении всей жизни у позвоночных хорда сохраняется только у круглоротых и некоторых низших рыб. У всех остальных животных она редуцируется. У человека в постэмбриональном периоде сохраняются рудименты хорды в виде nucleuspulposus межпозвоночных дисков. Сохранение избыточного количества хордального материала при нарушении его редукции чревато возможностью развития у человека опухолей — хордом, возникающих на его основе.
У всех позвоночных хорда постепенно вытесняется позвонками, развивающимися из склеротомов сомитов, и функционально заменяется позвоночным столбом. Это один из выраженных примеров гомотопной субституции органов. Формирование позвонков в филогенезе начинается с развития их дуг, охватывающих нервную трубку и становящихся местами прикрепления мышц. Начиная с хрящевых рыб обнаруживается охрящевение оболочки хорды и разрастание оснований позвонковых дуг, в результате чего формируются тела позвонков. Срастание верхних позвонковых дуг над нервной трубкой образует остистые отростки и позвоночный канал, в который заключена нервная трубка.
Замещение хорды позвоночным столбом — более мощным органом опоры, имеющим сегментарное строение,— позволяет увеличить общие размеры тела и активизирует двигательную функцию. Дальнейшие прогрессивные изменения позвоночного столба связаны с тканевой субституцией — заменой хрящевой ткани на костную, что обнаруживается у костных рыб, а также с дифференцировкой его на отделы.
У рыб только два отдела позвоночника: туловищный и хвостовой. Это связано с перемещением их в воде за счет изгибов тела.
Земноводные приобретают также шейный и крестцовый отделы, представленные каждый одним позвонком. Первый обеспечивает большую подвижность головы, а второй — опору задним конечностям.
У пресмыкающихся удлиняется шейный отдел позвоночника, первые два позвонка которого подвижно соединены с черепом и обеспечивают большую подвижность головы. Появляется поясничный отдел, еще слабо отграниченный от грудного, а крестец состоит уже из двух позвонков.
Млекопитающие характеризуются стабильным количеством позвонков в шейном отделе, равным 7. В связи с большим значением в движении задних конечностей крестец образован 5—10 позвонками. Поясничный и грудной отделы четко отграничены друг от друга.
У рыб все туловищные позвонки несут ребра, не срастающиеся друг с другом и с грудиной. Они придают телу устойчивую форму и обеспечивают опору мышцам, изгибающим тело в горизонтальной плоскости. Эта функция ребер сохраняется у всех позвоночных, совершающих змеевидные движения,— у хвостатых земноводных и пресмыкающихся, поэтому у них ребра также располагаются на всех позвонках, кроме хвостовых.
У пресмыкающихся часть ребер грудного отдела срастается с грудиной, формируя грудную клетку, а у млекопитающих в состав грудной клетки входит 12—13 пар ребер.
Онтогенез осевого скелета человека рекапитулирует основные филогенетические стадии его становления: в периоде нейруляции закладывается хорда, заменяющаяся впоследствии хрящевым, а затем и костным позвоночником. На шейных, грудных и поясничных позвонках развивается по паре ребер, после чего шейные и поясничные ребра редуцируются, а грудные срастаются спереди друг с другом и с грудиной, формируя грудную клетку.
Нарушение онтогенеза осевого скелета у человека может выразиться в таких атавистических пороках развития, как несрастание остистых отростков позвонков, в результате чего формируется spinabifida — дефект позвоночного канала. При этом часто через дефект выпячиваются мозговые оболочки и образуется спинномозговая грыжа.
В возрасте 1,5—3 мес. зародыш человека обладает хвостовым отделом позвоночника, состоящим из 8—11 позвонков. Нарушение их редукции в последующем объясняет возможность возникновения такой известной аномалии осевого скелета, как персистирование хвоста.
Нарушение редукции шейных и поясничных ребер лежит в основе их сохранения в постнатальном онтогенезе.
Скелет головы
Продолжением осевого скелета спереди является осевой, или мозговой, череп, служащий для защиты головного мозга и органов чувств. Рядом с ним развивается висцеральный, или лицевой череп, образующий опору передней части пищеварительной трубки. Обе части черепа развиваются по-разному и из разных зачатков. На ранних этапах эволюции и онтогенеза они не связаны между собой, но позже эта связь возникает.
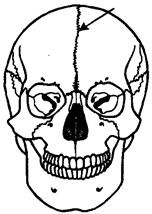
Рис. 14.8. Череп человека с методическим швом (указан стрелкой)
В задней части осевого черепа в процессе развития обнаруживаются следы сегментации, поэтому считают, что он представляет собой результат слияния друг с другом закладок передних позвонков. В состав мозгового черепа включаются также закладки хрящевых капсул мезенхимального происхождения, окружающие органы слуха, обоняния и зрения. Кроме того, часть мозгового черепа (лежащая кпереди от турецкого седла), не имеющая сегментации, развивается, по-видимому, как новообразование в связи с увеличением размеров переднего мозга.
Филогенетически мозговой череп прошел три стадии развития: перепончатую, хрящевую и костную.
У круглоротых он практически весь перепончатый и не имеет передней, несегментированной, части.
Череп хрящевых рыб почти полностью хрящевой, причем включает в себя как заднюю, первично сегментированную, часть, так и переднюю.
У костных рыб и остальных позвоночных осевой череп становится костным за счет процессов окостенения хряща в области его основания (основная, клиновидная, решетчатая кости) и за счет возникновения покровных костей в верхней его части (теменные, лобные, носовые кости). Кости осевого черепа в процессе прогрессивной эволюции претерпевают олигомеризацию. Появление большого количества зон окостенения и последующее слияние их вместе при формировании таких костей, как лобная, височная и др., свидетельствуют об этом. Широко известны у человека такие аномалии мозгового черепа, как наличие межтеменных, а также двух лобных костей с методическим швом между ними (рис. 14.8). Никакими патологическими явлениями они не сопровождаются и обнаруживаются поэтому обычно случайно после смерти.
Висцеральный череп впервые появляется также у низших позвоночных. Он формируется из мезенхимы эктодермального происхождения, которая группируется в виде сгущений, имеющих форму дужек, в промежутках между жаберными щелями глотки. Первые две дужки получают особенно сильное развитие и дают начало челюстной и подъязычной дугам взрослых животных. Следующие дуги в числе 4—5 пар выполняют опорную функцию для жабр и называются жаберными.
У хрящевых рыб впереди челюстной дуги располагаются обычно еще 1—2 пары предчелюстных дуг, имеющих рудиментарный характер. Это свидетельствует о том, что у предков позвоночных имелось большее количество висцеральных дуг, чем б или 7, а их дифференцировка происходила на фоне олигомеризации.
Челюстная дуга состоит из двух хрящей. Верхний называют нёбно-квадратным, он выполняет функцию первичной верхней челюсти. Нижний, или меккелев, хрящ — первичная нижняя челюсть. На вентральной стороне глотки меккелевы хрящи соединены друг с другом таким образом, что челюстная дуга кольцом охватывает ротовую полость. Вторая висцеральная дуга с каждой стороны состоит из гиомандибулярного хряща, сращенного с основанием мозгового черепа, и гиоида, соединенного с меккелевым хрящом. Таким образом, у хрящевых рыб обе первичные челюсти соединены с осевым черепом через вторую висцеральную дугу, в которой гиомандибулярный хрящ выполняет роль подвеска к мозговому черепу. Такой тип соединения челюстей и осевого черепа называют гиостильным.
У костных рыб начинается замещение первичных челюстей вторичными, состоящими из накладных костей — челюстной и предчелюстной сверху и зубной внизу. Нёбно-квадратный и меккелев хрящи при этом уменьшаются в размерах и смещаются кзади. Гиомандибулярный хрящ продолжает выполнять функции подвеска, поэтому череп остается гиостильным.
Земноводные в связи с переходом к наземному существованию претерпели значительные изменения висцерального черепа. Жаберные дуги частично редуцируются, а частично, меняя функции, входят в состав хрящевого аппарата гортани. Челюстная дуга своим верхним элементом — нёбно-квадратным хрящом — срастается полностью с основанием мозгового черепа, и череп становится, таким образом, аутостильным. Гиомандибулярный хрящ, сильно редуцированный и освободившийся от функции подвеска, располагаясь в области первой жаберной щели внутри слуховой капсулы, взял на себя функцию слуховой косточки — столбика,— передающей звуковые колебания от наружного к внутреннему уху.
Висцеральный череп пресмыкающихся также аутостилен. Для челюстного аппарата характерна более высокая степень окостенения, чем у земноводных. Часть хрящевого материала жаберных дуг входит в состав не только гортани, но и трахеи.
Нижняя челюсть млекопитающих сочленяется с височной костью сложным суставом, позволяющим не только захватывать пищу, но и совершать сложные жевательные движения.
Одна слуховая косточка — столбик,— характерная для земноводных и пресмыкающихся, уменьшаясь в размерах, превращается в стремечко, а рудименты нёбно-квадратного и меккелева хрящей, полностью выходящие из состава челюстного аппарата, преобразуются соответственно в наковаленку и молоточек. Таким образом, создается единая функциональная цепь из трех слуховых косточек в среднем ухе, характерная только для млекопитающих.
Рекапитуляция основных этапов филогенеза висцерального черепа происходит и в онтогенезе человека. Нарушение дифференцировки элементов челюстной жаберной дуги в слуховые косточки является механизмом формирования такого порока развития среднего уха, как расположение в барабанной полости только одной слуховой косточки — столбика, что соответствует строению звукопередающего аппарата земноводных и пресмыкающихся.
Скелет конечностей
У хордовых выделяются непарные и парные конечности. Непарные (спинные, хвостовой' и анальный плавники) являются основными органами передвижения у бесчерепных, рыб и в меньшей степени у хвостатых амфибий. У рыб возникают также парные конечности — грудные и брюшные плавники, на базе которых впоследствии развиваются парные конечности наземных четвероногих животных.
Подробнее рассмотрим происхождение и эволюцию парных конечностей.
У личинок рыб, а также у современных бесчерепных вдоль тела с обеих сторон тянутся боковые кожные складки, называемые метаплевральными (рис. 14.10). Они не имеют ни скелета, ни собственной мускулатуры, выполняя пассивную роль — стабилизацию положения тела и увеличение площади брюшной поверхности, облегчающие перемещение в водной среде. Вероятно, у предков рыб, переходящих к более активному образу жизни, в этих складках появились мышечные элементы и хрящевые лучи, связанные с сомитами по происхождению и поэтому расположенные метамерно. Такие складки, приобретя подвижность, могут выполнять роль рулей глубины, однако для изменения положения тела в пространстве большее значение имеют их передние и задние отделы, как наиболее удаленные от центра тяжести. Поэтому эволюция шла по пути интенсификации функций крайних отделов и ослабления функций центральных частей.
В результате из передних отделов складок развились грудные, а из задних — брюшные плавники. Не исключено, что формированию только двух пар конечностей на боковых сторонах тела предшествовал распад сплошных складок на ряд парных плавников, большее значение из которых также имели передние и задние. Об этом свидетельствует существование ископаемых остатков древнейших низкоорганизованных рыб с многочисленными плавниками. За счет слияния оснований хрящевых лучей возникли плечевой и тазовый пояса. Остальные их участки дифференцировались в скелет свободных конечностей.
У большинства рыб в скелете парных плавников выделяют проксимальный отдел, состоящий из небольшого числа хрящевых или костных пластинок, и дистальный, в состав которого входит большое количество расположенных радиально сегментированных лучей. С поясами конечностей плавники соединены малоподвижно. Они не могут служить опорой телу при передвижении по дну или суше. У кистеперых рыб скелет парных конечностей имеет иное строение. Общее количество их костных элементов уменьшено, и они имеют более крупные размеры. Проксимальный отдел состоит только из одного крупного костного элемента, соответствующего плечевой или бедренной костям передних или задних конечностей. Далее следуют две более мелкие косточки, гомологичные локтевой и лучевой или большой и малой берцовым костям. На них опираются 7—12 радиально расположенных лучей. В соединении с поясами конечностей у такого плавника участвуют только гомологи плечевой или бедренной костей, поэтому плавники кистеперых рыб оказываются активно подвижными и могут использоваться не только для изменения направления движения в воде, но и для перемещения по твердому субстрату.
Жизнь этих рыб в мелких пересыхающих водоемах в девонском периоде способствовала отбору форм с более развитыми и подвижными конечностями. Наличие у них добавочных органов дыхания стало второй предпосылкой выхода на сушу и возникновения других адаптации к наземному существованию, результатом чего явилось происхождение земноводных и всей группы Tetrapoda. Первые их представители — стегоцефалы — обладали семи- и пятипалыми конечностями, сохраняющими сходство с плавниками кистеперых рыб.
В скелете запястья сохранено правильное радиальное расположение костных элементов в 3—4 ряда, в пястье располагается 7—5 костей, а далее также радиально лежат фаланги 7—5 пальцев.
У современных земноводных количество пальцев в конечностях равно пяти или происходит их олигомеризация до четырех.
Дальнейшее прогрессивное преобразование конечностей выражается в увеличении степени подвижности соединений костей, в уменьшении количества костей в запястье сначала до трех рядов у амфибий и затем до двух — у пресмыкающихся и млекопитающих. Параллельно уменьшается также и количество фаланг пальцев. Характерно также удлинение проксимальных отделов конечности и укорочение дистальных.
Расположение конечностей в ходе эволюции также меняется. Если у рыб грудные плавники находятся на уровне первого позвонка и обращены в стороны, то у наземных позвоночных в результате усложнения ориентации в пространстве появляется шея и возникает подвижность головы, а у пресмыкающихся и особенно у млекопитающих в связи с приподнятием тела над землей передние конечности перемещаются кзади и ориентируются не горизонтально, а вертикально. То же касается и задних конечностей.
Многообразие условий обитания, предоставляемых наземным образом жизни, обеспечивает многообразие форм передвижения: прыжки, бег, ползание, полет, рытье, лазание по скалам и деревьям, а при возвращении в водную среду — и плавание. Поэтому у наземных позвоночных можно встретить как почти неограниченное многообразие конечностей, так и их полную вторичную редукцию, причем многие сходные адаптации конечностей в разнообразных средах многократно возникали конвергентно. Однако в процессе онтогенеза у большинства наземных позвоночных проявляются общие черты в развитии конечностей: закладка их зачатков в виде малодифференцированных складок, формирование в кисти и стопе вначале шести или семи зачатков пальцев, крайние из которых вскоре редуцируются и в дальнейшем развивается только пять.
Интересно, что в эмбриогенезе высших позвоночных рекапитулирует не только строение конечностей предков, но и процесс их гетеротопии. Так, у человека верхние конечности закладываются на уровне 3—4-го шейных позвонков, а нижние — на уровне поясничных позвонков. В это же время конечности получают иннервацию из соответствующих отделов спинного мозга. Гетеротопия конечностей сопровождается формированием шейного, поясничного и крестцового нервных сплетений, нервы которых связаны с одной стороны с теми сегментами спинного мозга, из которых они вырастали в момент формирования конечностей, а с другой — с конечностями, переместившимися на новое место.
В онтогенезе человека возможны многочисленные нарушения, ведущие к формированию врожденных пороков развития конечностей атавистического плана. Так, полидактилия, или увеличение количества пальцев, наследующаяся как аутосомно-доминантный признак, является результатом развития закладок дополнительных пальцев, характерных в норме для далеких предковых форм. Известен феномен полифалангии, характеризующийся увеличением числа фаланг обычно большого пальца кисти. В основе его возникновения — развитие трех фаланг в первом пальце, как это в норме наблюдается у пресмыкающихся и земноводных с недифференцированными пальцами конечностей. Двусторонняя полифалангия наследуется аутосомно-доминантно.
Серьезным пороком развития является нарушение гетеротопии пояса верхних конечностей из шейной области на уровень 1—2-го грудных позвонков. Эту аномалию называют болезнью Шпренгеля или врожденным высоким стоянием лопатки. Она выражается в том, что плечевой пояс с одной либо с двух сторон находится выше нормального положения на несколько сантиметров. В связи с тем что такое нарушение часто сопровождается аномалиями рёбер, грудного отдела позвоночника и деформацией лопаток, следует думать, что механизмы его возникновения — не только нарушение перемещения органов, но и обусловленное этим нарушение морфогенетических корреляций.
Сравнительно-анатомический обзор эволюции скелета хордовых свидетельствует о том, что скелет человека полностью гомологичен опорному аппарату предковых и родственных форм. Поэтому многие пороки его развития у человека можно объяснить родством млекопитающих с пресмыкающимися, земноводными и рыбами. Однако в процессе антропогенеза появились такие особенности скелета, которые характерны лишь для человека и связаны с его прямохождением и трудовой деятельностью. К ним относят: 1) изменения стопы переставшей выполнять хватательную функцию, выражающиеся в потере способности к противопоставлению большого пальца и появлению ее сводов, служащих для амортизации при ходьбе; 2) изменения позвоночного столба — его S-образный изгиб, обеспечивающий пластичность движений в вертикальном положении; 3) изменения черепа — резкое уменьшение его лицевой части и увеличение мозговой, смещение большого затылочного отверстия кпереди, увеличение сосцевидного отростка и сглаживание затылочного рельефа, к которому прикрепляются мышцы шеи и выйная связка; 4) специализация верхних (передних) конечностей как органа труда; 5) появление подбородочного выступа в связи с развитием членораздельной речи.
Несмотря на то что становление анатомо-морфологических особенностей скелета у человека, по-видимому, завершено, адаптации к прямохождению у него имеют, как и все адаптации вообще, относительный характер. Так, при большой физической нагрузке возможно смещение позвонков или межпозвонковых дисков. Человек, перейдя к прямохождению, утратил способность к быстрому бегу и передвигается значительно медленнее большинства четвероногих животных.
Естественно, что в ходе внутриутробного развития черты скелета, характеризующие человека как уникальный биологический вид, формируются на конечных его этапах либо даже, как, например, S-образная форма позвоночника, в раннем постнатальном периоде развития. Они являются фактически анаболиями, возникшими в ходе филогенеза приматов. Поэтому атавистические аномалии скелета, связанные с задержками развития признаков, характерных только для человека, встречаются наиболее часто. Они практически не снижают жизнеспособность, но дети, обладающие ими, нуждаются в ортопедической коррекции, гимнастике и массаже. К таким аномалиям относят легкие формы врожденного плоскостопия, косолапости, узкую грудную клетку, отсутствие подбородочного выступа и некоторые другие.
ПИЩЕВАРИТЕЛЬНАЯ
И ДЫХАТЕЛЬНАЯ СИСТЕМЫ
Уникальной особенностью организации хордовых является филогенетическая, эмбриональная, а также функциональная связь пищеварительной и дыхательной систем. Действительно, только у хордовых дыхательная система развивается на базе пищеварительной и на первых этапах эволюции функционирует совместно с ней. Так, у ланцетника, сохранившего в значительной степени черты организации предковых форм, специализированных органов дыхания нет, а дыхательную функцию выполняет глотка — передняя часть пищеварительной трубки, пронизанная сквозными отверстиями — жаберными щелями, главной функцией которых является фильтрация воды. Челюстного аппарата у ланцетника нет, и активно питаться он не может. Поэтому источником его питания является взвесь органических частиц в воде, которые задерживаются жаберными перегородками, прилипая к слизи, обильно выделяемой железистыми клетками глотки. Постоянный ток свежей воды через глотку способствует газообмену в кровеносных сосудах, расположенных в жаберных перегородках. На капилляры эти сосуды не распадаются, что свидетельствует о второстепенности дыхательной функции глотки.
У более высокоорганизованных хордовых начиная с рыб пищеварительная и дыхательная функции осуществляются специализированными системами, объединенными анатомически общей полостью рта и глотки, а также развитием из общего энтодермального зачатка.
Тесная связь обеих систем в филогенезе определяется в первую очередь их топографическими и динамическими координациями, а развитие в онтогенезе — морфогенетическими и эргонтическими корреляциями. Пищеварительная и дыхательная системы хордовых в эмбриогенезе закладываются вначале в виде прямой трубки, подразделяющейся на три участка. Переднюю ее часть, начинающуюся ротовым отверстием и заканчивающуюся переходом в глотку, называют stomodeum. Слизистая оболочка, выстилающая этот участок, эктодермального происхождения и в развитии связана с кожным эпидермисом и его производными. Средняя часть кишки начинается глоткой и заканчивается в том месте, где ее энтодермальная слизистая оболочка контактирует с эктодермальной слизистой оболочкой задней кишки, или proctodeum.
Ротовая полость
Рассмотрим вначале эволюцию ротовой полости и ее производных. У бесчерепных ротовая полость окружена предротовой воронкой со щупальцами и частично выстлана мерцательным эпителием, который вместе с таким же эпителием глотки создает постоянный ток воды в кишечную трубку, несущую пищевые частицы и кислород. Ротовое отверстие позвоночных окружено кожными складками — губами, которые становятся подвижными только у сумчатых и плацентарных млекопитающих в связи со вскармливанием детенышей молоком.
Крыша ротовой полости образована у рыб и земноводных основанием мозгового черепа, которое является первичным твердым нёбом. Хоаны земноводных открываются в их ротовую полость сразу позади альвеолярной дуги верхней челюсти. У пресмыкающихся объем ротовой полости увеличивается, и на верхнечелюстных и нёбных костях появляются горизонтальные складки, частично разделяющие ее на верхний, дыхательный, отдел и вторичную ротовую полость. Хоаны при этом несколько смещаются кзади. У млекопитающих наблюдается срастание этих складок по средней линии таким образом, что возникает сплошное вторичное твердое нёбо, полностью отделяющее друг от друга ротовую полость и полость носа. Хоаны при этом открываются в носоглотку. Этим достигается независимость функций органов ротовой полости от процесса дыхания.
До 7-й недели эмбрионального развития человека дифференцировка области stomodeum отсутствует. К концу 8-й недели происходит формирование вторичного твердого нёба за счет срастания горизонтальных костных складок. При нарушении адгезии клеток обеих складок возможно незаращение твердого нёба — порок развития, известный под названием волчья пасть. Эта аномалия имеет атавистическую природу. Доказаны генетические механизмы ее возникновения. Она сопровождает ряд хромосомных синдромов, а также наследуется изолированно, причем с разной частотой у населения разных популяций. Так, в Японии частота «волчьей пасти» равна 2,1, а в Нигерии — 0,4 случая на 1000 рождений.
Зубы позвоночных связаны по происхождению с плакоидной чешуей хрящевых рыб.У них наблюдается непрерывный переход от типичных чешуи к зубам в ротовой полости.Зубы первоначально расположены во много рядов и покрывают всю слизистую оболочку ротовой полости, располагаясь у многих рыб даже на языке. У ряда земноводных зубы также расположены не только на альвеолярной дуге, но и на других костях, например на сошнике. У пресмыкающихся обнаруживается только один ряд зубов, причем, так же как у земноводных и рыб, дифференцировка их отсутствует. Такую зубную систему, в которой все зубы одинаковы, называют гомодонтной. У животных перечисленных классов зубы, как и плакоидные чешуи, могут многократно выпадать, сменяясь новыми их поколениями. Многократную смену зубов называют полифиодонтизмом.
Зубы млекопитающих дифференцированы на резцы, клыки и коренные. Они выполняют различные функции. Такую зубную систему называют гетеродонтной. Если резцы, и в особенности клыки, млекопитающих еще весьма сходны с коническими зубами предков, то наибольшим эволюционным преобразованиям подверглись коренные зубы. Впервые они вознили у зверозубых ящеров начала мезозойской эры за счет расширения оснований зубов и появления дополнительных бугорков, а затем и увеличения жевательных поверхностей при их сглаживании. Общее количество зубов у млекопитающих уменьшается и достигает у высших приматов 32. Зубы располагаются только на альвеолярных дугах челюстей, в ячейках. Основание зуба сужается, образуя корень.
Зубы человека по сравнению с другими приматами уменьшены в размерах, особенно клыки. Моляры имеют четырехбугорчатое строение. Зубная дуга округлой формы. В связи с дифференцировкой зубов увеличилась продолжительность их функционирования, в результате чего в онтогенезе сменяются только два их поколения: молочные и коренные. Это явление называют дифиодонтизмом.
У человека возможны атавистические аномалии зубной системы, связанные с нарушениями как дифференцировки зубов, так и с их количеством. Редкой аномалией является гомодонтная зубная система, в которой все зубы имеют коническую форму. Более частой патологией является трехбугорчатое строение коренных зубов. Нередко встречается прорезывание сверхкомплектных зубов в ряду или за его пределами, иногда даже на твердом нёбе. Это свидетельствует о том, что у человека возможно образование большего количества зубных зачатков, чем 32, как это в норме встречается у низших млекопитающих и представителей более отдаленных классов позвоночных. Свидетельством тенденции к дальнейшему уменьшению количества зубов у человека является то, что нередко последние коренные зубы, так называемые «зубы мудрости», вообще не прорезываются, а если и прорезываются, то это происходит поздно — до 25 лет. Кроме того, эти зубы имеют явно рудиментарный характер, уменьшены в размерах и часто слабо дифференцированы.
На дне ротовой полости позвоночных располагается непарный выступ — язык, который у рыб представляет собой складку слизистой оболочки, лишенную мышц. Его движения осуществляются вместе с челюстями и подъязычной костью. У наземных позвоночных в языке располагаются мышцы, и они определяют его собственную подвижность. У пресмыкающихся и млекопитающих язык закладывается из трех зачатков: одного непарного и двух парных, лежащих кпереди от первого. Парные зачатки позже срастаются и дают начало телу языка. У большинства рептилий это срастание неполное, и язык раздвоен на конце. Весьма редким пороком развития языка у человека является раздвоенность его конца как результат несращения парных зачатков в эмбриогенезе.
Слюнные железы у рыб, заглатывающих пищу вместе с водой, отсутствуют и появляются только при переходе на сушу. Земноводные, обитающие на земле, имеют несколько слюнных желез, выделяющих только слизь. Слюна пресмыкающихся содержит уже и пищеварительные ферменты, а у некоторых змей—и токсины.У млекопитающих слюнные железы многочисленны: это и мелкие — зубные, щечные, нёбные, язычные, гомологичные железам земноводных и пресмыкающихся, и крупные — подъязычные, подчел







