

История создания датчика движения: Первый прибор для обнаружения движения был изобретен немецким физиком Генрихом Герцем...

Индивидуальные и групповые автопоилки: для животных. Схемы и конструкции...
История создания датчика движения: Первый прибор для обнаружения движения был изобретен немецким физиком Генрихом Герцем...
Индивидуальные и групповые автопоилки: для животных. Схемы и конструкции...
Топ:
Отражение на счетах бухгалтерского учета процесса приобретения: Процесс заготовления представляет систему экономических событий, включающих приобретение организацией у поставщиков сырья...
Теоретическая значимость работы: Описание теоретической значимости (ценности) результатов исследования должно присутствовать во введении...
Организация стока поверхностных вод: Наибольшее количество влаги на земном шаре испаряется с поверхности морей и океанов...
Интересное:
Национальное богатство страны и его составляющие: для оценки элементов национального богатства используются...
Инженерная защита территорий, зданий и сооружений от опасных геологических процессов: Изучение оползневых явлений, оценка устойчивости склонов и проектирование противооползневых сооружений — актуальнейшие задачи, стоящие перед отечественными...
Подходы к решению темы фильма: Существует три основных типа исторического фильма, имеющих между собой много общего...
Дисциплины:
|
из
5.00
|
Заказать работу |
|
|
|
|
Получить явное подтверждение тому, что квантовая когерентность действительно может играть в биологических системах ключевую роль, конечно же, отрадно, однако суть этой самой роли применительно к процессам, имеющим непосредственное отношение к функционированию мозга, пока совершенно не ясна. Наше понимание работы мозга, все еще очень смутное, сводится, по большей части, к классическому представлению (совпадающему, в основном, с тем, что предложили еще в 1943 году Маккаллох и Питтс), согласно которому нейроны и соединяющие их синапсы выполняют в мозге практически те же функции, что и транзисторы вместе с соединяющими их дорожками в печатных схемах современных компьютеров. Более детальная биологическая картина выглядит так: классические нервные сигналы распространяются из центрального тела нейрона (сомы) вдоль очень длинного волокна, называемого аксоном, причем от аксона в различных местах ответвляются отдельные отростки (см. рис. 7.1). Каждый отросток непременно заканчивается синапсом — соединением, посредством которого сигнал через синаптическую щель передается к следующему нейрону (как правило). Именно на этой стадии в процесс вступают химические вещества, называемые нейромедиаторами, — перемещаясь от одной клетки (нейрона) к другой, они переносят сообщение о возбуждении предыдущего нейрона. Такое синаптическое соединение приходится либо на древовидный отросток (дендрит) следующего нейрона (в большинстве случаев), либо на его сому. Одни синапсы являются по своей природе возбуждающими, их нейромедиаторы усиливают возбуждение следующего нейрона; другие же, напротив, — тормозящие, и их нейромедиаторы (отличные от первых) возбуждение следующего нейрона ослабляют. Воздействие различных синапсов на нейрон суммируется (возбуждение учитываем со знаком «плюс», а торможение — со знаком «минус»), и по достижении определенного порогового значения нейрон возбуждается[51]. Правильнее, впрочем, будет сказать, что существует высокая вероятность такого возбуждения. Определенный случайный фактор присутствует во всех процессах такого рода.
|
|

Рис. 7.1. Нейрон и его соединение с другими нейронами посредством синапсов.
Таким образом — во всяком случае, пока, — не возникает сомнений в том, что изложенная картина может быть эффективно смоделирована численными методами, если допустить, что синаптические связи и их индивидуальная интенсивность со временем не изменяются. (Наличие случайных составляющих, разумеется, никаких проблем в смысле вычислимости не представляет, см. §1.9). В самом деле, несложно заметить, что вышеописанная нейронно-синапсовая схема (с постоянными синапсами и их интенсивностями) существенно эквивалентна схеме компьютера (см. НРК, с. 392—396). Однако благодаря феномену так называемой пластичности мозга, интенсивность по крайней мере некоторых синаптических связей может время от времени изменяться — порой быстрее, чем за секунду, — а кроме того, изменяться могут и сами связи. Что ставит нас перед немаловажным вопросом: что же этими синаптическими изменениями управляет?
В коннекционистских моделях (применяемых при разработке искусственных нейронных сетей) синаптические изменения описываются определенным вычислительным правилом. Это правило устанавливается таким образом, чтобы система могла в процессе работы повышать свою эффективность, сравнивая поступающую на ее вход извне информацию с некоторыми заранее заданными критериями. Простое правило такого типа предложил Дональд Хебб еще в 1949 году [193]. Современные коннекционистские модели87 используют различные модификации (порой весьма значительные) все той же процедуры Хебба. Любая модель такого рода непременно должна иметь в своей основе хоть какое-нибудь четкое вычислительное правило, поскольку выполняются эти модели на самых обычных компьютерах; см. §1.5. Однако, в силу изложенной в первой части аргументации, никакая вычислительная процедура не может адекватно объяснить все операционные проявления человеческого сознательного понимания. Следовательно, нужно искать какой-то другой управляющий «механизм» — по крайней мере, для объяснения синаптических изменений, возможно, имеющих некоторое отношение к настоящей сознательной деятельности мозга.
|
|
Были выдвинуты и другие идеи; например, Джеральд Эдельман в своей книге «Прозрачный воздух, сверкающий огонь» [112] (и в более ранней трилогии [109, 110, 111]) предположил, что в мозге действуют не правила типа правила Хебба, а, скорее, некий вариант «дарвиновского» эволюционного принципа, позволяющий мозгу непрерывно повышать свою эффективность, управляя синаптическими связями посредством своеобразного естественного отбора, — при этом Эдельман указывает на весьма многозначительные параллели между своей моделью и процессом развития иммунной системой способности «распознавать» вещества. Особое значение в этой модели придается сложной роли нейромедиаторов и других химических соединений, задействованных в коммуникации между нейронами. Однако на сегодняшний день соответствующие процессы по-прежнему рассматриваются как классические и вычислимые. Вместе со своими коллегами Эдельман даже построил ряд устройств с компьютерным управлением (получивших названия DARWIN I, II, III, IV и т.д.), предназначенных для моделирования (с увеличением степени сложности) как раз той самой процедуры, которая, по его предположению, лежит в основе умственной деятельности. Однако тот факт, что управляющие функции в устройствах Эдельмана возложены на самый обычный универсальный компьютер, вполне недвусмысленно показывает, что и эта схема является исключительно вычислительной — просто здесь используется некая «восходящая» система правил. При этом совершенно не важно, какими именно деталями данная схема отличается от других вычислительных процедур. Она все равно принадлежит к той категории, что мы обсуждали в первой части, — см. §1.5, а также §3.9 и краткое изложение аргументации главы 3 в воображаемом диалоге в §3.23. Одного лишь этого диалога достаточно для того, чтобы убедиться в полном неправдоподобии любого утверждения о том, что модель, основанная только на подобного рода принципах, может иметь какое-то отношение к действительному функционированию сознательного разума.
|
|
Для того, чтобы избавиться от этих «пут» вычислительности, необходимо найти какой-нибудь другой механизм управления синаптическими связями — причем каким бы этот механизм ни был, он, по всей видимости, должен задействовать некий физический процесс, важную роль в котором играет та или иная форма квантовой когерентности. Если этот процесс окажется в каком-либо существенном отношении похожим на действие иммунной системы, то, значит, и иммунная система работает на квантовых эффектах. Возможно, какие-то процессы в работе иммунного механизма распознавания и впрямь носят существенно квантовый характер — как, в частности, утверждает Майкл Конрад [57, 58, 59]. Меня бы это не удивило, однако в эдельмановской модели мозга возможному участию квантовых процессов в работе иммунной системы места не нашлось.
Впрочем, даже если когерентные квантовомеханические эффекты каким-то образом замешаны в управлении синаптическими связями, все же трудно предположить, что и распространение нервных импульсов может быть связано с чем-то существенно квантовомеханическим. Иначе говоря, совершенно неясно, какую пользу можно извлечь из рассмотрения квантовой суперпозиции, в которой нейрон одновременно и возбужден, и заторможен. Нервные сигналы представляются нам явлениями вполне макроскопическими — во всяком случае, достаточно макроскопическими для того, чтобы такая картина выглядела крайне неправдоподобно, даже несмотря на тот факт, что собственно передача весьма хорошо изолирована от окружения благодаря плотному слою миелина, покрывающему нервные окончания. Согласно критерию, предложенному в §6.12 (OR), следует ожидать, что при возбуждении нейрона объективная редукция состояния происходит очень быстро — не потому, что имеет место значительное перемещение масс (его там даже по минимально требуемым стандартам далеко недостаточно), а потому, что распространяющееся вдоль нерва электрическое поле (порождаемое нервным сигналом), скорее всего, не остается «незамеченным» окружающими нерв тканями мозга. Это поле возмущает случайным образом весьма значительный объем вещества окружения — вполне достаточный, как мне представляется, для того, чтобы удовлетворить критерию срабатывания процедуры OR (из §6.12) почти сразу же после возникновения сигнала. Таким образом, сохранение в течение длительного времени квантовых суперпозиций возбуждения и торможения нейрона вряд ли возможно.
|
|
Квантовые вычисления
Свойство возбужденного нейрона возмущать окружение всегда представлялось мне донельзя неудобным — оно никак не вписывалось в то предварительное предположение, которое я пытался обосновать в НРК и в рамках которого квантовая суперпозиция одновременного возбуждения и торможения семейств нейронов была, как мне казалось, действительно необходимой. Согласно нашему новому критерию редукции состояний (OR), для редукции требуется еще меньшее возмущение окружения, чем в прежнем описании, и в возможность сохранения таких суперпозиций в течение сколько-нибудь заметного времени поверить еще сложнее. А собственно идея тогда заключалась в следующем: если бы возможно было выполнять несколько отдельных «вычислений» в суперпозиции в нескольких одновременно возбуждающихся нейронных структурах, то резонно было бы предположить, что в мозге вместо «обычных» тьюринговых вычислений выполняется нечто вроде вычислений квантовых. Несмотря на кажущуюся невозможность выполнения квантовых вычислений на этом уровне функционирования мозга, будет полезно познакомиться с некоторыми их аспектами подробнее.
Квантовое вычисление — теоретическая концепция, основы которой разработали Дэвид Дойч [83] и Ричард Фейнман [120, 121] (см. также [25] и [6]) и которая в настоящее время активно исследуется многими учеными. Основная идея заключается в распространении классического понятия машины Тьюринга на соответствующее квантовое устройство. Как следствие, все выполняемые такой расширенной «машиной» операции должны подчиняться квантовым законам — т.е. законам, по которым живут системы квантового уровня (с возможностью суперпозиций). Так, эволюция устройства происходит преимущественно под действием процедуры U, причем существенным свойством этого самого действия является как раз сохранение наличествующих суперпозиций. Процедура R получает «право голоса», как правило, лишь в конце операции, когда система «измеряется» с целью узнать результат вычисления. Вообще говоря (хотя не все это осознают), в процессе вычисления процедуру R необходимо время от времени вызывать дополнительно для того, чтобы проверить, не завершилось ли оно.
|
|
Выяснилось, что, хотя квантовый компьютер и не имеет сверхспособностей, в принципе недоступных для традиционного вычисления по Тьюрингу, в некоторых классах задач квантовое вычисление превосходит тьюрингово вычисление в смысле теории сложности ([83]). То есть при решении таких задач квантовый компьютер оказывается в принципе намного быстрее, нежели компьютер обычный, — но и только. Ряд интересных (хотя и несколько искусственных) задач такого типа, при решении которых квантовый компьютер оказывается победителем, приводят, в частности, Дойч и Йожа [88]. Более того, как недавно показал Питер Шор, с помощью квантового вычисления можно решить (за полиномиальное время) актуальную задачу факторизации больших целых чисел.
«Стандартное» квантовое вычисление использует обычные правила квантовой теории, согласно которым в течение практически всей операции система эволюционирует под действием процедуры U, a R вмешивается в процесс на строго определенных этапах. В такой процедуре нет ничего «невычислимого» в смысле обычной «вычислимости», так как U — вычислимая операция, a R — чисто вероятностная процедура. Все, что в принципе можно получить с помощью квантового компьютера, можно в принципе получить и с помощью соответствующей машины Тьюринга, снабженной генератором случайных чисел. Таким образом, согласно представленным в первой части книги аргументам, даже квантовый компьютер не способен выполнять операции, требуемые для человеческого сознательного понимания. Остается надеяться лишь на то, что подлинная невычислимость скрывается где-то за тонкими особенностями процесса, в действительности происходящего в момент «кажущейся» редукции вектора состояния, потому что во временно заменяющей этот реальный процесс случайной процедуре R никакой невычислимости нет. Таким образом, полная теория гипотетической процедуры OR будет по необходимости носить существенно невычислимый характер.
Предложенная в НРК идея основывалась на предположении, что в мозге возможны достаточно длительные тьюринговы вычисления в суперпозиции, прерываемые время от времени неким невычислимым действием, которое можно объяснить лишь в терминах того нового физического процесса (например, OR), какой придет на смену редукции R. Теперь, когда на такие суперпозиции нейронных вычислений мы больше рассчитывать не можем по причине слишком сильного возмущения окружения проходящими по нейрону импульсами, становится непонятно, каким образом можно здесь хотя бы воспользоваться самой идеей стандартного квантового вычисления, не говоря уже о какой-либо модификации этой процедуры посредством замены R на некий гипотетический невычислимый процесс (например, OR). Однако, как мы очень скоро убедимся, существует еще одна, весьма многообещающая возможность. Для того чтобы понять, что она собой представляет, нам необходимо более подробно рассмотреть биологическое устройство клеток мозга.
Цитоскелет и микротрубочки
Если мы вдруг вообразим, что сложное поведение животных управляется только лишь нейронами, то скромная парамеция поставит нас перед фундаментальной проблемой. Эта инфузория перемещается по своему пруду с помощью многочисленных крохотных волосообразных конечностей — ресничек, — преследуя бактерий, которыми она питается и которых обнаруживает посредством различных внутренних механизмов, или отступая от возможной опасности, готовая мгновенно устремиться прочь. Она также может преодолевать препятствия, огибая их. Более того, парамеция, по всей видимости, способна обучаться на собственном опыте88 — хотя эта наиболее замечательная ее способность некоторыми учеными оспаривается89. Как же все это может проделывать существо, не имеющее ни единого нейрона и синапса? В самом деле, поскольку вся парамеция — это всего лишь одна, пусть и большая, клетка, и притом не нейрон, ей просто негде все перечисленные способности разместить (см. рис. 7.2).
Несомненно, поведение парамеции — да собственно и прочих одноклеточных организмов, например, амеб — регулируется какой-то сложной системой управления, просто эта система построена не из нервных клеток. Ответственная за поведение парамеции структура, очевидно, является частью ее так называемого цитоскелета. Как можно предположить из названия, цитоскелет служит для поддержания формы клетки, однако у него имеются и многочисленные иные функции. Упоминавшиеся выше реснички представляют собой окончания волокон цитоскелета, но помимо них цитоскелет, похоже, содержит еще и собственно систему управления движением клетки, а также систему «конвейеров», осуществляющих транспортировку молекул внутри клетки. Словом, в единичной клетке цитоскелет выступает в роли этакой комбинации скелета, мускулатуры, конечностей, системы кровообращения и нервной системы.
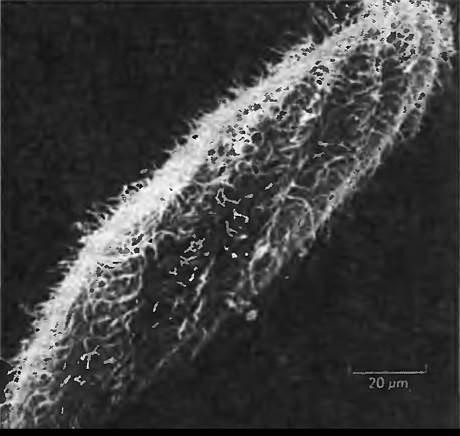
Рис. 7.2. Парамеция. Обратите внимание на волосообразные реснички, используемые для перемещения в воде. Они представляют собой наружные окончания цитоскелета парамеции.
Нас с вами в настоящий момент больше всего интересует, каким образом цитоскелет выполняет функции клеточной «нервной системы». Нейроны в нашем мозге сами являются отдельными клетками, причем у каждого нейрона есть свой собственный цитоскелет! Означает ли это, что в некотором смысле каждый отдельный нейрон располагает чем-то вроде «личной нервной системы»? Предположение весьма интригующее, и многие ученые склоняются к мнению, что нечто подобное действительно может иметь место. (См. первопроходческий труд Стюарта Хамероффа «Первичное вычисление: биомолекулярное сознание и нанотехнология» [183]; также рекомендую обратить внимание на статью [184] и многочисленные статьи в новом журнале «Нанобиология»[52].)
Прежде чем переходить к этим вопросам, необходимо рассмотреть вкратце общее устройство цитоскелета. Он состоит из протеиноподобных молекул, организованных в различного типа структуры: актин, микротрубочки и промежуточные волокна. Нас сейчас интересуют, главным образом, микротрубочки. Они представляют собой полые цилиндрические трубки с внешним диаметром около 25 нм и внутренним — около 14 нм (где «нм» обозначает «нанометр», т.е. 10—9 м), иногда организованные в более крупные трубкообразные волокна, состоящие из девяти дублетов, триплетов или частичных триплетов микротрубочек; в поперечном сечении такое волокно напоминает лопасти вентилятора, как показано на рис. 7.3, причем иногда по его центру также проходит пара микротрубочек. Как раз такое строение имеют реснички парамеции. Каждая микротрубочка представляет собой белковый полимер, состоящий из субъединиц, называемых тубулинами. Каждая субъединица тубулина, в свою очередь, представляет собой «димер», т.е. состоит из двух соединенных тонкой перемычкой частей, называемых α -тубулин и β -тубулин (приблизительно по 450 аминокислот в каждой). Эти, пары глобулярных белков, напоминающие по форме орех арахиса, уложены в слегка скошенную гексагональную решетку вдоль всей трубки, как показано на рис. 7.4. Обычно на каждую миктротрубочку приходится по 13 рядов димеров тубулина. Размеры димера составляют приблизительно 8 нм × 4 нм × 4 нм, а его атомное число — около 11 × 104 (т.е. в одном димере содержится такое количество нуклонов, что его масса в абсолютных единицах равна приблизительно 10—14).
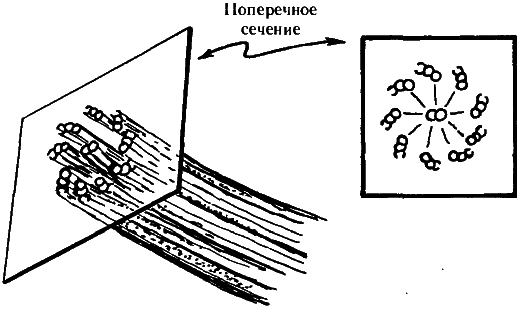
Рис. 7.3. Важной частью цитоскелета являются пучки крохотных трубочек (микротрубочек), организованных в структуры, напоминающие в поперечном сечении лопасти вентилятора. Такое строение имеют, например, реснички парамеции.
Димер тубулина может существовать в двух (по крайней мере) различных геометрических конфигурациях, называемых конформациями. В одной из таких конформаций молекулы тубулина располагаются под углом около 30° к оси микротрубочки. Есть основания полагать, что эти две конформаций соответствуют двум различным состояниям электрической поляризации димера, возникающим вследствие того, что электрон в центре перемычки α -тубулин/ β -тубулин занимает в различных конформациях различные положения.
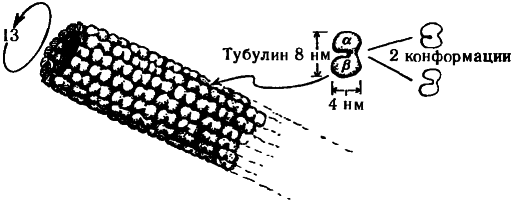
Рис. 7.4. Микротрубочка. Полая трубка, обычно состоящая из 13 рядов димеров тубулина. Каждая из молекул тубулина может существовать в двух (по крайней мере) конформациях.
«Центром управления» в цитоскелете является, по всей видимости, структура, называемая центром организации микротрубочек, или центросомой. Внутри центросомы имеется особая структура, называемая центриолью, которая состоит из двух цилиндрических волокон, по девять триплетов микротрубочек в каждом, образующих в пространстве структуру, похожую на «разделенную» букву «T» (см. рис. 7.5). (Цилиндрические волокна в общем аналогичны по структуре ресничкам, показанным на рис. 7.3.) Согласно Альбрехту-Бюлеру [7, 9], центриоль действует как глаз (!) клетки — идея чрезвычайно захватывающая, хотя и далеко еще не общепринятая. Какой бы ни была роль центросомы в нормальной, «повседневной», жизни клетки, она выполняет по крайней мере одну фундаментально важную задачу. На некоем критическом этапе она разделяется на две части, каждая из которых, по всей видимости, утягивает за собой пучок микротрубочек — хотя, пожалуй, точнее будет сказать, что каждая часть становится своего рода фокусом, вокруг которого и собираются микротрубочки. Эти микротрубочковые волокна каким-то образом связывают центросому с отдельными цепочками ДНК в ядре (в центральных точках, называемых центромерами), и цепочки ДНК расходятся — начиная тем самым удивительный процесс, известный специалистам под названием митоз, что означает всего-навсего деление клетки (см. рис. 7.6).

Рис. 7.5. Центриоль (по некоторым предположениям, глаз клетки) состоит из двух пучков микротрубочек (очень похожих на те, что изображены на рис. 7.3), образующих «разделенную» букву «T».

Рис. 7.6. При митозе (делении клетки) хромосомы разделяются, растаскиваемые пучками микротрубочек.
Может показаться странным, что внутри одной клетки действуют две столь разные «штаб-квартиры». Одна из них — ядро, где хранится основной генетический материал клетки, определяющий ее наследственность и уникальность, а также управляющий производством белкового материала, из которого, собственно, «строится» клетка. Другой управляющий центр — центросома с центриолью в качестве основного компонента, являющаяся, по всей видимости, главным узлом цитоскелета — структуры, которая, опять же по всей видимости, контролирует движение клетки и ее пространственную организацию. Предполагается, что присутствие этих двух различных «центров» в эукариотических клетках (клетках всех животных и почти всех растений на нашей планете, за исключением бактерий, сине-зеленых водорослей и вирусов) является результатом древней «инфекции», распространившейся по миру несколько миллиардов лет назад. Клетки, населявшие Землю прежде, были прокариотическими; они существуют и поныне в виде бактерий и сине-зеленых водорослей, и у них нет цитоскелета. Согласно одному из предположений [332], часть древнейших прокариот оказались каким-то образом связаны (возможно, «инфицированы») с неким видом спирохет (бактерий, перемещающихся с помощью нитеобразного хвоста, состоящего из цитоскелетных белков). Эти чуждые друг другу организмы постепенно «научились» жить вместе в симбиотической связи как единые эукариотические клетки. Так «спирохеты» превратились, в конечном счете, в цитоскелеты клеток — со всеми вытекающими последствиями для будущей эволюции, среди которых мы с вами!
Организация микротрубочек млекопитающих представляет интерес с математической точки зрения. На первый взгляд, число 13 не имеет какого-либо особого математического значения, однако это не совсем так. Оно принадлежит к знаменитой последовательности чисел Фибоначчи:
0, 1, 1, 2, 3, 5, 8, 13, 21, 34, 55, 89, 144, …
где каждое последующее число получается сложением двух предыдущих. Это может показаться случайным совпадением, однако хорошо известно, что числа Фибоначчи в биологических системах не редкость (и в гораздо более крупном масштабе). Например, в еловых шишках, цветках подсолнечника и пальмовых стволах наблюдаются спиральные или винтовые структуры с взаимопроникновением левых и правых закручиваний, причем количество рядов, закрученных в одном направлении, и количество рядов, закрученных в другом направлении, суть два соседних числа Фибоначчи (см. рис. 7.7). (Если внимательно рассмотреть такую структуру от одного конца до другого, можно обнаружить «место перехода», где числа рядов сменяются на следующую пару соседних чисел Фибоначчи.) Любопытно, что гексагональный узор микротрубочек демонстрирует очень похожую особенность — в общем случае даже еще более точно, — причем состоит этот узор (по крайней мере, обычно) из 5 правых и 8 левых винтовых структур, как показано на рис. 7.8. На рис. 7.9 я попытался изобразить, как такие структуры могли бы «выглядеть» изнутри микротрубочки. Число 13 выступает здесь как общее количество витков в спирали: 5 + 8. Любопытно также, что в двойных микротрубочках, встречающихся достаточно часто, внешний слой составной трубки обычно содержит 21 ряд димеров тубулина — следующее число Фибоначчи! (Не стоит, впрочем, чересчур увлекаться подобными построениями; например, в пучках микротрубочек в ресничках и центриолях бывает и по 9 рядов димеров — число, определенно не принадлежащее последовательности Фибоначчи.)

Рис. 7.7. Цветок подсолнечника. Как и во многих других растениях, отчетливо наблюдаются числа Фибоначчи. Во внешней области круга имеем 89 спиралей, закрученных по часовой стрелке, и 55 спиралей, закрученных против часовой стрелки. Ближе к центру появляются другие числа Фибоначчи.

Рис. 7.8. Представим, что микротрубочка разрезана вдоль и затем развернута в полосу. Можно видеть, что молекулы тубулина располагаются вдоль наклонных линий, причем каждый новый виток смещен относительно предыдущего на 5 или 8 молекул (в зависимости оттого, куда наклонена линия, вправо или влево).

Рис. 7.9. Заглянем внутрь микротрубочки! Можно наблюдать спиральную структуру молекул тубулина 5 + 8.
Откуда в структуре микротрубочек берутся числа Фибоначчи? Относительно еловых шишек, цветков подсолнечника и т.д. существует несколько вполне убедительных теорий — кстати, среди тех. кто серьезно занимался этим вопросом, был Алан Тьюринг (см. [198], с. 437). Однако к случаю микротрубочек эти теории, вполне возможно, неприменимы, и для такого уровня следует искать какие-то другие объяснения. Коруга [228] высказал предположение, что числа Фибоначчи в структуре микротрубочки повышают эффективность ее как «информационного процессора». В самом деле, согласно Хамероффу с коллегами (которые пытаются нам это втолковать вот уже более десяти лет90), микротрубочки могут действовать как клеточные автоматы, передавая и обрабатывая сложные сигналы в виде волн различных состояний электрической поляризации молекул тубулина. Вспомним, что димеры тубулина могут существовать в двух (по крайней мере) различных конформационных состояниях и способных переходить из одного состояния в другое; последнее, очевидно, обусловливается сменой электрической поляризации молекулы на альтернативную. На состояние каждого димера воздействуют состояния поляризации каждого из шести его соседей (вследствие ван-дер-ваальсовых взаимодействий между ними), т.е. существуют вполне конкретные правила, определяющие конформацию каждого димера через конформации его соседей. Благодаря этому обстоятельству, каждая микротрубочка способна осуществлять передачу и обработку любого рода сообщений. С распространением сигналов, похоже, как-то связана транспортировка различных молекул вдоль микротрубочек, а также всевозможные соединения между соседними микротрубочками в виде своеобразных белковых «мостиков» — так называемые MAP (от microtubule associated proteins[53]); см. рис. 7.10. Коруга доказывает, что в случае структуры с числами Фибоначчи, подобной той, что реально наблюдается в микротрубочках, информация обрабатывается особенно эффективно. Должно быть, для такой организации микротрубочек и в самом имеется серьезная причина, поскольку, несмотря на некоторый разброс в числах, наблюдаемый в эукариотических клетках вообще, микротрубочки почти всех млекопитающих составлены именно из 13 рядов димеров.

Рис. 7.10. Микротрубочки обычно соединяются друг с другом посредством «мостиков» из так называемых белков, ассоциированных с микротрубочками (MAP).
Для чего микротрубочки нейронам? Каждый отдельный нейрон имеет свой цитоскелет. Какова его роль? Я уверен, что будущим исследователям предстоит сделать в этой области еще немало открытий, однако кое-что мы знаем уже сейчас. В частности, микротрубочки нейронов могут быть очень и очень длинными (по сравнению с диаметром нейрона, который составляет лишь 25—30 нм) — до нескольких миллиметров или даже длиннее. Более того, в зависимости от обстоятельств они способны расти или сокращаться, а также транспортировать молекулы нейромедиаторов. Внутри аксонов и дендритов также имеются микротрубочки. Хотя, как правило, на всю длину аксона каждая отдельная микротрубочка не тянется, они образуют сообщающиеся сети, охватывающие всю клетку, соединяясь между собой посредством упоминавшихся выше MAP-мостиков. Микротрубочки, по-видимому, ответственны за поддержание интенсивности синапсов и, несомненно, за изменение этой интенсивности в случае необходимости. Более того, они, похоже управляют ростом новых нервных окончаний, направляя их к точкам соединений с другими нервными клетками.
Поскольку после окончательного формирования мозга деление нейронов прекращается, необходимости в этой функции центросомы здесь нет. В центросомах нейронов, расположенных вблизи ядра, часто вовсе нет центриолей. Микротрубочки тянутся от центросом к окрестности пресинаптических окончаний аксона, а также в другую сторону, к дендритам и, через сокращающиеся актиновые нити, к дендритным шипикам, часто образующим постсинаптические окончания синаптической щели. Эти шипики способны расти и вырождаться, что, по-видимому, является существенным элементом общей пластичности мозга, благодаря которой система взаимных соединений в мозге подвергается непрерывным тонким изменениям. Насколько мне известно, существуют убедительные экспериментальные свидетельства важной роли микротрубочек в управлении пластичностью мозга.
Упомянем еще об одном любопытном факте. В пресинаптических окончаниях аксонов содержатся некие ассоциированные с миктротрубочками вещества, «работа» которых связана с высвобождением нейромедиаторов, а молекулы весьма примечательны с геометрической точки зрения. Эти вещества — клатрины — строятся из белковых тримеров (так называемых клатриновых трискелионов), этаких полипептидных трехлучевых звезд. Объединяясь в молекулу клатрина, трискелионы образуют геометрически правильные структуры, идентичные по общему строению многоатомным молекулам углерода, называемым «фуллеренами» (а также «бакиболами», или «мячами Баки»[54]) из-за их внешнего сходства со знаменитыми геодезическими куполами, которые проектировал и возводил американский архитектор Бакминстер Фуллер91. Клатрины, впрочем, гораздо больше фуллереновых молекул, поскольку одному атому углерода в фуллерене соответствует в клатрине целый трискелион, состоящий из нескольких аминокислот. Те клатрины, что заняты в высвобождении нейромедиаторов в синапсах, имеют форму усеченного икосаэдра — всем нам знакомого многогранника, по образу и подобию которого делают современные футбольные мячи (см. рис. 7.11 и 7.12).
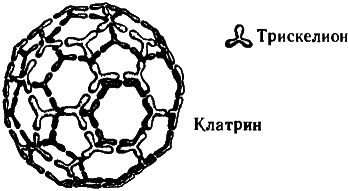
Рис. 7.11. Молекула клатрина (похожая общей структурой на фуллерен, но составленная не из атомов углерода, а из более сложных субструктур — белковых тримеров, называемых трискелионами). Изображенный на рисунке клатрин напоминает внешне обыкновенный футбольный мяч.
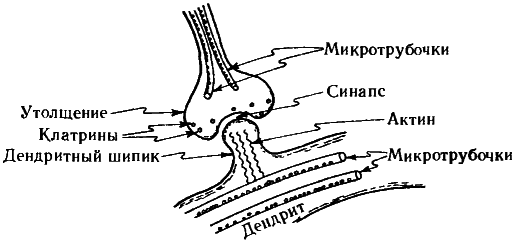
Рис. 7.12. Клатрины, подобные тому, что изображен на рис. 7.11, располагаются (вместе с окончаниями микротрубочек) в пресинаптическом утолщении аксона и, по всей видимости, участвуют в управлении интенсивностью синапса; также на интенсивность синапса влияют сокращающиеся актиновые нити в дендритных шипиках, управляемых микротрубочками.
В одном из предыдущих параграфов был поставлен важный вопрос: что управляет изменением интенсивности синапсов и определяет места размещения функционирующих синаптических связей? Учитывая имеющиеся свидетельства, можно уверенно предположить, что центральную роль в этих процессах играет цитоскелет. Как же это предположение может нам помочь в поиске невычислимой сущности разума? Пока что оно, похоже, говорит нам лишь о том, что потенциальная вычислительная мощность мозга оказывается гораздо большей, чем можно было бы ожидать, используй мозг в качестве простейших вычислительных блоков «цельные» нейроны.
В самом деле, если простейшими вычислительными блоками мы теперь будем считать димеры тубулина, то придется предположить, что потенциальная вычислительная мощность мозга просто неимоверно превосходит все то, что предполагали самые смелые теоретики от ИИ. Основываясь на «цельнонейронной» модели, Ханс Моравек в своей книге «Дети разума» [267] предположил, что человеческий мозг может в принципе достичь производительности порядка 1014 операций в секунду, но не более того; это при том, что в мозге имеется около 1011 функционирующих нейронов, каждый из которых способен посылать примерно по 103 сигналов в секунду (см. §1.2). Если же в качестве элементарного вычислительного блока взять димер тубулина, то следует учесть, что на каждый нейрон приходится около 107 димеров; соответственно, элементарные операции теперь выполняются где-то в 106 раз быстрее, в результате чего получаем 1027 операций в секунду. Возможно, производительность современных компьютеров и вправду уже начинает приближаться к первой цифре, 1014 операций в секунду (как весьма убежденно доказывают Моравек и его единомышленники), однако несмотря на все эти успехи, достичь в обозримом будущем производительности 1027 операций в секунду не представляется возможным.
Разумеется, можно смело утверждать, что мозг работает далеко не со стопроцентной «микротрубочковой» эффективностью, какую приведенные выше цифры предполагают. Тем не менее, ясно, что возможность «микротрубочкового вычисления» (см. [183]) позволяет совсем по-иному взглянуть на некоторые из аргументов в пользу неминуемого наступления эпохи искусственного интеллекта человеческого уровня. Можем ли мы теперь поверить хотя бы в то, что уже сегодня возможно92 численно воспроизвести умственную деятельность червя нематоды, только потому, что мы вроде бы «закартографировали» и численно смоделировали его нервную систему? Как было отмечено в §1.15, умственные способности обычного муравья намного превосходят все то, что на настоящий момент реализовано посредством стандартных ИИ-процедур. Впору поинтересоваться, сколько же муравей выигрывает в производительности благодаря гигантскому массиву своих «микротрубочковых информационных нанопроцессоров», если сравнивать с тем, чего он смог бы добиться, располагай он лишь «переключателями цельнонейронного типа». Что до парамеции, то тут, как вы понимаете, оснований для предъявления иска нет.
Однако аргументы, представленные в первой части, предполагают гораздо более сильное заявление. Я утверждаю, что способность человека к пониманию выходит за рамки какой угодно вычислительной схемы. Если мозгом человека управляют микротрубочки, то в микротрубочковых процессах должно быть что-то принципиально отличное от простого вычисления. Я утверждал, что такая невычислимая активность должна быть следствием достаточно макроскопической квантовой когерентности, объединенной неким тонким образом с макроскопическим поведением — с тем, чтобы обеспечить возможность протекания в системе тех новых физических процессов, что придут на смену бытующей в современной физике паллиативной R -процедуре. В качестве первого шага мы должны выяснить, какова же подлинная роль квантовой когерентности в цитоскелетной активности.
|
|
|

Организация стока поверхностных вод: Наибольшее количество влаги на земном шаре испаряется с поверхности морей и океанов (88‰)...

История развития хранилищ для нефти: Первые склады нефти появились в XVII веке. Они представляли собой землянные ямы-амбара глубиной 4…5 м...

Индивидуальные очистные сооружения: К классу индивидуальных очистных сооружений относят сооружения, пропускная способность которых...

Историки об Елизавете Петровне: Елизавета попала между двумя встречными культурными течениями, воспитывалась среди новых европейских веяний и преданий...
© cyberpedia.su 2017-2024 - Не является автором материалов. Исключительное право сохранено за автором текста.
Если вы не хотите, чтобы данный материал был у нас на сайте, перейдите по ссылке: Нарушение авторских прав. Мы поможем в написании вашей работы!