

История создания датчика движения: Первый прибор для обнаружения движения был изобретен немецким физиком Генрихом Герцем...

Общие условия выбора системы дренажа: Система дренажа выбирается в зависимости от характера защищаемого...
История создания датчика движения: Первый прибор для обнаружения движения был изобретен немецким физиком Генрихом Герцем...
Общие условия выбора системы дренажа: Система дренажа выбирается в зависимости от характера защищаемого...
Топ:
Выпускная квалификационная работа: Основная часть ВКР, как правило, состоит из двух-трех глав, каждая из которых, в свою очередь...
Проблема типологии научных революций: Глобальные научные революции и типы научной рациональности...
Интересное:
Мероприятия для защиты от морозного пучения грунтов: Инженерная защита от морозного (криогенного) пучения грунтов необходима для легких малоэтажных зданий и других сооружений...
Влияние предпринимательской среды на эффективное функционирование предприятия: Предпринимательская среда – это совокупность внешних и внутренних факторов, оказывающих влияние на функционирование фирмы...
Средства для ингаляционного наркоза: Наркоз наступает в результате вдыхания (ингаляции) средств, которое осуществляют или с помощью маски...
Дисциплины:
|
из
5.00
|
Заказать работу |
|
|
|
|
ОШИБКИ МИТОЗА И ИХ ПОСЛЕДСТВИЯ
Ошибки митоза представляют собой нарушения, связанные с распределением генетического материала в ходе клеточного деления. Известно, что распределение наследственного материала имеет место в анафазе и происходит путем продольного расщепления центромеры, разделения хроматид и миграции их к полюсам клетки, за чем следует разделение цитоплазмы и образование двух дочерних клеток, каждая из которых содержит одинаковый набор хромосом. В силу различных причин (генетические и метаболические дефекты, действие факторов среды) эти процессы могут нарушаться. К таким нарушениям относятся следующие:
- нарушения в сборке веретена деления и взаимодействии кинетохоров с его нитями;
- асинхронная деполимеризация микротрубочек, вследствие чего хромосомы мигрируют к полюсам неодновременно;
- нарушения в структуре центромеры, препятствующие ее нормальному расщеплению и разделению сестринских хроматид;
- изменения вязкости цитоплазмы, что препятствует нормальному процессу перемещения хромосом к полюсам клетки и др.
Основные ошибки, ведущие к нарушениям распределения генетического материала:
- поперечное расщепление центромеры с образованием изохромосом;
- нерасхождение хроматид с образованием анеуплоидных клеток (трисомных и моносомных);
- анафазное отставание с образованием анеуплоидных клеток (моносомных).

Различные ошибки митоза и типы образованных клеток из исходной клетки с участием двух пара хромосом:
Нормальный набор хромосом – дисомия — 2n;
Аномальный набор хромосом:
- трисомия – 2n + 1;
- моносомия— 2n - 1;
наборы со структурными нарушениями хромосом:
- с iso p - 2n, i(?p);
- с iso q - 2n, i(?q).
|
|
Поперечное расщепление центромеры
В анафазе двухроматидные хромосомы разделяются на две хроматиды путем расщепления центромеры. В норме это расщепление происходит продольно, в результате чего образуются две идентичные по размерам, морфологии и генетическому материалу хромосомы, каждая из которых имеет два плеча: проксимальное (р) и дистальное (q). Иногда расщепление может происходить поперечно, что приводит к образованию двух разных хромосом, отличающихся по размерам, морфологии и генетическому материалу. Они называются изохромосомами и бывают двух типов:
- iso p (ip) - хромосома из двух плеч р, но без плеча q;
- iso q (iq)- хромосома из двух плеч q, но без плеча р.
В результате поперечного расщепления дочерние клетки будут иметь удвоенный генетический материал одного плеча и в них будет отсутствовать генетический материал другого плеча. Например:
46,X,i(Xp) - кариотип ♀ с iso p хромосомы X
46,X,i(Xq) - кариотип ♀, с iso q хромосомы X
46,X,i(Yp) - кариотип ♂, с iso p хромосомы Y
46,X,i(Yq) - cariotip ♂, с iso q хромосомы Y
46,XX (XY), i(21p) - кариотип ♀ или ♂, с iso p хромосомы 21
46,XX (XY), i(21q) - кариотип ♀ или ♂, с iso q хромосомы 21.
Хроматидное нерасхождение
Сестринские хроматиды, образованные в результате предшествующей митозу репликации, соединены в области центромеры до анафазы, когда они разделяются и мигрируют к полюсам. Этот процесс является ключевым моментом в распределении генетического материала в ходе деления. В случае нерасщепления/нерасхождения обе сестринские хроматиды одной хромосомы мигрируют к одному полюсу, что приводит к неравному распределению генетического материала. В результате одна клетка наследует одну дополнительную хромосому (2n+1 → 47 хромосом - трисомия ), в то время как в другой клетке одна их хромосом этой пары отсутствует (2n-1 →45 хромосом - мономосия). Например:
- 45,Х / 47,ХХХ - кариотип ♀, с моносомией X в одной клетке и трисомией X в другой клетке;
- 45,Х / 47,XYY- кариотип ♀/♂, с моносомией X в одной клетке и гоносомной трисомией
|
|
(дисомия Y) в другой клетке;
- 45,ХХ,-13 / 47,ХХ,+13 - кариотип ♀, с моносомией 13 в одной клетке и трисомией 13 в
другой клетке;
- 45,XY, -18 / 47,XY,+18 - кариотип с ♂, с моносомией 18 в одной клетке и трисомией 18 в другой клетке;
- 45,XY,-21 /47,XY,+21 - кариотип ♂,с моносомией 21 в одной клетке и трисомией 21 в другой клетке;
- 45,ХХ,-8 / 47,ХХ,+8 - кариотип ♀, с моносомией 8 в одной клетке и трисомией 8 в другой клетке.
Анафазное отставание
Разделившиеся после расщепления центромеры сестринские хроматиды (однохроматидные хромосомы) одновременно мигрируют к противоположным полюсам клетки. Таким образом, к концу анафазы на каждом из полюсов располагается по 46 хромосом. После реорганизации ядерной оболочки в телофазе и цитокинеза образуются две дочерние клетки с набором хромосом, идентичным материнской клетке. Иногда синхронность миграции хромосом к полюсам может нарушаться вследствие дефектов деполимеризации микротрубочек или изменения вязкости цитоплазмы. В результате, не все хромосомы достигнут одновременно полюсов и, соответственно, не попадут в дочерние клетки. "Опоздавшие" хромосомы образуют в цитоплазме микроядра, которые затем деградируют. Образованные в результате деления дочерние клетки имеют разные наборы хромосом: одна из клеток - нормальный диплоидный набор, в то время как другая клетка является моносомной. Например:
45,Х / 46,ХХ - кариотип ♀, с моносомией X в одной клетке и нормальным набором хромосом - в другой;
45,Х / 46,XY - кариотип ♀/♂, с моносомией X в одной клетке и нормальным набором хромосом - в другой;
45,ХХ,-13 / 46,ХХ - кариотип ♀, с моносомией 13 в одной клетке и нормальным набором хромосом - в другой;
45,XY, -18 / 46,XY - кариотип ♀,с моносомией 18 в одной клетке и нормальным набором хромосом - в другой.
Последствия ошибок митоза
Нарушения в распределении генетического материала приводят к появлению в организме клеток с различным набором хромосом, которые делятся и образуют два или более клеточных клона, отличающиеся по числу хромосом. Такие организмы называются мозаиками. В результате поперечного расщепления центромеры образуются следующие мозаики:
- 46,isop/46,isoq - если ошибка происходит при делении зиготы;
- 46 /46,isop /46,isoq - если ошибка происходит при любом последующем делении.
|
|

Хроматидное нерасхождение приводит к появлению мозаик 45/46/47 sau 45/47:
- 45/47 - если ошибка происходит при делении зиготы;
- 45/46/47 - если ошибка происходит при любом последующем делении.
Анафазное отставание является причиной образование мозаик 45/46.
Эволюция аномальных клонов зависит от жизнеспособности клеток с нарушениями:
- тяжелые аномалии, как правило, несовместимы с жизнью и вызывают гибель клеток (аномальные клоны элиминируются);
- при незначительных нарушениях аномальные клетки делятся и образуют аномальные клоны.
Трисомии характеризуются меньшими последствиями, чем моносомии, а нарушения половых хромосом не столь тяжелы, как нарушения аутосом. Кроме того, чем меньше хромосомы по размерам и количеству генов, тем к меньшим последствиям приводдят нарушения в них.
| Хромосомные аномалии | Ошибки митоза – причины хросомомных аномалий | Жизнеспособные хромосомные аномалии | Летальные хромосомные аномалии |
| Трисомии (47,+?) | Хроматидное нерахождение | 47,ХХХ 47,XXY 47,XYY 47,XX(XY),+21 47,XX(XY),+13 47,XX(XY)+18 47,XX(XY),+8 | Все остальные аутосомные трисомии |
| Моносомии (45,-?) | Хроматидное нерахождение Анафазное отставание | 45,X | Все аутосомные моносомии |
| i(?p) | Поперечное расщепление центромеры | 46,X,i(xp) 46,X,i(Yp) | Все остальные аномалии 46,XX(XY),i(p) |
| i(?q) | 46,X,i(Xq) 46,Xi(Yq) 46,XX(XY),i(Dq) 46,XX(XY),i(Gq) | Все остальные аномалии 46,XX(XY),i(q) |

Фенотипические последствия ошибок митоза зависят от времени их возникновения в онтогенезе. Если они происходят в эмбриогенезе - нарушается нормальное формирование тканей и органов с развитием врожденных аномалий (патологический фенотип при рождении); в случае,если нарушения возникают в постнатальный период, они приводят к изменению структуры или функций отдельно взятого органа или ткани и неоплазиям (опухоли).
ГАМЕТОГЕНЕЗ
Гаметогенез представляет собой сложный комплекс генетических, биохимических и морфологических процессов, лежащих в основе образования и созревания гамет в гонадах (семенниках и яичниках). Овогенез — это процесс формирования гаплоидных яйцеклеток из недифференцированных диплоидных овогониев. Он происходит в яичниках, причем начальные этапы имеют место еще до рождения девочки, в пренатальном периоде, а другие - только с наступлением половой зрелости и продолжаются до менопаузы. В процессе овогенеза есть две фазы ожидания: диктиотена профазы I и метафаза II мейоза. Сперматогенез — это процесс формирования гаплоидных сперматозоидов из недифференцированных диплоидных сперматогониев. Он происходит в семенниках, начинается с наступлением половой зрелости, включает несколько этапов и характеризуется постоянным обновлением половых клеток.
|
|
Гаметогенез состоит их следующих этапов:
- период размножения, когда клетки делятся митотически и образуют первичные половые клетки - гаметогонии. (2n=2с);
- период роста, в ходе которого происходит репликация ДНК, синтез необходимых для мейоза веществ, накопление факторов, обеспечивающих образование и развитие зиготы; в результате образуются гаметоциты I-го порядка (2n=46);
- период созревания, когда путем мейоза - решающего для гаметогенеза процесса - образуются гаплоидные гаметы:
- в овогенезе из одного овогония образуется только одна функциональная яйцеклетка и два/три полярных тельца;
- в сперматогенезе из одного сперматогония образуются четыре функциональных сперматозоида;
- период формирования/дифференциации, характерный только для сперматогенеза, когда в результате морфологических изменений сперматиды трансформируются в сперматозоиды, подвижные и способные к оплодотворению.

ОПЛОДОТВОРЕНИЕ
Процесс слияния гаплоидных гамет разного родительского происхождения называется оплодотворением. Образованная в результате этого клетка имеет диплоидный набор хромосом и называется зиготой. Половые клетки (гаметы) отличаются большим генетическим разнообразием, как следствие внутри- и межхромосомной рекомбинации в ходе гаметогенеза. Сочетание мужской и женской гамет при оплодотворении носит случайный характер (геномная рекомбинация), обеспечивая, тем самым, образование новых генетических вариантов в зиготе.
Кроме того, слияние гамет восстанавливает нормальный диплоидный набор хромосом, характерный для данного вида. Следующие за этим митотические деления обеспечивают рост и развитие многоклеточного организма. В соматических клетках организма хромосомы представлены парами, а хромосомы одной пары являются гомологичными - идентичными по морфологии и набору генов, но разными по происхождению.
Таким образом, организм взрослого человека содержит:
(i) соматические клетки, из которых состоят ткани и органы:
- имеют диплоидный набор хромосом (2n=46);
- возникают в результате митотических делений зиготы;
- 51% генетического материала имеет материнское происхождение (включая митохондриальную ДНК) и 49% - отцовское;
|
|
- делятся митотически, обеспечивая рост, регенерацию и обновление тканей;
(ii) половые клетки, которые обеспечивают передачу наследственной информации от родителей детям и образование зиготы; имеют гаплоидный набор хромосом (n=23);
образуются из диплоидных гаметогониев - специализированных клеток, обеспечивающих гаметогенез;
- созревают в гонадах в результате мейоза
- определяют постоянство числа хромосом данного вида.
ДИНАМИКА ХРОМОСОМ В МЕЙОЗЕ
Мейоз ("meiosis" - уменьшение) является сложным процессом из двух последовательных делений, завершающихся уменьшением числа хромосом в два раза. Из каждой клетки с 46 хромосомами (диплоидный набор) образуются 4 гаметы с 23 хромосомами (гаплоидный набор). Гаметы отличаются чрезвычайным генетическим разнообразием вследствие внутрихромосомной и межхромосомной рекомбинации.
Мейозу предшествует интерфаза, во время которой имеет место репликация ДНК. Между двумя мейотическими делениями есть очень непродолжительная фаза - интеркинез - в ходе которой ДНК не реплицируется.
Первое мейотическое деление - редукционное, обеспечивает уменьшение числа хромосом в два раза. В результате этого деления из гаметоцитов 1-го порядка с 46 двухроматидными хромосомами образуются гаметоциты II-го порядка с 23 двухроматидными хромосомами.
Сокращение числа хромосом в два раза обеспечивается следующими механизмами:
- конъюгация гомологичных хромосом с образованием 23 бивалентов (тетрад) в профазе I;
- расположение бивалентов в экваториальной плоскости в метафазе I;
- расщепление гомологичных хромосом и миграция их к противоположным полюсам, по одной к каждому из них;

Рис. Особенности расщепления хромосом в мейозе овогенеза и сперматогенеза
- цитокинез обеспечивает разделение цитоплазматической массы с образованием двух клеток с гаплоидным набором двухроматидных хромосом (1n=2с) - гаметоцитов II-го порядка.
Одновременно с процессами, обеспечивающими распределение хромосом в ходе редукционного деления, происходит рекомбинация генетического материала:
- внутрихромосомная рекомбинация — в результате кроссинговера, или обмена участками (генами) между материнской и отцовской гомологичными хромосомами, в профазе I, благодаря конъюгации и образованию бивалентов;
- межхромосомная рекомбинация - результат независимого сочетания негомологичных отцовских и материнских хромосом в ходе их распределения к полюсам клетки в анафазе I,благодаря случайному расположению бивалентов в экваториальной плоскости.

Биологическая роль мейоза
- обеспечивает воспроизводство организмов и передачу родительских признаков и, тем самым, - связь между родителями и детьми;
- увеличивает генетическое разнообразие в популяции человека путем внутрихромосомной (в профазе I) и межхромосомной (в анафазе I) рекомбинации;
- является материальной основой основных законов наследственности (закон независимого наследования, закона сцепленного наследования).
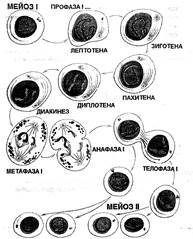
Динамика хромосом в мейозе
Ошибки оплодотворения
Двойное оплодотворение - возможно в случае, когда в процессе овуляции образуются две яйцеклетки, которые, будучи оплодотворенными двумя сперматозоидами, дают начало двум зиготам; зиготы, в дальнейшем могут развиваться независимо или совместно, образуя, в последнем случае, одну эмбриональную структуру, называемую химерой; если образующие химеру зиготы одного генетического пола, то отклонений в репродуктивном развитии обычно не наблюдается, в то время как, если зиготы имеют разный генетический пол, то кариотип организма будет XX/XY с вытекающими из этого нарушениями в дифференцировке пола - истинный гермафродитизм.
Диспермия - наблюдается тогда, когда одна яйцеклетка оплодотворяется двумя сперматозоидами, приводя к образованию триплоидных зигот (69,ХХХ или 69,XXY или 69,XYY).
Дигиния или диандрия - процесс слияния нормальной гаплоидной гаметы с диплоидной (яйцеклеткой или сперматозоидом); образованные триплоидные зиготы развиваются с серьезными нарушениями, как правило, несовместимыми с жизнью.

ЛЕКЦИЯ 3
ГЕНЫ ЧЕЛОВЕКА
Свойства генов человека
1. Репликация. Гены, являясь участками ДНК, реплицируются и передаются от клетки к клетке (в митозе) и от поколения к поколению (в мейозе). Это обеспечивает преемственность наследственной информации в ряду поколений и генеалогическую передачу признаков - наследственность;
2. Специфичность - один ген кодирует один полипептид, определяет один признак;
3. Эффект дозы. Ген обладает дозированным действием, то есть количество конечного продукта гена (РНК или полипептид) строго определено;
4. Стабильность. Гены стабильны, благодаря преемственности наследственного материала, что определяет сходство между родителями и детьми. Исключением являются нестабильные гены, которые реорганизуются в ходе дифференцировки (например, гены тяжелых и легких цепей иммуноглобулинов);
5. Различная экспрессивность. Проявление некоторых генов зависит от негенетических факторов и отличается при разных условиях или у различных лиц; при этом факторы среды могут: модулировать проявление генов (усиливать или ослаблять); изменять экспрессию генов (измененное проявление, отсутствие проявления);
6. Плейотропное действие. Плейотропия, или множественное действие гена, позволяет одному гену контролировать несколько признаков. Различают первичную и вторичную плейотропию. В случае первичной плейотропии ген непосредственно контролирует проявление нескольких признаков, а при вторичной плейотропии результат экспрессии плейотропного гена влияет на другие признаки (в разных клетках, тканях, органах);
7. Множественный аллелизм. Гены могут существовать в нескольких молекулярных формах. Путем генных мутаций возникают новые варианты гена - аллели, которые контролируют различные состояния или формы одного и того же признака. Эти аллели образуют серии и называются множественными, а контролируемые ими признаки называются полиморфными. Около 25% генов человека характеризуются множественным аллелизмом.
ФУНКЦИИ ГЕНОВ ЧЕЛОВЕКА
Функциями генов человека является хранение и передача наследственной информации о полипептидах и различных признаках (биохимических, морфологических, физиологических, психических и поведенческих).
Гены передают наследственную информацию благодаря репликации ДНК и деления клетки тем самым осуществляют связь между поколениями и обеспечивают генеалогическое наследование признаков в пределах семьи, вида.
Реализация наследственной информации обеспечивается транскрипцией в РНК и трансляцией в ходе синтеза белка - материального субстрата различных признаков на уровне клетки, ткани и организма.
ЛОКАЛИЗАЦИЯ ГЕНОВ
Согласно хромосомной теории наследственности Т.Моргана (1911): (1) гены расположеныв хромосомах в линейном порядке, занимая определенное место - локус; (2) гены одной хромосомы образуют одну группу сцепления и наследуются вместе; (3) количество групп сцепления равно гаплоидному числу хромосом; (4) между гомологичными хромосомами может происходить обмен участками - кроссинговер (5) частота кроссинговера прямо пропорциональна расстоянию между генами и обратно пропорциональна силе сцепления; (6) расстояние между генами измеряется в сантиморганидах - 1сМ = 1% кроссинговера.

Гены, расположенные в идентичных локусах гомологичных хромосом выполняют одинаковые функции и называются аллельными, в то время как гены, расположенные в разных локусах гомологичных или негомологичных хромосом, называются неаллелъными
.

Распределение генов вдоль хромосом неодинаково: хромосомы отличаются по количеству и плотности расположения генов. Некоторые гены представлены в одном экземпляре, а другие - многими копиями и образуют семейства повторяющихся и неповторяющихся генов.
Гены одной хромосомы, расположенные близко друг от друга, образуют гаплотип и имеют часто общие регуляторные элементы. В широком смысле под гаплотипом понимают совокупность генов одной молекулы ДНК.
Гены, локализованные в аутосомах, определяют аутосомные признаки и наследуются независимо от пола, а гены, расположенные в гоносомах, определяют сцепленные с полом признаки, и их наследование зависит от пола:
- Х-сцепленные гены и признаки передаются от матери к дочерям и сыновьям, а от, отца - только дочерям;
- Y-сцепленные гены и признаки (голандрические) передаются исключительно от отца к сыну.
ГЕНЕТИЧЕСКИЕ КАРТЫ
В состав генома человека входят две различающиеся по организации и особенностям наследования системы: ядерный геном и митохондриальный геном. В ядре соматических клеток человека содержится около 30 000 пар генов, которые локализованны в 46 молекулах ДНК.
Каждая хромосома содержит в среднем 2000 генов. Гены расположены в линейном порядке и разделены некодирующими последовательностями (сателлитная ДНК, спейсеры). Гены одной хромосомы наследуются совместно. Это явление называется сцеплением генов. Каждая хромосома, таким образом, представляет одну группу сцепления. Митохондриальный геном представлен кольцевыми молекулами ДНК и содержит 37 генов, расположенными очень компактно и наследуемыми по материнской линии.

Явление сцепление характерно только для генов одной и той же хромосомы, в то время как расположенные в разных хромосомах гены наследуются независимо, по законам Менделя. Сцепление бывает полным и неполным. Причина неполного сцепления - кроссинговер, происходящий в мейозе. В ходе кроссинговера происходит взаимный обмен аллельными генами.
Частота кроссинговера неодинакова для различных локусов и может варьировать от 0% до 50%, коррелируя с расстоянием между генами. Чем далее друг от друга расположены гены, тем больше частота кроссинговера между ними и, наоборот. Данное положение позволяет определить расстояние между генами по частоте кроссинговера и лежит в основе составления генетических карт. Эти карты представляют собой графическое изображение хромосом и расположенных на них генов с указанием расстояния между ними.

В настоящее время, благодаря методам дифференциальной окраски и соматической гибридизации, разработаны физические карты хромосом, в которых показано точное расположение генов с указанием расстояния между ними в парах нуклеотидов.
Установление сцепления между генами и определение групп сцепления представляет значительный интерес в медицинской генетике. Примерами сцепленных генов у человека являются: ген фактора Rh и ген эллиптоцитоза; ген АВО и ген пигментной ксеродермы (ХР); гены Duffy и врожденной катаракты; ген MNSs и dentinogenesis imperfecta-1 (DI-1); ген Xg и гены гемофилии А (НЕМА), гемофилии В (НЕМВ), дальтонизмаI (Dalt).

МЕТОДЫ АНАЛИЗА ГЕНОВ
Методы рекомбинантной ДНК создали предпосылку для разработки новых методов молекулярной диагностики с большей разрешающей способностью и, следовательно, более точных и информативных. Об абсолютном преимуществе молекулярного подхода говорит тот факт, что в отличие от других методов диагностики, ограниченных выявлением исключительно фенотипических аспектов, анализ ДНК, направленный непосредственно на изучение генотипа, является единственным методом, изучающим первичные нарушения (мутации), то есть именно первопричину возникновения болезней.
Технология рекомбинантной ДНК позволяет определить нормальные гены и/или мутантные их варианты, установить носителей мутантного гена, диагностировать наследственную патологию до рождения ребенка или в предсимптоматической стадии, а в ближайшем будущем проводить и генную терапию.
Молекулярное изучение генов может осуществляться многими путями в зависимости от поставленной цели:
1. секвестрование ДНК для определения первичной структуры гена;
2. метод Саузерн-блот для определения позиции гена в геноме;
3. метод Нозерн-блот для определения экспрессии генов (анализ мРНК);
4. метод Вестерн-блот для определения белкового продукта гена;
5. метод ПЦР для специфичной амплификации фрагментов ДНК.
В лабораториях молекулярной биологии используются различные варианты перечисленных методов.
Все перечисленные методы основаны на различных принципах манипулирования нуклеиновыми кислотами:
- специфическая рестрикция ДНК с целью получения интересующего фрагмента;
- идентификация фрагментов ДНК или РНК е помощью специальных зондов, комплементарных искомому участку;
- идентификация нормальных и патологических генов путем ПЦР - специфическая реакция отбора праймеров, комплементарных гену/последовательности;
- визуализация необходимого фрагмента по результатам электрофореза и специфического маркирования ДНК/РНК с использованием компьютерных программ чтения и анализа результатов;
- комплексная интерпретация результатов в зависимости от использованного метода.
| Для разделения полученных фрагментов ДНК применяется электрофорез макромолекул в агарозном или полиакриламидном геле. Будучи отрицательно заряжены, фрагменты молекул нуклеиновых кислот в электрическом поле перемещаются с разной скоростью в зависимости от молекулярной массы. Более короткие фрагменты перемещаются быстрее, в то время, как более длинные - медленнее. Для определения размеров фрагментов в геле, одновременно с интересующими фрагментами помещаются на соседние дорожки и фрагменты - маркеры длины. Молекулы нуклеиновых кислот могут быть обнаружены в геле при окраске флуоресцентным красителем или по радиоактивной метке. В случае радиоактивного мечения, фрагменты определяются с помощью авторадиографии, которая заключается в накладывании на гель светочувствительной пленки. |

Секвенирование ДНК
Секвенирование заключается в определении последовательности нуклеотидов (азотистых оснований) в исследуемом фрагменте ДНК. Анализ последовательностей нуклеотидов ДНК может быть осуществлен двумя путями: 1) химическим (метод Максима - Гильберта), при котором используются химические реакции последовательного расщепления фрагмента ДНК на отдельные нуклеотиды, однако этот метод очень трудоемкий, сложный и в последнее время не используется; 2) ферментативным (метод Сэнгера), при котором фрагменты ДНК синтезируются in vitro таким образом, что реакция завершается в положении, соответствующем определенному основанию. Для определения последовательности нуклеотидов любым из этих методов, ДНК подвергается серии из 4-х отдельных реакций, каждая из которых является специфической для одного из 4-х видов нуклеотидов. При одновременном электрофорезе продукты реакции на 4-х соседних дорожках будут мигрировать по ним с разной скоростью. Размер синтезированных фрагментов можно определить, а, следовательно, может быть определен и порядок нуклеотидов в
ДНК.
Метод Сэнгера (дидеокси) использует ферментативный синтез одной цепи, комплементарной клонированной матрице. В ходе данного процесса синтез ДНК останавливается добавлением одного из дидезоксинуклеозидтрифосфата, аналога нуклеотидов. Дидезоксинуклеозидтрифосфат содержит в позиции 3' не группу ОН, а Н-группу, которая препятствует полимеризации нуклеотидов. Используя четыре разных аналога дидезокси в процессе синтеза новой цепи ДНК, можно, таким образом, определить каждый нуклеотид матричной цепи. Электрофорез полученных фрагментов позволяет определить порядок нуклеотидов ДНК.
В последние годы применяются методы автоматического секвенирования.
С целью диагностики носителей мутантного гена результат секвенирования сравнивают с первичной структурой гена. К сожалению, не для всех генов человека к настоящему моменту известны последовательности нуклеотидов, поэтому применяются и непрямые методы диагностики: сцепление с близлежащими ДНК-маркерами (определение гйпервариабельных мини- и микросателлитных повторов), определение характерных для данного гена полиморфных сайтов рестрикции, гибридизация со специфическими зондами и др.

Метод Саузерн-блот
Метод основан на специфическом анализе определенных фрагментов геномной ДНК/гена, полученных путем рестрикции ДНК одной или несколькими рестриктазами. Ферменты рестрикции действуют не случайно, а разрезают ДНК в строго определенных местах, поэтому в результате действия одной рестриктазой получают двухцепочечные фрагменты ДНК разной длины (количество фрагментов и их длина специфичны для данной рестриктазы). Учитывая полиморфизм ДНК/генов, обусловленный точечными мутациями, длины фрагментов специфичны и неодинаковы у разных индивидов (полиморфизм длины рестрикционных фрагментов - ПДРФ).
Для анализа ПДРФ ряда генов необходимо знать специфическую локализацию сайтов рестрикции. Эта информация может быть полезна для сравнения нормальных и мутантных генов для раннего выявления носителей патологических генов. На данном принципе основан метод Саузерн-блот, который позволяет идентифицировать интересующий фрагмент в смеси из тысячи разных фрагментов, полученных в результате рестрикции геномной ДНК. Идентификация необходимого фрагмента осуществляется на основе гибридизации ДНК-мишени с комплементарным меченым зондом.
Метод Саузерн-блота состоит из последовательности следующих этапов:
(1) выделение высокомолекулярной геномной ДНК из клетки;
(2) ферментативное расщепление ДНК разными рестриктазами с образованием фрагментов разной длины;
(3) разделение рестрикционных фрагментов в агарозном геле;
(4) денатурация фрагментов ДНК щелочным раствором;
(5) нейтрализация геля буферным раствором;
(6) капиллярный перенос фрагментов на нейлоновый или нитроцеллюлезный фильтр;
(7) гибридизация с одноцепочечными радиоактивными зондами;
(8) авторадиография для выявления гибридов: исследуемая ДНК / зонд.
Перенос фрагментов на нитроцеллюлезный фильтр - блоттинг - предложил Эдвард Саузерн.

В результате анализа полученного материала можно определить наличие или отсутствие некоторых сайтов рестрикции, характерных для изучаемого гена, которые ассоциируются с определенными мутациями. Различные варианты расположения сайтов рестрикции в ДНК у двух разных людей называют Полиморфизмом Длины Рестрикционных Фрагментов (ПДРФ). Этот полиморфизм используется как генетический маркер в изучений генотипа.
Практическое применение метода Саузерн-блот:
- выявление точечных мутаций, затрагивающих сайты рестрикции (появляются новые или исчезают старые сайты рестрикции) по изменению количества и длины фрагментов рестрикции;
- выявление мутаций типа делении, дупликации, инсерции более длинных фрагментов (50-100 п.н.) по изменению длины фрагментов рестрикции.
Это позволяет проводить пренатальную и пресимптоматическую диагностику патологических мутаций и выявление гетерозиготных носителей мутантных генов.
Метод Саузерн-блот имеет, однако, следующие недостатки: (1) не позволяет выявить точечные мутации, не затрагивающие сайтов рестрикции (в пределах фрагмента рестрикции); (2) является достаточно трудоемким, сложным и дорогим методом.
Метод Нозерн-блот
Метод состоит в переносе разделенных молекул РНК на нейлоновые или нитроцеллюлезные фильтры с последующей гибридизацией с мечеными зондами. Метод схож с методом Саузерн-блот, за исключением того, что выделенные и очищенные мРНК не подвергают рестрикции, а электрофорез не происходит в условиях денатурации.
Метод Нозерн-блот позволяет идентифицировать транскрипты анализируемых генов, количества мРНК, их размеры.
Метод Бестерн-блот
Этот метод заключается в определении специфического белка из смеси клеточных белков. Для этого белки разделяют электрофорезом в условиях денатурации, в присутствии додецилсульфатa Na. Белки, разделённые по молекулярной массе, переносят на плотный фильтр и обрабатывают специфическими мечеными антителами. Этот метод позволяет определить наличие или отсутствие белка, его размер и скорость экспрессии гена по количеству синтезированного белка.
Техника ПЦР в анализе генов
Метод ПЦР используется для выборочного размножения определенной последовательности гена. В результате получают гомогенные популяции фрагментов, которые используются в исследованиях по молекулярной генетике или диагностике. Для осуществления амплификации определенной последовательности необходимо знать первичную структуру нормального или мутантного гена и синтезировать специфические праймеры, комплементарные концам интересующих фрагментов. Праймеры представляют собой искусственно синтезированные одноцепочечные олигонуклеотиды (20-30 нуклеотидов). ПЦР основана на гибридизации исследуемой последовательности ДНК - праймер и полуконсервативной репликации ДНК.
Преимущества метода ПЦР следующие: требуются минимальные количества ДНК; высокая скорость амплификации (за несколько часов получают более миллиона копий одинаковых фрагментов); благодаря специфичности праймера амплифицируетея только нужный фрагмент, который затем используется как зонд в других методах.

НАСЛЕДСТВЕННЫЕ ПРИЗНАКИ
ГРУППЫ КРОВИ
Группа крови - это 100% наследственный биохимический признак, который определен присутствием на поверхности эритроцитов некоторых антигенных веществ (в основном белков или мукополисахаридов).
Система групп крови представляет собой совокупность антигенных веществ, контролирующих их генов и взаимодействий между генами, а также взаимоотношений генотип-фенотип. Группы крови изучают давно из-за необходимости учета их особенностей при переливании крови:
- 1900 - Landsteiner вперые описал систему АВО;
- 1940 - Landsteiner совместно с Weiner описал систему Rh, имеющую особое значение в несовместимости плод-мать;
- на сегодняшний день описаны более 20 систем среди которых: ABO, Rh, MN, Ss, Le, Xg.
ГРУППЫ КРОВИ СИСТЕМЫ АВО
Система АВО представлена следующими фенотипами - группами крови I (0), П (Al, A2), Ш (В), IV (А1В, А2В) и фенотипом Bombay (0). Генотипы обусловлены аллельными генами Al, A2, В, О, расположенными в хромосоме 9 и существующими в эпистатическом взаимодействии с генами Н и h, локализованными в хромосоме 19.
Отношения между аллелями могут быть следующими: кодоминирования: А1 и В; А2 и В;
- доминантности/рецессивности: А1 > О, А2 > О; В > О; А1 > А2; Н > h;
- эпистаза: Н и О, Al, A2, В.
[(Al>A2)=B]>o;H>h
Разнообразные отношения между аллелями можно объяснить следующим образом:
-
|
|
|

Своеобразие русской архитектуры: Основной материал – дерево – быстрота постройки, но недолговечность и необходимость деления...

Эмиссия газов от очистных сооружений канализации: В последние годы внимание мирового сообщества сосредоточено на экологических проблемах...

Автоматическое растормаживание колес: Тормозные устройства колес предназначены для уменьшения длины пробега и улучшения маневрирования ВС при...
Общие условия выбора системы дренажа: Система дренажа выбирается в зависимости от характера защищаемого...
© cyberpedia.su 2017-2024 - Не является автором материалов. Исключительное право сохранено за автором текста.
Если вы не хотите, чтобы данный материал был у нас на сайте, перейдите по ссылке: Нарушение авторских прав. Мы поможем в написании вашей работы!