

Индивидуальные и групповые автопоилки: для животных. Схемы и конструкции...

Типы оградительных сооружений в морском порту: По расположению оградительных сооружений в плане различают волноломы, обе оконечности...
Индивидуальные и групповые автопоилки: для животных. Схемы и конструкции...
Типы оградительных сооружений в морском порту: По расположению оградительных сооружений в плане различают волноломы, обе оконечности...
Топ:
Процедура выполнения команд. Рабочий цикл процессора: Функционирование процессора в основном состоит из повторяющихся рабочих циклов, каждый из которых соответствует...
Когда производится ограждение поезда, остановившегося на перегоне: Во всех случаях немедленно должно быть ограждено место препятствия для движения поездов на смежном пути двухпутного...
Генеалогическое древо Султанов Османской империи: Османские правители, вначале, будучи еще бейлербеями Анатолии, женились на дочерях византийских императоров...
Интересное:
Наиболее распространенные виды рака: Раковая опухоль — это самостоятельное новообразование, которое может возникнуть и от повышенного давления...
Принципы управления денежными потоками: одним из методов контроля за состоянием денежной наличности является...
Подходы к решению темы фильма: Существует три основных типа исторического фильма, имеющих между собой много общего...
Дисциплины:
|
из
5.00
|
Заказать работу |
|
|
|
|
Двигательная пластинка представляет собой функционально-анатомическую структуру, обеспечивающую связь окончания нервного волокна мотонейрона с мышечным волокном непосредственно. Она состоит из синапса, где электрический сигнал, исходящий из нервного волокна, изменяется на химический мессенджер (ацетилхолин), который в свою очередь вызывает другой электрический сигнал в клеточной мембране (сарколемма) мышечного волокна.
Зона концевой двигательной пластинки является территорией, где происходит иннервация мышечных волокон. В настоящее время этот район называют двигательной точкой [153]. Клинически каждая двигательная точка определяется областью, где видимые или пальпируемые мышцы дают локальную судорожную реакцию в ответ на минимальное поверхностное раздражение электричеством (стимуляция). Первоначально двигательную точку ошибочно представляли как зону вхождения нерва в мышцы [4].
Местоположение концевых двигательных пластинок
Точное представление о местоположении концевых двигательных пластинок крайне важно для постановки правильного клинического диагноза и лечения миофасциальных триггерных точек. Если, как это часто бывает у больного, патофизиология триггерных точек тесно ассоциируется с концевыми пластинками, можно ожидать, что миофасциальные триггерные точки располагаются только там, где находятся концевые двигательные пластинки. Почти во всех скелетных мышцах концевые двигательные пластинки располагаются почти по середине каждого волокна, т. е. на середине расстояния между точками их прикрепления. Этот принцип, характеризующий мышцы человека, представлен схематически Coёrs и Woolf [44], одними из первых исследовавших концевые двигательные пластинки (рис. 2.8). Aquilonius и соавт. [5] представили результаты подробного анализа местонахождения концевых двигательных пластинок двуглавой мышцы и плеча, передней большеберцовой и портняжной мышц взрослого человека.
|
|
Christensen [36] описал распределение срединных концевых двигательных пластинок у мертворожденного в следующих мышцах: мышце, противопоставляющей большой палец, плечелучевой, полусухожильной (два поперечных пучка концевых пластинок), двуглавой мышце плеча, тонкой (два определённых типа уплотнения мышечного волокна внутри каждой двигательной единицы), портняжной (разбросанные концевые пластинки), трёхглавой мышце плеча, икроножной, передней большеберцовой, мышце, противопоставляющей V палец кисти, прямой мышце бедра, коротком разгибателе пальцев стоп, перстнещитовидной и дельтовидной.

Рис. 2.8. Расположение концевых двигательных пластинок в скелетных мышцах различной структуры.
Красные линии представляют мышечные волокна;
чёрные точки показывают концевые двигательные пластинки этих волокон,
а чёрные линии обозначают прикрепление волокон к апоневрозу.
Концевые двигательные пластинки обнаруживаются в средней части каждого мышечного волокна.
а — линейные концевые двигательные пластинки, находящиеся в мышце с короткими волокнами, расположенные между параллельными апоневрозами, как это наблюдается в икроножной мышце;
б — петлеобразное расположение концевых пластинок в двуперистой мышце (например, m.flexor carpi radialis и m.patmaris longus;
в — синусоидное расположение концевых пластинок в мышечных волокнах средней части дельтовидной мышцы, характеризующихся сложной перистой конфигурацией. (Из Coёrs С. Contribution а létude de la jonction neuromusculaire. II Topographie zonale de l'innervation motrice terminale dans les muscles striés. Arch. Biol. Paris 64, 495—505, 1953 [42], адаптировано с разрешения.)
Как было сказано выше, принцип используется вне зависимости от строения мышечных волокон. Для этой цели важно знать, как расположены мышечные волокна: это поможет понять, как расположены концевые пластинки внутри каждой мышцы и, следовательно, определить место, где следует искать триггерные точки.
|
|
В мышце волокна могут располагаться следующим образом: параллельно, параллельно с сухожильными вставками, веретенообразно, веретенообразно с двумя брюшками. Мышцы также могут быть одноперистыми, двуперистыми, многоперистыми, обладать спиральным расположением волокон (рис. 2.9).
Рис. 2.9. Параллельное и веретенообразное расположение мышечных волокон обеспечивает большее изменение длины при затрате силы. Перистое строение обеспечивает большую силу при издержках в длине. Обратите внимание на то, что расположение мышечных волокон 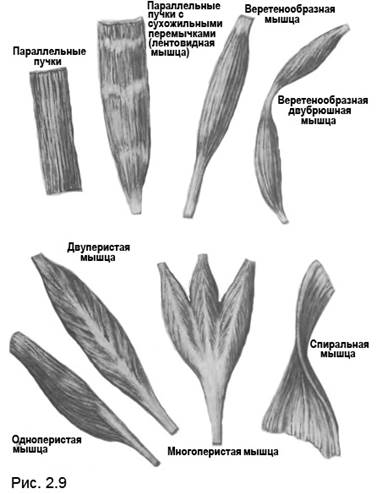 в каждой отдельной мышце обеспечивает почти равную длину всех составляющих её мышечных волокон.
в каждой отдельной мышце обеспечивает почти равную длину всех составляющих её мышечных волокон.
На рис. 2.8 можно видеть расположение концевых двигательных пластинок в мышцах разной формы. (Из Clemente С. D. Gray's Anatomy of the Human Body. 30th ed. Philadelphia: Lea & Fibiger, 1985, 429, с разрешения, адаптировано)

Рис. 2.10. Микрофотографии и рисунки, показывающие расположение концевых пластинок в скелетных мышцах мыши (по результатам исследования Schwarzacher, использовавшего окрашивание на холинэстеразу по Koelle в модификации Соёrs, чтобы показать концевые двигательные пластинки [231].
На схемах, выполненных с использованием компьютера (в, д, е),
красные линии означают мышечные волокна;
чёрными точками представлены концевые двигательные пластинки этих мышечных волокон,
а чёрные линии изображают прикрепления мышечных волокон либо непосредственно к кости, либо к апоневрозу.
а — микрофотография,
б — опубликованный схематический рисунок, выполненный с M.gracillis posterior;
в — компьютерная версия рис. б для сравнения. Видно два скопления концевых пластинок;
г — микрофотография диафрагмы, видна зона концевых пластинок, проходящая между концами мышечных волокон;
д — схематическое изображение расположения концевых пластинок в полусухожильной мышце;
е — в большой ягодичной мышце. (Из Schwarzacher V. H. Zurlage der motorischen endplallen in den skeletmuskeln. Acta Anat 30, 758—774, 1957, с разрешения. Схематические изображения получены из этого же источника.)
Рис. 2.11. Схематическое изображение двух концевых двигательных пластинок млекопитающих и нервно-сосудистых пучков, ассоциированных с ними.
|
|

Нервные окончания двигательного аксона закрываются внутри компактного мионеврального соединения, погружённого внутрь слегка приподнятой области концевой пластинки в мышечном волокне.
Волокна двигательного нерва сопровождают чувствительные нервные волокна и кровеносные сосуды.
Вегетативные нервы находятся в тесной взаимосвязи с этими мелкими кровеносными сосудами, расположенными в мышечной ткани.
Пиковые потенциалы, зарегистрированные на уровне области концевой пластинки мышечного волокна, показывают отрицательное первоначальное угасание.
На очень небольшом расстоянии в обе стороны от концевом пластинки, справа, пиковые потенциалы этого волокна обладают положительным первоначальным угасанием.
Это один из путей, с помощью которого осуществляется электромиографический поиск концевых двигательных пластинок. Конфигурация пиковых потенциалов внизу рисунка соответствует форме волны, которая могла бы регистрироваться в разных местах вдоль передней плоскости мышечного волокна. (Из рис.5 Salpeter М.М. Vertebral neuromuscular junctions: General morphology, molecular organization, and functional consequences. In: Salpeter M. M., Ed. The Vertebrate Neuromuscular Junction. New York: Alan R. Liss, Inc. 1987: 1—54 [225], с разрешения, адаптировано.)
Среди скелетных мышц имеется по крайней мере четыре рода исключений из правила, что концевая пластинка может располагаться только в середине брюшка мышцы.
1. В некоторых мышцах человека, включая мышцу живота, полуостистую мышцу головы и полусухожильную мышцу, есть перемычки, делящие мышцы на серию сегментов, каждый из которых обладает своей собственной зоной расположения концевых пластинок, что показано на примере мышц грызунов (рис. 2.10, а, б, в, д). Сравните с рис. 2.10, г, е, иллюстрирующими обычное построение элементов концевой пластинки.
2. В портняжной мышце человека концевые двигательные пластинки разбросаны по всей мышце. Эти концевые пластинки обеспечивают параллельные пучки из укороченных волокон, которые могут переплетаться друг с другом по всей длине. При этом хорошо определяемой зоны концевых пластинок может и не быть [44]. По мнению Christensen [36], нежная мышца человека обладает двумя поперечно расположенными зонами, содержащими концевые пластинки, подобно полуперепончатой мышце, но к тому же снабжёнными переплетающимися волокнами с разбросанными концевыми пластинками, как у портняжной мышцы [44]. Такая переплетающаяся конфигурация волокон необычна для скелетных мышц человека, а строение концевой пластинки в обеих указанных мышцах может различаться у разных индивидов.
|
|
3. Внутри мышцы наблюдается разделение на ячейки и отделы (компартментализация), причём, и это очень важно, каждая ячейка или футляр изолированы фасциальной оболочкой.
Отдельная веточка двигательного нерва иннервирует зону расположения каждой концевой двигательной пластинки или каждый футляр. Каждый такой анатомо-физиологический отдел обладает определённой функцией. В качестве примера можно привести проксимальную и дистальную части лучевого длинного разгибателя кисти и дистального лучевого сгибателя кисти.
Жевательная мышца также представляет собой наглядное свидетельство разделения на ячейки и футляры (компартментализация) двигательной единицы [185]. С этой точки зрения изучено относительно небольшое число мышц человека, однако, вероятно, это общий признак мышц.
4. Икроножная мышца представляет собой особый пример аранжировки мышечных волокон, которые увеличивают мышечную силу путём уменьшения объёма подвижности. Волокна искривляются под значительным углом так, что одно мышечное волокно представляется минимальной долей от общей длины мышцы. Следовательно, зона концевой пластинки проходит центрально вниз по наибольшей длине каждого участка мышцы. Пример такого строения приведён на рис. 2.8 а.
На рис. 2.11 схематично изображены две концевые пластинки и маленький нейрососудистый пучок, который пересекает мышечные волокна в местах, где терминальные аксоны снабжают двигательные концевые пластинки [60]. Линейное расположение концевых пластинок, которые идут по ходу нейрососудистого пучка, ориентировано поперёк направления мышечных волокон [5, 44]. Нейрососудистый пучок включает болевые рецепторы чувствительных нервов и вегетативные нервы, тесно связанные с сопровождающими их сосудами. Непосредственное соприкосновение этих структур с двигательными концевыми пластинками является исключительно важным для представления и понимания процесса происхождения боли и вегетативных феноменов, сочетанных с миофасциальными триггерными точками.
Нейромышечное соединение
У разных видов топографическое расположение нервных окончаний на уровне концевых двигательных пластинок различное. Так, у лягушки обнаружили расширенные синаптические желобовидные канавки. У крыс и мышей желобовидные канавки бывают извитыми или свёрнутыми в виде спирали так, как это показано на рис. 2.11. На рис. 2.12 представлено расположение нервных окончаний у человека.
|
|
При окрашивании концевой пластинки на холинэст-разу (см. рис. 2.12, а) чётко видны более или менее отделённые друг от друга группы синаптических щелей. Благодаря достаточному разделению эта структура может эффективно функционировать в качестве многочисленных отдельных синапсов, которые могли бы отвечать за сложные серии пиковых потенциалов, исходящих из активного локуса, расположенного в мышечном волокне (см. раздел Г).
На рис. 2.12, б схематично представлено расположение концевых пластинок в мышечных волокнах у человека (поперечное сечение).

Рис. 2.12. Строение концевой двигательной пластинки. Микрофотография субневрального аппарата и схема поперечного разреза нервного окончания в мышце человека.
а — на микрофотографии области концевой пластинки человека, окрашенной по модифицированному методу Koelle для выявления присутствия холинэстеразы, видны многочисленные группы разрозненных (дискретных) синаптических щелей в субневральном аппарате.
Такое нервное окончание двигательного нерва одной концевой пластинки состоит из 11 отдельных округлых или овальных пар. Эта структурная форма отличается от извилистых и искривлённых, сетчатых окончаний, встречающихся у крыс и мышей. (Из Cёrs С. Structural organization of the motor nerve endings in mammalian muscle spindles and other striated muscle fibers. In: Bouman HD, Woolf AL, eds. Innervation of Muscle. Baltimore: Williams & Wilkins, 1960, 40—49, с разрешения [43];
б — схема поперечного сечения через область концевой двигательной пластинки. На этом немиелинизированном нервном окончании видно шесть расширений (чёрные дольки). Каждое расширение имеет свою собственную синап-тическую канавку и систему из постсинаптических складок. Пунктирные линии представляют расширение шванновских клеток, прикреплённых сарколеммной мембраной мышечной клетки и изолирующих содержимое синаптической щели от внеклеточной среды.
Вертикальные параллельные линии означают испещрённость (Z-линии) мышечного волокна. (Из Coёrs С. Contribution а l'étude de la jonction neuromusculaire. Donnés nouvelles concernant la structure de l'arborosation terminale et de l'appareil sousneural chez l'homme. Arch. Biol. Paris 64, 133—147, 1953 [41], с разрешения.)

Рис. 2.13. Схема поперечного сечения части нейромышечного соединения, которое передаёт нервные потенциалы действия через синапсы путём химической передачи, после чего они становятся мышечным потенциалом действия. В ответ на распространение потенциала действия вниз по двигательному нерву синаптическая мембрана нервного окончания раскрывает «входные ворота» для прохождения электрического напряжения по кольцевым канальцам, делая возможным приток кальция из синаптической щели (маленькое направленные вверх красные стрелки). Кальций вызывает высвобождение многочисленных порций ацетилхолина внутри синаптической щели (большие направленные вниз стрелки).
Рецепторы, специфичные для ацетилхолина, деполяризуют постсинаптическую мембрану мышечного волокна в такой степени, чтобы открылись натриевые канальцы в глубине складок постсинаптической мембраны. Достаточная деполяризация этих натриевых канальцев инициирует распространение потенциала действия в мышечном волокне.
Нейромышечное соединение является синапсом, который, подобно многим другим структурам в центральной нервной системе, зависит от ацетилхолина в качестве нейротрансмиттера (передатчика).
Основная структура и функция нейромышечного соединения схематично представлены на рис. 2.13. Нервное окончание продуцирует ацетилхолин. При этом потребляется энергия, которую в достаточном количестве поставляют митохондрии, находящиеся в нервных окончаниях.
Нервное окончание реагирует на прибытие активного потенциала из α-мотонейрона путём раскрытия ионных кальциевых каналов. По этим каналам ионизированный кальций продвигается от синаптической щели внутрь нервного окончания. Эти канальцы располагаются по обеим сторонам специализированного участка нервной мембраны, из которой в норме в ответ на присутствие ионизированного кальция высвобождаются порции ацетилхолина.
Одновременное высвобождение множества порций ацетилхолина позволяет быстро преодолеть барьер холинэстеразы в синаптической щели. Большая часть ацетилхолина затем пересекает синаптическую щель, чтобы достичь перекреста складок постсинаптической мембраны мышечного волокна, где располагаются ацетилхолиновые рецепторы (см. рис. 2.13). Однако, вскоре холинэстераза разрушает остатки ацетилхолина, ограничивая время его действия. Теперь синапс становится способным немедленно отвечать на другой потенциал действия.
Нормальное произвольное высвобождение отдельных порций ацетилхолина из нервного окончания вырабатывает изолированные индивидуальные миниатюрные потенциалы концевых пластинок. Такие индивидуальные миниатюрные потенциалы концевых двигательных пластинок не распространяются и вскоре исчезают. С другой стороны, массовое высвобождение ацетилхолина из многочисленных пузырьков в ответ на потенциал действия, возникающий в нервном окончании, деполяризует постсинаптическую мембрану в достаточной мере, чтобы достичь порога его возбуждения. Это событие вызывает потенциал действия, который передаётся поверхностной мембраной (сарколемма) по мышечному волокну.
__________________________________________________________________________________
Мышечная боль
Современное представление о нейрофизиологии мышечной боли — достаточно сложный вопрос, чтобы посвятить его обсуждению отдельные книги [188, 189, 190а, 191, 240].
Схематично известно, что некоторые эндогенные субстанции могут повышать чувствительность мышечных болевых рецепторов (ноцицепторов), т. е. сенсибилизировать их. К ним относятся брадикинин, простагландин Е и 5-окситриптамин, которые в комбинации могут усиливать эффекты сенсибилизации. Освобождение простагландинов из соседних симпатических волокон с помощью норадреналина может оказывать влияние на механизм функции миофасциальной точки на уровне концевой пластинки.
Имеется очевидное доказательство того, что повышенная чувствительность болевых рецепторов, вызываемая индукцией простагландинов, усиливается циклическим аденозинмонофосфатом (цАМФ). Активность других факторов, способных повышать местную чувствительность, возрастает при повышении концентрации ионов водорода (рН снижается до 6,1) и субстанции Р [188].
Периферическое повышение чувствительности болевых рецепторов ответственно за местную болезненность при надавливании на триггерную точку и, по-видимому, за отражённую боль. Какие из этих или других субстанций являются ответственными за повышение чувствительности болевых рецепторов в районе активных локусов миофасциальных триггерных точек, в настоящее время неизвестно, но это издание «Руководства» предлагает широкое поле для последующего плодотворного научного поиска и углублённого исследования вопроса, включая и лекарственную терапию.
Некоторые феномены, возникающие на уровне спинного мозга, могут относиться к определённым типам отражённой боли. Экспериментальный мониторинг ответов с клеток задних рогов спинного мозга на стимуляцию глубоких тканей (включая мышцы) может помочь оценить местонахождение и протяжённость рецептивных полей, нейрона.
Инъекции агентов, вызывающих болевые ощущения в мышечные рецептивные поля нейрона, могут привести к появлению в этой конечности дополнительных рецептивных полей [121]. Такой феномен относится к «пробуждающимся» или к «спящим» болевым рецепторным (ноцицептивным) полям в спинном мозге. Чувствительность только оригинальных ноцицепторов боли меток задних рогов спинного мозга может возрасти в достаточной мере, чтобы сделать их ответственными за наиболее нежные неноцецептивные стимулы. Сходные феномены могут наблюдаться тогда, когда повреждающий агент вводится в другую мышцу на этой же конечности, но вне первоначального (оригинального) рецептивного поля [121, 190].
Входные импульсы из некоторых тканей в один чувствительный поясничный спинальный нейрон встречаются часто. В экспериментах на кошках [95], большая часть из 188 изученных единиц (77 %) были гиперконвергентными и отвечали на ноцицептивный входной сигнал, поступающий из двух или более глубоко расположенных тканей: суставных фасеток, надкостницы, связок, межпозвоночных дисков, твёрдой мозговой оболочки спинного мозга, поясничных, бедренных и проксимальных концов мышцы и сухожилий нижних конечностей. Большая часть таких единиц (93 %) также обладала чувствительными болевыми зонами в кожных покровах [95]. Это не противоречит клиническим наблюдениям, согласно которым поясничную боль и боль, исходящую из тканей нижних конечностей, нельзя достаточно чётко локализовать или отнести к определённой ткани, не имея дополнительной информации.
Подавляющее большинство клеток задних рогов спинного мозга, которые обладают висцеральным входом, также имеет соматический вход, являющийся ноцицептивным [33]. При более углублённом понимании феномена отражённой боли как с клинической, так и с нейрофизиологической точки зрения становится очевидным, что ощущаемая пациентом боль, скорее всею, является отражённой не из того участка, который стал причиной первоначальной жалобы на боль. Представляется маловероятным, что разветвление первичных афферентных волокон может являться ответственным за возникновение отражённой боли, за редким исключением [190].
Распознавание неврологических изменений [296] в центральной нервной системе представляет собой относительно новую фундаментальную разработку с глубоким клиническим смыслом. Остро возникающий болевой разряд может усиливать пролонгированные изменения в выработке болевых сигналов, поступающих в центральную нервную систему, что неминуемо приводит к функциональным н структурным изменениям в чувствительной и двигательной функции человека. Нейрофизиологическое доказательство «взвинчивания» невральной активности было недавно представлено Yaksh и Abram [295]. Более пролонгированный болевой разряд может усиливать существующие в течение продолжительного периода времени изменения, которые не могут быть обратимыми только за счёт фактора времени.
Yu и соавт. [296] продемонстрировали нейропластическую сенсибилизацию чувствительных нервов. Они сообщили о повышении их восприимчивости к стимулированию в кожных и глубоких рецептивных полях мышцы путём обкалывания временным повреждающим (болезненным) агентом, например горчичным маслом, вводимым к мышцу языка крыс, находившихся под наркозом. Повреждающая стимуляция одной мышцы повышала восприимчивость другой мышцы на такое раздражение.
Большинство страданий, возникающих из-за хронической боли, вполне можно предотвратить, если быстро и эффективно контролировать острую боль. Клинические исследования в этой области в настоящее время быстро расширяются. Что касается миофасциальных триггерных точек, следует сказать, что Hong н Simmons [127] показали, что продолжительность лечения, требующегося пациенту, у которого развился синдром миофасциальной триггерной точки в большой грудной мышце вследствие хлыстовой травмы, непосредственно зависит от продолжительности времени между эпизодом несчастного случая и началом лечения. Чем дольше откладывалось лечение, тем сложнее оно будет, а вероятность освобождения от симптомов заболевания заметно снижается.
Местное обезболивание во время выполнения хирургической операции, направленное на предотвращение проникновения ноцицептивных сигналов в спинной мозг, является полезным вмешательством [295], но оно становится более эффективным, если наряду с этим осуществляется тщательный контроль послеоперационной боли. Концепция превентивного местного обезболивания успешно внедряется путём блокирования триггерной болевой точки и ещё до того, как её будут обкалывать [76, 78]. Katz и соавт. [148] показали, что предотвращение острой хирургической боли в свою очередь предотвращало переход острой боли в хроническую. Они также отметили прямую зависимость между степенью тяжести острой послеоперационной боли и степенью интенсивности последующей хронической боли после операции.
Современные исследования показали, что в ответ на экспериментально вызванную острую боль и на хроническую нейрогенную боль активируются разные области головного мозга [130]. С помощью позитронной эмиссионной томографии установлено, что при нейрогенной боли происходит ударная активация преимущественно правой сингулярной части коры головного мозга (поле 24 по Бродманну), вне зависимости от стороны болезненной мононевропатии (Привет вопросам «ах, почему же сколиоз преимущественно правосторонний» — H.B.). Активация этой области головного мозга ассоциируется с эмоциональным расстройством, вызываемым тем или иным заболеванием.
При острой боли активируются двигательные и чувствительные участки коры головного мозга, порождая скорее когнитивный (познавательный) и двигательный опыт поведения, чем эмоциональное чувство. Эти данные нейрофизиологического исследования подчёркивают особо значение афферентного мотивированного параметра хронического нейрогенного болевого паттерна, не задействованного при острой боли.
Хроническая боль приводит к мучительному страданию, однако при этом возбуждение в головном мозге происходит иначе, чем в случае острой боли. Такие нейрофизиологические наблюдения особо подчёркивают, что как для каждого больного, так и для всей системы здравоохранения в целом крайне важна профилактика хронической боли. Вновь активированные миофасциальные триггерные точки, не выявленные или плохо леченные, становятся главной причиной хронической боли.
|
|
|

Адаптации растений и животных к жизни в горах: Большое значение для жизни организмов в горах имеют степень расчленения, крутизна и экспозиционные различия склонов...

Типы сооружений для обработки осадков: Септиками называются сооружения, в которых одновременно происходят осветление сточной жидкости...

История развития хранилищ для нефти: Первые склады нефти появились в XVII веке. Они представляли собой землянные ямы-амбара глубиной 4…5 м...

История создания датчика движения: Первый прибор для обнаружения движения был изобретен немецким физиком Генрихом Герцем...
© cyberpedia.su 2017-2024 - Не является автором материалов. Исключительное право сохранено за автором текста.
Если вы не хотите, чтобы данный материал был у нас на сайте, перейдите по ссылке: Нарушение авторских прав. Мы поможем в написании вашей работы!