

Типы сооружений для обработки осадков: Септиками называются сооружения, в которых одновременно происходят осветление сточной жидкости...

Индивидуальные и групповые автопоилки: для животных. Схемы и конструкции...
Типы сооружений для обработки осадков: Септиками называются сооружения, в которых одновременно происходят осветление сточной жидкости...
Индивидуальные и групповые автопоилки: для животных. Схемы и конструкции...
Топ:
Устройство и оснащение процедурного кабинета: Решающая роль в обеспечении правильного лечения пациентов отводится процедурной медсестре...
Когда производится ограждение поезда, остановившегося на перегоне: Во всех случаях немедленно должно быть ограждено место препятствия для движения поездов на смежном пути двухпутного...
Особенности труда и отдыха в условиях низких температур: К работам при низких температурах на открытом воздухе и в не отапливаемых помещениях допускаются лица не моложе 18 лет, прошедшие...
Интересное:
Распространение рака на другие отдаленные от желудка органы: Характерных симптомов рака желудка не существует. Выраженные симптомы появляются, когда опухоль...
Инженерная защита территорий, зданий и сооружений от опасных геологических процессов: Изучение оползневых явлений, оценка устойчивости склонов и проектирование противооползневых сооружений — актуальнейшие задачи, стоящие перед отечественными...
Влияние предпринимательской среды на эффективное функционирование предприятия: Предпринимательская среда – это совокупность внешних и внутренних факторов, оказывающих влияние на функционирование фирмы...
Дисциплины:
|
из
5.00
|
Заказать работу |
|
|
|
|
Тема № 9
Формирование пространственной
Организации
Гилберт С. Гл. 17, 18. Т. 3.
Корочкин Л.И. С. 84 -124, 154 -172.
Градиентные модели позиционной информации. Краткий обзор развития дрозофилы. Сегрегационные и гомеозисные гены. Гомеобокс и гомеодомен. Роль гомеобокссодержащих генов в развитии млекопитающих. Гомеозисные гены и морфогенетические процессы. Гены, контролирующие гомеозисные гены. Эволюция на основе гомеозисных генов.
Формирование пространственной организации - это результат функциональной активности эмбриональных клеток, посредством которой осуществляется упорядоченное расположение дифференцированных тканей. Каким образом в зародыше не только возникают различные типы клеток, но и создаются функциональные ткани и органы? Одно дело - дифференцировка хондроцитов или остеоцитов, синтезирующих соответственно хрящевой и костный матриксы, другое дело - образовывать эти клетки в такой пространственно-временной ориентации, которая приводит к созданию функциональной кости. При этом должна выполняться еще одна задача, а именно создаваться определенная кость, например плечевая, тазовая или бедренная. Способность клеток узнавать свое положение в пределах ограниченной популяции клеток и дифференцироваться в соответствии с получаемой ими позиционной информацией стала объектом интенсивного изучения.
Формирование пространственной организации включает реализацию выбора одного из возможных путей развития. Каким образом происходит так, что клетка, потенциально способная стать хрящевой или мышечной, становится мышечной, если располагается в одном месте, и хрящевой, если оказывается в другом?
|
|
Градиентные модели позиционной информации
 Каким образом клетки получают информацию относительно своего положения в зародыше и как они используют эту информацию для дифференцировки в соответствующий клеточный тип? Чтобы объяснить это, были предложены различные гипотезы. В основе одной из них лежит представление о градиентах морфогенетических веществ (Бовери, 1901; Чайлд, 1941; Волперт, 1971). Предполагается, что растворимое вещество (морфоген) диффундирует от «источника» (где оно продуцируется) к «стоку» (где оно разрушается), образуя в области диффузии постепенный спад концентрации. Согласно теоретическим расчетам, такие градиенты могут проявлять свое действие на относительно небольших расстояниях (менее 100 клеточных диаметров).
Каким образом клетки получают информацию относительно своего положения в зародыше и как они используют эту информацию для дифференцировки в соответствующий клеточный тип? Чтобы объяснить это, были предложены различные гипотезы. В основе одной из них лежит представление о градиентах морфогенетических веществ (Бовери, 1901; Чайлд, 1941; Волперт, 1971). Предполагается, что растворимое вещество (морфоген) диффундирует от «источника» (где оно продуцируется) к «стоку» (где оно разрушается), образуя в области диффузии постепенный спад концентрации. Согласно теоретическим расчетам, такие градиенты могут проявлять свое действие на относительно небольших расстояниях (менее 100 клеточных диаметров).
Модель французского флага (Волперт, 1978)
Волперт иллюстрировал возможное действие отдельного градиента, представив плоскую ткань в виде французского флага (рис.). Предположим, что источник морфогена располагается на левой границе синей полосы, а на другом конце флага на правой границе красной полосы - находится место выведения морфогена. Таким образом, перед нами - градиент концентрации, причем на одном конце флага/ткани концентрация самая высокая, а на другом - самая низкая. Все клетки полипотентны, т. е. каждая может стать либо синей, либо белой, либо красной, и эта спецификация коррелирована с концентрацией морфогена. Клетки, получившие наибольшее количество морфогена, становятся синими. Однако существует порог концентрации морфогена, ниже которого клетки становятся белыми. При падении концентрации ниже следующего порога клетки становятся красными.
Генетика формирования пространственной организации
У дрозофилы
Из всех эукариот генетика именно дрозофилы была изучена наиболее полно. Анализ путей, посредством которых мутации оказывают влияние на пространственную организацию, стал ключом к пониманию механизмов морфогенеза.
Гены материнского эффекта
|
|
Эти гены контролируют формирование градиентов в ходе оогенеза. Анимально-вегетативный, дорсо-вентральный и терминальные структуры формируются несколькими независимыми генными системами.
Можно выделить две основные системы генов, особенно важных для формирования анимально-вегетативного градиента (передне-заднего).
Результаты эмбриологических экспериментов свидетельствуют о существовании в яйцах насекомых по крайней мере двух организующих центров: переднего и заднего. Именно эти области формируют два градиента: в передней и задней частях зародыша.
1) Среди генов первой группы главным является bicoid. У его мутантов вместо передних структур формируется дополнительный тельсон. Этот ген транскрибируется в клетках овариолы яичника и можно проследить транспорт этих клеток в анимальный полюс ооцита. Поэтому его мРНК обнаруживается исключительно в передней части ооцита. Но при ее трансляции белковый продукт гена формирует градиент с наибольшей концентрацией в передней части яйца и фоновой концентрацией – в задней.Для поддержания стабильности этого продукта в переднем полюсе зародыша необходима активность еще двух генов exuperantia и swallow. При их мутациях градиент белка bicoid становится размытым и зародыши не формируют передних структур.
2)  С другой стороны, фолликулярные клетки поставляют в ооцит мРНК, синтезированную геном nanos и концентрирующуюся на заднем конце зародыша. У мутантов по этому гену нарушается развитие заднего конца зародыша. Для его активации необходимы 5 генов с материнским эффектом. Белок образуется в области заднего полюса и затем транспортируется в область брюшных сегментов.
С другой стороны, фолликулярные клетки поставляют в ооцит мРНК, синтезированную геном nanos и концентрирующуюся на заднем конце зародыша. У мутантов по этому гену нарушается развитие заднего конца зародыша. Для его активации необходимы 5 генов с материнским эффектом. Белок образуется в области заднего полюса и затем транспортируется в область брюшных сегментов.
3) В формировании плана строения на самых ранних этапах созревания ооцита принимает участие еще один важный ген – hunchback. Он активируется геном bicoid, поэтому его продукт тоже накапливается в передней половине зародыша. Этот ген репрессирует гены, активные в брюшных сегментах, так что в зоне его распределения формируются головные и грудные структуры. Оказывается, белок гена bicoid обнаруживается в ядрах и содержит гомеодомен, с помощью которого он может связываться с ДНК и регулировать таким образом экспрессию генов. Было показано, что белок гена bicoid способен присоединяться к 5 сайтам, расположенным выше промотора гена hunchback и все эти участки имели общую последовательность. В норме белок nanos способен блокировать трансляцию мРНК hunchback, вероятно, в этом и заключается его основная функция, и в результате формируются задние структуры зародыша.
|
|
 Вторая система генов контролирует формирование дорсо-вентрального градиента. Последовательность активации этих генов представлена на рис. Как следует из рисунков, гены посылают сигнал (неизвестной природы) к рецептору, кодируемому геном torpedo, который функционирует в фолликулярных клетках. Белок torpedo частично гомологичен рецептору фактора роста (ЕGF-рецептору) позвоночных и имеет экстра-целлюлярную часть, способную связывать лиганд из ооцита. Затем гены pipe, nudel, windbeutel, функционирующие в фолликулярных клетках, посылают сигнал (также неидентифицированный) в вентральную область ооцита. После оплодотворения гены snake, easter, кодирующие протеазы, активируют ген spatzle, так что сигнал возвращается в ооцит. Продукт spatzle активирует toll на вентральной стороне яйца. Продукт этого последнего активирует продуцируемую геном реllе киназу. Она в свою очередь фосфорилирует продукт гена сасtus. В результате этого процесса высвобождается белковый продукт гена dorsal, поступающий в ядра клеток. Активация tоll-рецептора ведет к высвобождению белка dorsal из комплекса dorsal-cactus, так что создается градиент распределения белка dorsal, действующего как транскрипционный фактор.
Вторая система генов контролирует формирование дорсо-вентрального градиента. Последовательность активации этих генов представлена на рис. Как следует из рисунков, гены посылают сигнал (неизвестной природы) к рецептору, кодируемому геном torpedo, который функционирует в фолликулярных клетках. Белок torpedo частично гомологичен рецептору фактора роста (ЕGF-рецептору) позвоночных и имеет экстра-целлюлярную часть, способную связывать лиганд из ооцита. Затем гены pipe, nudel, windbeutel, функционирующие в фолликулярных клетках, посылают сигнал (также неидентифицированный) в вентральную область ооцита. После оплодотворения гены snake, easter, кодирующие протеазы, активируют ген spatzle, так что сигнал возвращается в ооцит. Продукт spatzle активирует toll на вентральной стороне яйца. Продукт этого последнего активирует продуцируемую геном реllе киназу. Она в свою очередь фосфорилирует продукт гена сасtus. В результате этого процесса высвобождается белковый продукт гена dorsal, поступающий в ядра клеток. Активация tоll-рецептора ведет к высвобождению белка dorsal из комплекса dorsal-cactus, так что создается градиент распределения белка dorsal, действующего как транскрипционный фактор.
Существует третья система генов, контролирующих формирование градиентов в оогенезе. Эта группа генов контролирует формирование терминальных структур, т.е. акрона (несегментированного головного конца) и тельсона (несегментированного хвостового конца). Ключевую роль в этом процессе играет ген torso. При отсутствии его продукта ни акрон, ни тельсон не развиваются, так что эмбрион оказывается полностью сегментированным. Активация белка torso осуществляется как на переднем, так и на заднем концах зародыша продуктом гена torsoless, функционирующим в фолликулярных клетках.
 Итак, гетерогенизация цитоплазмы созревающего ооцита и формирование полярных градиентов, химически преформирующих план строения будущего организма, реализуется на основе взаимодействия трех систем генов и при участии питающих клеток материнского организма, окружающих ооцит.
Итак, гетерогенизация цитоплазмы созревающего ооцита и формирование полярных градиентов, химически преформирующих план строения будущего организма, реализуется на основе взаимодействия трех систем генов и при участии питающих клеток материнского организма, окружающих ооцит.
|
|
gap-гены
Сегрегационные гены последовательно активируются в процессе индивидуального развития. В первую очередь активируются gap-гены (от англ. gap — брешь, пролом, щель). Их транскрипция стимулируется продуктами генов материнского эффекта, формирующими градиенты в ходе оогенеза. Они начинают функционировать на синцитиальной стадии развития, когда к 10—11-му циклу клеточного деления ядра мигрируют к периферии развивающегося эмбриона и "прочитывают" позиционную информацию, возникшую благодаря активности генов материнского организма, формируя на поверхности яйца синцитий. В результате этого процесса зародыш подразделяется на несколько пространственных доменов. Мутации gар-генов вызывают выпадения групп сегментов, характер их экспрессии изменяется во времени, но зоны экспрессии не перекрываются.
Pаir-rulе-гены
На фоне специфического распределения продуктов gар-генов под их влиянием активируются, раir-rulе-гены, которые "дробят" зародыш на повторяющиеся домены шириной по два парасегмента, в одном из которых этот ген активен, в другом — нет. Благодаря функционированию этих генов зародыш подразделяется на отдельные сегменты. Нарушение их функционирования ведет к выпадению отдельных (чередующихся) сегментов.
Гомеозисные гены
ОТКРЫТИЕ ГОМЕОЗИСНЫХ ГЕНОВ, ИХ РОЛЬ В РАЗВИТИИ
Название этой группы генов происходит от термина "гомеозис", который ввел в 1894 г. один из классиков генетики У.Бэтсон. Под гомеозисом он понимал превращение одной части тела в другую. Гомеозисные гены, следовательно, не представляют собой нечто самостоятельное, но являются частью специфической системы генов, контролирующих сегментацию тела насекомых, в частности дрозофилы и других организмов. Примером гомеозисных мутаций является превращение антенны в ногу или аристы в ногу.
ГЕННЫЕ КОМПЛЕКСЫ ANT-С И ВХ-С
Гомеозисные гены, которых у дрозофилы описано около полусотни, как уже отмечалось, контролируют качественные особенности сегментов и в свою очередь подразделяются на два комплекса: Antennapedia-Соmрlех (ANT-С) и Bithorax-Соmрlех (ВХ-С).
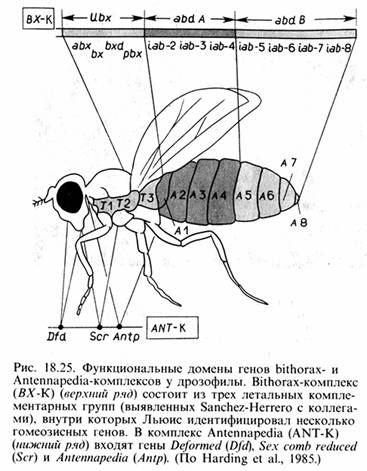 Гены, принадлежащие к АNТ-С, контролируют развитие головных сегментов, в том числе интеркалярного, максиллярного, мандибулярного, лабиального, а также грудных сегментов. При утрате функции гена Аntр область тела, включающая заднюю часть первого грудного сегмента Т1, весь второй грудной сегмент Т2 и переднюю часть ТЗ, приобретают свойства головных сегментов, что проявляется в образовании головных структур в грудной области. Можно сказать, что ген Аntр участвует в выборе программы развития сегмента в направлении головы или груди.
Гены, принадлежащие к АNТ-С, контролируют развитие головных сегментов, в том числе интеркалярного, максиллярного, мандибулярного, лабиального, а также грудных сегментов. При утрате функции гена Аntр область тела, включающая заднюю часть первого грудного сегмента Т1, весь второй грудной сегмент Т2 и переднюю часть ТЗ, приобретают свойства головных сегментов, что проявляется в образовании головных структур в грудной области. Можно сказать, что ген Аntр участвует в выборе программы развития сегмента в направлении головы или груди.
|
|
Гены комплекса BX-С ответственны за спецификацию грудных и брюшных сегментов. Мутации: bithoraxoid – 1-ый брюшной сегмент развивается как заднегрудь, в результате формируется 6 крыльев и 8 ног; bithorax-postbithorax – 4 крыла. Мутации обычно бывают вызваны мобильными элементами. Предполагается, что активация генов комплекса BX-С осуществляется последовательно – 2-ой торакальный сегмент является базисным, в каждом последующем активируется дополнительный ген, в последнем (8-ом) – все гены.
Характеристика экспрессии ГОМЕОЗИСНЫХ ГЕНОВ
Время первого появления транскриптов ВХ-С очень раннее — уже на 2—4-м часах после оплодотворения яйца, т.е. на стадии бластодермы, когда происходит процесс первичной детерминации тканевых закладок. Таким образом, гены морфогенеза функционируют как бы с опережением, задолго до осуществления контролируемых ими формообразовательных событий.
Можно предположить, что суть клеточной детерминации заключается в создании паттерна транскрипционно активных локусов, специфичного для данной ткани или клеточного типа. Однако определенного момента этот специфический рисунок функциональной организации генома не имеет выхода в специфику клеточного фенотипа, поскольку для активной продукции тканеспецифических веществ необходимо наличие сложного аппарата белкового синтеза. Следующий за клеточной детерминацией первый этап клеточной дифференцировки является, вероятно, процессом неспецифическим, направленным на формирование такого аппарата, и сопровождается высокой транскрипционной активностью house-keeping генов.
Как и предвидел Э.Льюис, гомеозисные гены функционируют повсеместно. При этом оказалось, что образуемый их транскриптами "тигровый" рисунок возникает очень рано, до четкого проявления признаков самой сегментации. Налицо, таким образом, еще один пример своеобразной химической преформации.
Экспрессия гомеозисных генов тканеспецифична. Так, ген Ubх транскрибируется в эктодерме и ее производных. В мышцах посегментная экспрессия менее выражена, в энтодермальных производных ее нет. При гибридизации на срезах ножные имагинальные диски в этом случае мечены, крыловые — нет. По всей вероятности, сегментоподобное распределение некоторых изоферментов в органах и тканях дрозофилы связано с функцией генов сегментации, в том числе и гомеозисных генов.
ГОМЕОБОКС И ГОМЕОДОМЕН
Особенно важным открытием было обнаружение в 3-х экзонах всех проклонированных гомеозисных генов (и большинства генов сегментации) высоконсервативной области ДНК из 180 пар оснований. Эту короткую последовательность В.Геринг назвал гомеобоксом. Соответствующая последовательность из 60 аминокислот в кодируемых этими генами белках, обогащенная аргинином и лизином, была обозначена термином гомеодомен. Гомеодомены являются составной частью белков-регуляторов транскрипции и принадлежат, следовательно, к числу активирующих транскрипцию факторов. Они характеризуются специфической структурой типа "спираль-поворот-спираль". Аминокислотные последовательности гомео-домена образуют три альфа-спирали, при этом первая и вторая расположены антипараллельно, а третья почти под прямым углом к ним. Вторая и третья спирали гомеодомена входят в состав НТН. Третья спираль, распознающая, взаимодействует с молекулой ДНК и располагается в ее большой бороздке. Аминокислотные остатки N-терминального конца гомеодомена "укладываются" в малой бороздке ДНК. Последняя спираль гомеодомена Аntennapediа представлена либо единой структурой, либо двумя, образующими слабый перегиб.
Гомеодомены могут быть включены в состав генов, как собранных в кластер, так и диспергированных по геному. В.Геринг разделил "кластерированные" гомеодомены дрозофилы на 6 классов, одним из которых является Аntеnnареdiа. Дисперсные гомеодомены подразделяют более чем на 16 классов. Показано, что гомеодомены имеют специфическое сродство к определенным участкам ДНК, которые очень часто характеризуются наличием последовательности ТААТ. Отмечено, что гомеодомены одного и того же типа могут взаимодействовать с разными сайтами связывания ДНК и, наоборот, разные гомеодомены могут взаимодействовать с одним и тем же сайтом связывания ДНК.
Гомеобокссодержащие гены (с повсеместным сохранением принципа колинеарности) были найдены практически у всех живых организмов (губок, гидры, пиявок, нематод и др.). Эти гены богато представлены и у млекопитающих. Гомеобокссодержащие гены, которые собраны у представителей этого класса животных в кластер, принято в настоящее время называть HOX-генами. Они особенно хорошо изучены у мыши и человека. В геноме млекопитающих обнаружено 38 НОХ-генов, собранных в 4 кластера. Кластеры занимают область примерно по 120 кб каждый и расположены в четырех разных хромосомах: у мыши — II, VI, XI, XV, у человека — II, VII, XII, XVII. Расположение генов в кластере в основном соответствует расположению гомологичных генов в хромосомах дрозофилы.
РОЛЬ ГОМЕОБОКССОДЕРЖАЩИХ ГЕНОВ В РАЗВИТИИ МЛЕКОПИТАЮЩИХ
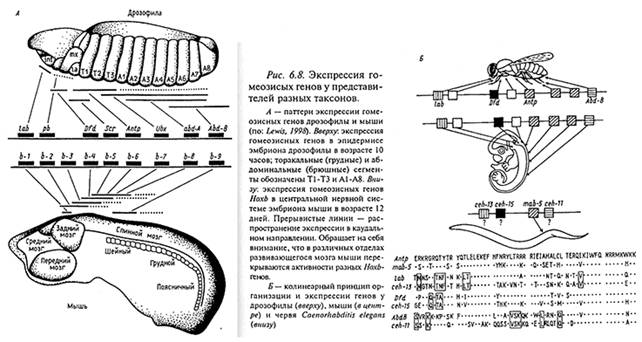 Прежде всего было выявлено, что у позвоночных, как и у дрозофилы, выполняется принцип коллинеарности, т.е. корреляции между положением генов в комплексе НОX-С и расположением зон их экспрессии вдоль оси тела. Так, у млекопитающих активность генов вдоль переднезадней оси тела зародыша соответствует последовательности этих генов в кластере: передние границы экспрессии генов Ноха-1 и НохЬ-1, расположенных в самом начале кластера (в 3-положении), находятся в передней области головы зародыша. Ген Ноха-5, расположенный в средней части кластера, экспрессируется в грудной области зародыша, Ноb-7— в туловище, а гены Ноха-9, Ноха-10, Ноха-11, Нохс1-12, замыкающие кластер, экспрессируются в задних отделах зародыша. В целом, зона, расположенная на заднем дистальном крае почки конечности, характеризуется экспрессией пяти Hох-генов: Ноха-9 — Ноха-13, далее следует зона, где экспрессируются четыре Яох-гена: Нохс!-9 — Ноха-12, далее идут полосы экспрессии трех (Нохс1-9 — Ноха-11), двух (Ноха-9 — Ноха-10) и, наконец, одного — Hох-9-генов.
Прежде всего было выявлено, что у позвоночных, как и у дрозофилы, выполняется принцип коллинеарности, т.е. корреляции между положением генов в комплексе НОX-С и расположением зон их экспрессии вдоль оси тела. Так, у млекопитающих активность генов вдоль переднезадней оси тела зародыша соответствует последовательности этих генов в кластере: передние границы экспрессии генов Ноха-1 и НохЬ-1, расположенных в самом начале кластера (в 3-положении), находятся в передней области головы зародыша. Ген Ноха-5, расположенный в средней части кластера, экспрессируется в грудной области зародыша, Ноb-7— в туловище, а гены Ноха-9, Ноха-10, Ноха-11, Нохс1-12, замыкающие кластер, экспрессируются в задних отделах зародыша. В целом, зона, расположенная на заднем дистальном крае почки конечности, характеризуется экспрессией пяти Hох-генов: Ноха-9 — Ноха-13, далее следует зона, где экспрессируются четыре Яох-гена: Нохс!-9 — Ноха-12, далее идут полосы экспрессии трех (Нохс1-9 — Ноха-11), двух (Ноха-9 — Ноха-10) и, наконец, одного — Hох-9-генов.
Как следует из рис., у млекопитающих в отличие от дрозофилы регионализация тех или иных структур обеспечивается скорее не специфической активностью отдельных гомеозисных генов, а перекрытием зон активности наборов этих генов, так что каждая обособленная область той или иной структуры обязана своим происхождением функционированию специфического, свойственного именно этой области набора гомеозисных генов.
ГОМЕОЗИСНЫЕ ГЕНЫ И МОРФОГЕНЕТИЧЕСКИЕ ПРОЦЕССЫ
Внимание к гомеозисным генам велико еще и потому, что они имеют прямое отношение к регуляции морфогенетических процессов. Характерной особенностью развития этого объекта является самостоятельная реализация двух различных программ — личиночной и имагинальной. Популяции клеток, которые дают начало личиночным и имагинальным тканям, разделяются очень рано — еще на стадии бластодермы. Клетки, формирующие впоследствии взрослую, имагинальную ткань, выделяются в специфические структуры, так называемые имагинальные диски.
Оказалось, что морфогенез заключается в образовании компартментов. Клеточные клоны, потомки отдельных клеток-предшественниц, объединяются и образуют поликлон. Появляется целый ряд поликлонов, которые формируют морфогенетические зоны. Эти зоны отделены друг от друга четкими границами и обозначаются термином компартмент. Ф.Крик и П.Лоуренс предположили, что гомеозисные гены как раз и ответственны за образование компартментов и являются своеобразными генами-селекторами, поскольку именно они "выбирают" путь развития клетки. Этот путь определяется бинарными решениями — к примеру, стать ли клетке передней или задней, дорсальной или вентральной, грудной или крыловой. Для образования 8 компартментов необходима активность трех селекторных генов, последовательно принимающих бинарные решения:
Все данные о гомеозисных генах, наводят на мысль, что существуют специфические регуляторные гены, которые при их активации "запускают" целый каскад структурных генов, обеспечивающих в конечном итоге дифференцировку различных тканевых и органных структур.
ГЕНЫ, КОНТРОЛИРУЮЩИЕ ФУНКЦИОНИРОВАНИЕ ГОМЕОЗИСНЫХ ГЕНОВ
Обнаружены две группы генов, контролирующих активность гомеозисных генов: Роlусоmb (РС) и группа trithorax (trx). РС репрессируют гомейозисные гены (нарушают их способность связываться с хроматином) или создают гетерохроматиновые структуры. Trx – активируют транскрипцию – формируют взаимодействующие с хроматином комплексы и модифицируют структуру хроматина. Гомейозисные гены содержат регуляторные элементы Pre (polycomb responsible elements), которые держат энхансеры в неактивном состоянии. У генов РС и trx идет конкуренция за один и тот же участок связывания.
Итак, дифференциальная транскрипция обеспечивается взаимодействием продуктов многих регуляторных генов, контролирующих в конечном итоге функциональное состояние соответствующих участков ДНК. Было выдвинуто также предположение о существовании как бы "суперрегуляторных" генов, способных запускать последовательные каскады генов, реализующих в конце концов программу специфической клеточной дифференцировки. Для таких генов выдающийся шведский цитолог Я.Э.Эдстрем еще в начале 60-х годов XX в. предложил термин "Master Genes" ("гены-господа"), а для контролируемых ими структурных генов соответственно — "Slaves Genes " ("гены-рабы").
Из опытов В.Геринга можно сделать ряд заключений.
1. Действительно существуют "гены-господа" и "гены-рабы".
2. Сложнейшая морфогенетическая реакция, завершающаяся формированием целого органа, может быть запущена одним, "главным" геном, который, следовательно, является ответственным за процессы морфогенеза, разрешая или запрещая (в случае мутации) целый сложный комплекс формообразовательных событий (Pax 6 – запускает развитие глаза у дрозофилы, позвоночных).
3. Морфогенетические процессы основываются на молекулярно-генетических событиях, специфика формы обусловливается спецификой последовательных ткане- и органоспецифических синтезов, разрешенных активацией "главного" гена.
4.Соответствующие молекулярно-морфогенетические системы являются высококонсервативными и обеспечиваются в высшей степени гомологичными молекулярно0-генетическими системами у самых разнообразных таксономических единиц.
5. Следует, очевидно, расстаться с широко распространенной идеей неких специфически биологических полей, берущей начало с Аристотеля и утверждающей, будто формообразовательные процессы детерминируются особыми факторами наподобие электромагнитных полей, лежащих вне развивающейся системы. Допускалось, таким образом, наличие двух независимых "сил", регулирующих процессы индивидуального развития — генетических, которые контролируют молекулярные процессы, и "механических" (биополе), которые контролируют становление формы организма.
В действительности оправдывается предсказание Т.Моргана, что развитие формы напрямую связано с функцией генов и со специфичностью их продуктов, из взаимодействия которых и складывается путь от специфики молекул к специфике формы. Следовательно, формообразовательные события зависят от внутренних процессов, а не от таинственных внешних сил. И совершенно очевидно, что ключевая роль в этих процессах принадлежит гомеозисным генам.
В то же время следует отметить, что феномен, обозначаемый как морфогенетическое поле, реально существует, но под таким полем следовало бы понимать равнодействующую межклеточных взаимодействий, которая имманентна самой развивающейся системе и так или иначе направляет формообразовательные перемещения клеточного материала. Вместе с тем специфика таких перемещений связана с особенностями адгезионных поверхностных клеточных белков, детерминирующих клеточное сродство (или, напротив, антагонизм), и в конечном итоге определяется активностью соответствующих генов.
Эволюция на основе гомеозисных генов (гипотеза)
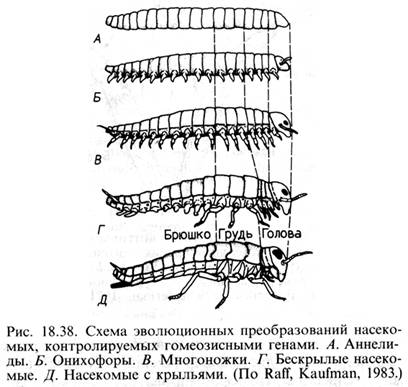 В конце 30-х гг. Рихард Гольдшмидт и Конрад Уоддингтон предположили, что гомеозисные гены являются тем звеном, которое связывает развитие, генетику и эволюцию. Уоддингтон подчеркивал, что гомеозисные гены представляют собой эмбриональные переключатели, способные регулировать направление дифференцировки. Гольдшмидт рассматривал гомеозисные гены как гены, вызывающие одновременно крупные изменения в развитии зародыша и, таким образом, играющие важную роль в эволюционном процессе. Внимание, которое уделяли гомеозисным генам Гольдшмидт и Уоддингтон, предвосхитило успехи науки наших дней. Используя делеционных мутантов дрозофилы, можно в общих чертах воспроизвести эволюционное становление этого вида. Полагают, что дрозофила как представитель отряда двукрылых происходит от насекомых с четырьмя крыльями (Д). Этот эволюционный шаг может быть генетически смоделирован делецией гена Ubх. Исследователи считают, что крылатые насекомые происходят от первичнобескрылых насекомых (В). Это можно представить как результат делеции гена Antennapedia, которая трансформирует все грудные сегменты в первый грудной сегмент, не имеющий крыльев. Бескрылые насекомые произошли от членистоногих, подобных многоножкам. Это имитирует делеция всего Bitorах -комплекса и соответствующая ей трансформация всех брюшных сегментов в грудные. И наконец, делеция обоих наборов гомеозисных генов (Ant и Btx-комплексов) приводит к состоянию примитивных членистоногих – онихофор (Б). Однако они предостерегают от буквальной интерпретации подобной аналогии, поскольку полная делеция даже одного комплекса Antp приводит к появлению не многоножки, а летального мутанта.
В конце 30-х гг. Рихард Гольдшмидт и Конрад Уоддингтон предположили, что гомеозисные гены являются тем звеном, которое связывает развитие, генетику и эволюцию. Уоддингтон подчеркивал, что гомеозисные гены представляют собой эмбриональные переключатели, способные регулировать направление дифференцировки. Гольдшмидт рассматривал гомеозисные гены как гены, вызывающие одновременно крупные изменения в развитии зародыша и, таким образом, играющие важную роль в эволюционном процессе. Внимание, которое уделяли гомеозисным генам Гольдшмидт и Уоддингтон, предвосхитило успехи науки наших дней. Используя делеционных мутантов дрозофилы, можно в общих чертах воспроизвести эволюционное становление этого вида. Полагают, что дрозофила как представитель отряда двукрылых происходит от насекомых с четырьмя крыльями (Д). Этот эволюционный шаг может быть генетически смоделирован делецией гена Ubх. Исследователи считают, что крылатые насекомые происходят от первичнобескрылых насекомых (В). Это можно представить как результат делеции гена Antennapedia, которая трансформирует все грудные сегменты в первый грудной сегмент, не имеющий крыльев. Бескрылые насекомые произошли от членистоногих, подобных многоножкам. Это имитирует делеция всего Bitorах -комплекса и соответствующая ей трансформация всех брюшных сегментов в грудные. И наконец, делеция обоих наборов гомеозисных генов (Ant и Btx-комплексов) приводит к состоянию примитивных членистоногих – онихофор (Б). Однако они предостерегают от буквальной интерпретации подобной аналогии, поскольку полная делеция даже одного комплекса Antp приводит к появлению не многоножки, а летального мутанта.
Тема № 9
Формирование пространственной
Организации
Гилберт С. Гл. 17, 18. Т. 3.
Корочкин Л.И. С. 84 -124, 154 -172.
Градиентные модели позиционной информации. Краткий обзор развития дрозофилы. Сегрегационные и гомеозисные гены. Гомеобокс и гомеодомен. Роль гомеобокссодержащих генов в развитии млекопитающих. Гомеозисные гены и морфогенетические процессы. Гены, контролирующие гомеозисные гены. Эволюция на основе гомеозисных генов.
Формирование пространственной организации - это результат функциональной активности эмбриональных клеток, посредством которой осуществляется упорядоченное расположение дифференцированных тканей. Каким образом в зародыше не только возникают различные типы клеток, но и создаются функциональные ткани и органы? Одно дело - дифференцировка хондроцитов или остеоцитов, синтезирующих соответственно хрящевой и костный матриксы, другое дело - образовывать эти клетки в такой пространственно-временной ориентации, которая приводит к созданию функциональной кости. При этом должна выполняться еще одна задача, а именно создаваться определенная кость, например плечевая, тазовая или бедренная. Способность клеток узнавать свое положение в пределах ограниченной популяции клеток и дифференцироваться в соответствии с получаемой ими позиционной информацией стала объектом интенсивного изучения.
Формирование пространственной организации включает реализацию выбора одного из возможных путей развития. Каким образом происходит так, что клетка, потенциально способная стать хрящевой или мышечной, становится мышечной, если располагается в одном месте, и хрящевой, если оказывается в другом?
|
|
|

Автоматическое растормаживание колес: Тормозные устройства колес предназначены для уменьшения длины пробега и улучшения маневрирования ВС при...

Механическое удерживание земляных масс: Механическое удерживание земляных масс на склоне обеспечивают контрфорсными сооружениями различных конструкций...

Папиллярные узоры пальцев рук - маркер спортивных способностей: дерматоглифические признаки формируются на 3-5 месяце беременности, не изменяются в течение жизни...

Адаптации растений и животных к жизни в горах: Большое значение для жизни организмов в горах имеют степень расчленения, крутизна и экспозиционные различия склонов...
© cyberpedia.su 2017-2024 - Не является автором материалов. Исключительное право сохранено за автором текста.
Если вы не хотите, чтобы данный материал был у нас на сайте, перейдите по ссылке: Нарушение авторских прав. Мы поможем в написании вашей работы!