Вопросы к итогу №1
Понятие «биомембрана»
МЕМБРА́НА, -ы, ж. от лат. membrana — кожица, перепонка.
Мембрана - упругая перепонка, тонкая пленка или пластинка, способная совершать колебания. Гибкая тонкая пленка, находящаяся в состоянии натяжения и поэтому обладающая упругостью.
Принцип построения всех биологических мембран одинаков, независимо от того, какой клетки (растительной или животной) или клеточной органелле она принадлежит.
Под мембраной чаще подразумевают перегородку, разграничивающую пространства, компартменты. Считается, что плазматическая мембрана вместе с ядром и цитоплазмой является одним из основных компонентов клетки.
Ядро и органеллы клетки также отделены от цитоплазмы мембранами, которые предупреждают свободное движение воды и растворенных в ней веществ из цитоплазмы в них и наоборот. Это создает условия для разделения биохимических процессов, протекающих в различных компартментах внутри клетки.
Гук действительно был первым человеком, увидевшим клетки. В 1663 году, пытаясь понять, почему пробковое дерево так хорошо плавает, Гук стал рассматривать тонкие срезы пробки с помощью усовершенствованного им микроскопа. Он обнаружил, что пробка разделена на множество крошечных ячеек, напомнивших ему монастырские кельи, и он назвал эти ячейки клетками. Р.Гук видел оболочку, стенку, образованную целлюлозой. Но не биомембрану.
Физические модели
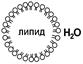
Различают 2 основных типа – плоскую и сферическую (липосому).
Плоскую бислойную липидную мембрану предложил в 1962 г. Мюллер. Она представляет собой саморганизующийся бислой фосфолипидов в отверстии тефлоновой пластинки.
Слева: Тефлоновый стакан (3) с отверстием (7), закрытым бислоем фосфолипидов. Сам стакан, содержащий водный раствор какого-либо вещества, помещён в другой сосуд (1), содержащий также водный раствор (5). С помощью электродов 2 и 4 можно проводить электрофизиологические измерения.
 Справа: Окно (7) при увеличении: фосфолипиды образовали плоскую модельную мембрану.
Справа: Окно (7) при увеличении: фосфолипиды образовали плоскую модельную мембрану.
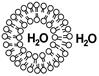
 Пример сферической модели биомембраны — липосомы - Пузырек в водном растворе (полярном растворителе) образованный бислоем фосфолипидов, содержащий внутри водный раствор.
Пример сферической модели биомембраны — липосомы - Пузырек в водном растворе (полярном растворителе) образованный бислоем фосфолипидов, содержащий внутри водный раствор.
Мицелла не может являться моделью биомембраны!!!
Биологические модели биомемраны
Это мембраны живых клеток. Наиболее часто в эксперименте используют тени эритроцитов и гигантский аксон кальмара.
Мембранные липиды
Липиды клеточных структур эукариотических клеток представлены 3 основными группами:
· фосфолипиды,
· гликолипиды
· стероиды.
Фосфолипиды
 Фосфолипиды - основа биомембран.
Фосфолипиды - основа биомембран.
Фосфолипи́ды — сложные липиды, в которых гидрофобная липидная часть молекулы соединена с гидрофильными остатками фосфорной кислоты.
Молекулы, одна часть которых является гидрофобной, а другая – гидрофильной называются амфифильными (Амфи — приставка, означающая: «вокруг», «с обеих сторон», - любить, иметь склонность)
C водной средой взаимодействует полярная часть молекулы. У фосфолипидов гидрофобным является довольно длинный неполярный "хвост".. Фосфолипиды подразделяются на 2 группы:
· глицерофосфолипиды (производные фосфатидной кислоты)
· сфингофосфолипиды (производные церамида, сфингомиелины).
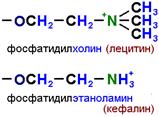 Глицерофосфолипиды
Глицерофосфолипиды
Глицерофосфолипиды представляют собой производные фосфатидной кислоты
Классификация фосфолипидов осуществляется
по структуре полярных радикалов.
Основными липидами мембран животных клеток являются глицерофосфолипиды: фосфатидилхолин (лецитин) и фосфатидилэтаноламин (кефалин).
Фосфолипиды активно используются в фармацевтической и «парафармацевтической» практике с различными целями, в том числе как средство для улучшения состояния кожи и похудения.
Сфингофосфолипиды
Наиболее распространенный сфингофосфолипид – сфингомиелин. Сфингомиелин содержится в больших количествах в белом веществе мозга, в миелиновых оболочках нервных стволов.
Мембранные белки
К мембранным белкам относятся белки, которые встроены в клеточную мембрану или мембрану клеточной органеллы. Около 25% всех белков являются мембранными.
Биохимическая классификация
По биохимической классификации мембранные белки делятся на интегральные и периферические.
Интегральные мембранные белки прочно встроены в мембрану и могут быть извлечены из липидного окружения только с помощью детергентов или неполярных растворителей. По отношению к липидному бислою интегральные белки могут быть трансмембранными политопическими или интегральными монотопическими.
Периферические мембранные белки являются монотопическими белками. Они либо связаны слабыми связями с липидной мембраной, либо ассоциируют с интегральными белками за счёт гидрофобных, электростатических или других нековалентных сил. Таким образом, в отличие от интегральных белков они диссоциируют от мембраны при обработке соответствующим водным раствором. Эта диссоциация не требует разрушения мембраны.
Свойства мембранных белков
В структуре многих мембранных белков, как правило, четко различаются участки, ответственные за их биологическую активность. Очень часто биологически активный участок состоит преимущественно из полярных аминокислот. Поэтому вероятно, что полярная часть мембранного обеспечивает функциональную активность белка, а неполярная часть обеспечивает структурную устойчивость молекулы. Однако такое правило не является универсальным. Так, белки, превращающие гидрофобные субстраты имеют гидрофобные карманы, концентрирующие молекулы субстрата для его ферментативной модификации.
Белки-ферменты
Наиболее распространены среди всех мембранных белков. В их число входят как интегральные, так и белки.
Фермент взаимодействует с субстратом небольшим участком своей поверхности – активным центром. Ферменты входят в состав как плазматических, так и внутриклеточных мембран.
Рецепторные белки
Рецепторными называют белки, специфически связывающие те или иные низкомолекулярные вещества. При связывании специфических лигандов рецепторные белки обратимо меняют свою форму. Эти изменения запускают внутри клетки ответные химические реакции. Таким способом клетка воспринимает различные сигналы, поступающие из внешней среды, и отвечает на них.
Белки-рецепторы и белки, определяющие иммунную реакцию клетки, – антигены, также могут быть как интегральными, так и периферическими компонентами мембраны.
Углеводы мембран
Углеводы, в составе мембран обнаруживаются лишь в соединении с белками (гликопротеины и протеогликаны) и липидами (гликолипиды).
В числе углеводных компонентов – глюкоза, галактоза, нейраминовая кислота, фукоза и манноза.
Углеводные компоненты мембранных структур в подавляяющем большинстве открываются во внеклеточную среду.
Их функции связаны с контролем за межклеточными взаимодействиями, поддержанием иммунного статуса клетки.
Типичным примером гликоконъюгатов, выполняющих свои функции в составе мембран, являются антигенные детерминанты эритроцитов различных групп крови. Они представлены как гликолипидами, так и гликопротеинами, в числе которых – белок гликофорин.
Очень важна роль углеводного компонента белковых молекул в формировании специфических функций мембранных белков и липидов. Многие белковые молекулы, особенно биологически активные вещества, синтезируются в виде крупных, неактивных предшественников, которые затем расщепляются специфическими протеазами с формированием «зрелых» биологически активных продуктов.
Деятельность протеаз контролируется уровнем гликозилирования белков. Так, многие белки, синтезируемые вначале как гликопротеины, в дальнейшем в результате процессинга теряют олигосахаридную часть.
Надмембранные структуры
· Собственно надмембранный комплекс, или гликокаликс. В его состав входят периферические белки мембраны, углеводные части гликолипидов и гликопротеинов. Гликокаликс играет важную роль в рецепторной функци. В состав гликокаликса входят полисахаридные цепочки мембранных интегральных белков – гликопротеидов. Они содержат такие углеводы, как манноза, глюкоза. Углеводные гетерополимеры гликокаликса образуют ветвящиеся цепочки, между которыми располагаются свободные гликолипиды и протеогликаны. Слой гликокаликса имеет желеобразную консистенцию, что значительно снижает диффузию различных веществ из клетки. Здесь же могут накапливаться выделенные клеткой ферменты, участвующие во внеклеточное пищеварении.
· Производные надмембранных структур. К ним относятся специфические химические соединения, не производящиеся самой клеткой. Наиболее изучены они на микроворсинках клеток кишечного эпителия млекопитающих. Здесь ими являются гидролитические ферменты, адсорбирующиеся из полости кишки. Их переход из взвешенного в фиксированное состояние создает базу для качественно иного типа пищеварения.
Как это было сделано?
Умертвили участок нерва. При этом электротоническое проведение тока на поврежденном участке сохранилось, а возбудимость этого участка исчезла. Один из электродов установили на поврежденном участке, а второй — на неповрежденный.
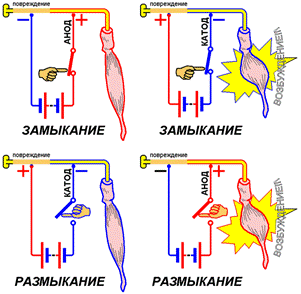
Если с неповрежденным участком соприкасается катод, возбуждение возникало в момент замыкания тока. Если же катод устанавливали на поврежденном участке, а анод — на неповрежденном, возбуждение возникает только при размыкании тока.
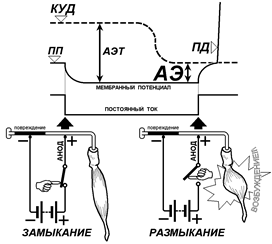
Анодно-размыкательное возбуждение. КУД – критический уровень деполяризации, ПП – потенциал покоя, ПД – потенциал действия, АЭТ – анэлектротон, АЭ – анодическая экзальтация. Объяснение в тексте.
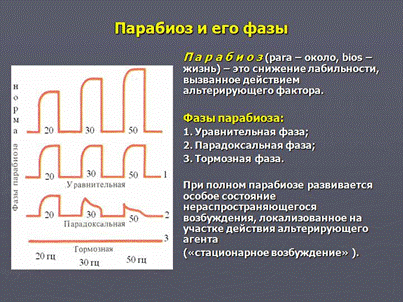
Парабиоз Н.Е.Введенского
Н. Е. Введенский изучал явление парабиоза на нервах, мышцах, железах, спинном мозге и пришел к выводу о том, что
парабиоз - это общая, универсальная реакция возбудимых тканей на сильное или длительное воздействие.
Сущность парабиоза состоит в том, что под влиянием раздражителя в возбудимых тканях изменяются их физиологические свойства, в первую очередь резко снижается лабильность.
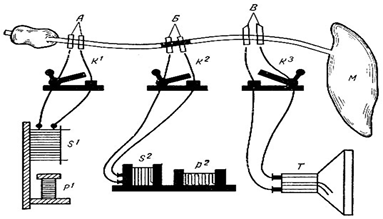
Опыт
3 элкутрона – нанесение,отверденя биотоков
Нерв на небольшом участке подвергали повреждению (альтерация) химическими веществами (кокаин, хлороформ, фенол, хлорид калия), сильным фарадическим током, механическим фактором.
Затем наносили раздражение электрическим током на альтерированный участок нерва или же выше его. Таким образом, импульсы должны были или возникать в альтерированном отрезке нерва, или проходить через него на своем пути к мышце. Сокращение мышцы свидетельствовало о проведении возбуждения по нерву.
Распространение электротона
Строго говоря, мы должны говорить не о распространения электротона, а о распространении локального ответа, даже точнее о локальной деполяризации, включающей как пассивный (физический электротон), так и активный (собственно локальный ответ) компоненты. Однако суть процессов местной деполяризации от этого не изменится и без ущерба истине мы будем использовать традиционный термин «электротон».
Распространение электротона - важный механизм клеточной сигнализации.
С помощью распространения электротона (главным образом катэлектротона) осуществляется функциональная связь между различными участками мембраны в клетках, не генерирующих ПД (глия, эпителий, так называемые тонические мышечные волокна[Б20]). Электротонические сигналы используются в телах нейронов и в дендритах таких мультиполярных нейронов позвоночных, как альфа-мотонейроны, клетки Пуркинье мозжечка, гигантские пирамидные клетки коры и крупные нейроны ретикулярной формации ствола мозга.
Если источник тока действует достаточно долго, то вдоль волокна (в обоих направлениях от электрода) устанавливается определенный градиент электротона (около анода — градиент гиперполяризации, около катода — деполяризации).
Величина электротона (U) экспоненциально убывает с расстоянием
В толстых волокнах – дольше. Дескрементом
(x) при удалении от источника:
U = Uo ∙ e-х/λ,
где U0 — электротон в точке приложения электрода; λ — постоянная длины, на которой электротон снижается в е раз (т.е. до 37 %).
где Rм — удельное сопротивление мембраны; RI— удельное сопротивление аксоплазмы; d ‑ диаметра волокна.
Ионотропные рецепторы (ПСП)
Ионотропные рецепторы – это ионные каналы, которые открываются в результате воздействия на них медиаторов. Медиатор присоединяется к специфическому месту на внешней стороне рецептора, что вызывает изменение конфигурации рецептора на несколько миллисекунд, в течение которого через ионный канал могут течь ионы.
Ионотропные рецепторы различаются:
По виду медиаторов, на которые они реагируют:
Например, ацетилхолиновые рецепторы взаимодействуют только с медиатором ацетилхолином).
По проницаемости для разных видов ионов:
Например, есть рецепторы, которые проницаемы только для ионов Na+).
Если ионотропные рецепторы проницаемы для ионов Na+, то в ответ на их открытие на постсинаптической мембране развивается деполяризация – возбуждающий постсинаптический потенциал (ВПСП).
Если ионотропные рецепторы проницаемы для ионов Cl-, то в ответ на их открытие на постсинаптической мембране развивается гиперполяризация – тормозящий постсинаптический потенциал (ТПСП). Разные медиаторы вызывают различные эффекты. Например, глутамат и ацетилхолин вызывают ВПСП, а ГАМК и глицин вызывают ТПСП.
41. Метаботропные синапсы (структура G-белка)
Метаботропные рецепторы
Примеры:
· Мускариновые ацетилхолиновые рецепторы (постганглионарные волокна симпатической нервной системы)
· Опиоидые
· Дофаминовые
· Норадреналиновые (постганглионарные волокна симпатической нервной системы
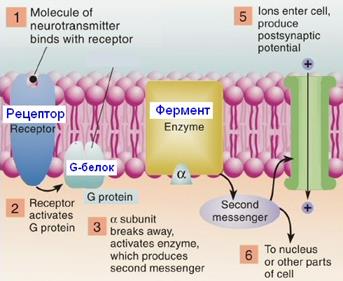
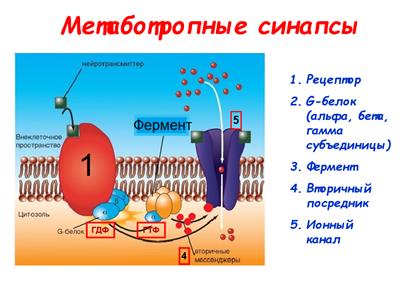
Рис. 809251152. Структура G-белка. Альфа-субъединица изображена с полостью, символизирующей сайт связывания ГДФ или ГТФ
ГТФ-азный цикла G‑ белков
В покое на постсинаптической мембране находится комплекс рецептор‑G-белок c ГДФ (рис. 809251335, 809251304).
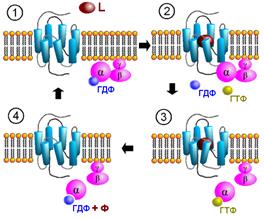 Рис. 809251335. Цикл активации G-белка под действием G-белок-связанного рецептора. 1 - фаза покоя; 2 - лиганд присоединяется и активирует рецептор; G-белок сталкивается с активированным рецептором, диссоциирует, альфа-субъединица теряет ГДФ; 3 - альфа-субъединица активируется, присоединяя ГТФ; альфа-субъедница присоединяется к эффектору и активирует его. Эффектор, в свою очередь, катализирует синтез вторичного мессенджера; 4 - дефосфорилирование ГТФ деактивирует альфа‑субъединицу, которая отсоединяется от эффектора, готовая начать весь цикл сначала. L – медиатор (лиганд, агонист). Рис. 809251335. Цикл активации G-белка под действием G-белок-связанного рецептора. 1 - фаза покоя; 2 - лиганд присоединяется и активирует рецептор; G-белок сталкивается с активированным рецептором, диссоциирует, альфа-субъединица теряет ГДФ; 3 - альфа-субъединица активируется, присоединяя ГТФ; альфа-субъедница присоединяется к эффектору и активирует его. Эффектор, в свою очередь, катализирует синтез вторичного мессенджера; 4 - дефосфорилирование ГТФ деактивирует альфа‑субъединицу, которая отсоединяется от эффектора, готовая начать весь цикл сначала. L – медиатор (лиганд, агонист).
|
Использованные в рис. 809251335 обозначения фаз ГТФ-азного цикла G‑белков не являются обязательными. В литературе выделяют 5, 6 и более фаз. На рис. 809251304 использовано иное обозначения. Однако принципиальной разницы в представленных схемах нет. Рисунок 809251304 Вам при желании легче будет воспроизвести.
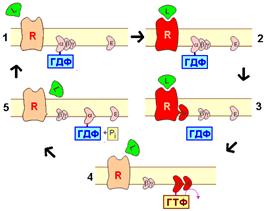
Рис. 809251304. ГТФ-азный цикла G‑белков. 1- фаза покоя; 2 - оединяется медиатора (лиганда) к рецептору; 3 - диссоциация G‑белка; 4 - активация эффектора; 5 - дефосфорилирование ГТФ. Красное окрашивание рецептора на рисунке символизирует активацию; R – рецептор; Е - эффектор; L – медиатор (лиганд, агонист).
Связывание медиатора (агониста) с рецептором приводит к белок-белковому взаимодействию между рецептором и G-белком и ускоряет диссоциацию ГДФ. В результате образуется короткоживущий комплекс агонист - рецептор - G-белок, не связанный ни с каким нуклеотидом.
Связывание с этим комплексом молекулы ГТФ снижает сродство рецептора к G‑белку, что приводит к диссоциации комплекса и высвобождению рецептора.
Потенциально рецептор может активировать большое количество молекул G-белка, обеспечивая, таким образом, высокий коэффициент усиления внеклеточного сигнала на данном этапе.
Активированная альфа-субъединица G‑белка диссоциирует от бета-гамма-субъединиц и вступает во взаимодействие с соответствующим эффектором, оказывая на него активирующее или ингибирующее воздействие.
Альфа-субъдиница с присоединенным с ней ГТФ способна взаимодействовать с эффектором в мембране - ферментами, такими, как аденилатциклаза, или, возможно, ионными каналами. Фермент может активироваться или ингибироваться, а ионный канал - открываться или закрываться. Конкретные примеры будут рассмотрены ниже и в последующих лекциях.
Взаимодействие с эффектором, однако, длится до тех пор, пока альфа-субъединица, являющаяся ГТФ-азой, удерживает ГТФ.
Вскоре присоединенный ГТФ гидролизуется до ГДФ. Когда это происходит, альфа‑субъединица снова меняет свою конформацию и теряет способность активировать эффектор. После этого альфа-ГДФ взаимодействует с бета-гамма-комплексом и снова образует тримерный комплекс, завершая, таким образом, цикл.
Предполагают, что комплекс из бета-гамма-субъединиц тоже может (прямо или опосредованно) влиять на эффекторные ферменты и ионные каналы (рис. 809251405).
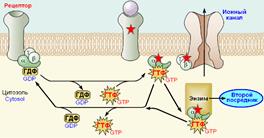 Рис. 809251405. Влияние бета-гамма-субъединиц G‑белка на ионные каналы.
Рис. 809251405. Влияние бета-гамма-субъединиц G‑белка на ионные каналы.
|
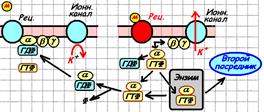 Рис. 809251405. Влияние бета-гамма-субъединиц G‑белка на ионные каналы (вариант для воспроизведения).
Рис. 809251405. Влияние бета-гамма-субъединиц G‑белка на ионные каналы (вариант для воспроизведения).
|
Пример прямой активация калиевого канала субъединицами G-белка показан на рис. 809251442.
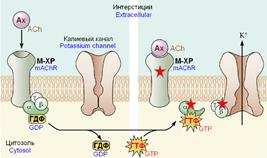 Рис. 809251442. Прямая активация калиевого канала субъединицами G-белка.
Рис. 809251442. Прямая активация калиевого канала субъединицами G-белка.
|
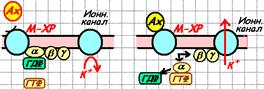 Рис. 809251442. Прямая активация калиевого канала субъединицами G-белка (вариант для воспроизведения).
Рис. 809251442. Прямая активация калиевого канала субъединицами G-белка (вариант для воспроизведения).
|
Пример прямого ингибирования Са2+-канала субъединицами G-белка показан на рис. 809251458.
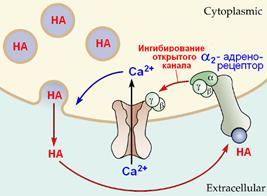 Рис. 809251458. Прямое ингибирование Са2+‑канала субъединицами G-белка.
Рис. 809251458. Прямое ингибирование Са2+‑канала субъединицами G-белка.
|
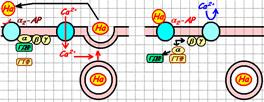 Рис. 809251512. Прямое ингибирование Са2+‑канала субъединицами G-белка (вариант для воспроизведения).
Рис. 809251512. Прямое ингибирование Са2+‑канала субъединицами G-белка (вариант для воспроизведения).
|
Активация пресинаптических адренорецепторов (ауторецепторов) приводит к диссоциации G-белка и последующему ингибированию (закрытию) кальциевых каналов, т.е. снижению уровня освобождение медиатора (норадреналина)
Пример метаботропной модуляции ионных каналов показан на рис. 709200023.
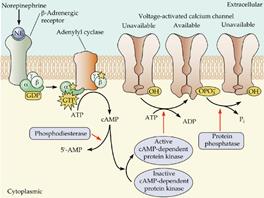 Рис. 709200023. Модуляция потенциал-зависимых кальциевых каналов норадреналином.
Рис. 709200023. Модуляция потенциал-зависимых кальциевых каналов норадреналином.
|
Пример регуляции экспрессии белков путем активации метаботропных рецепторов показан на рис. 809251517.
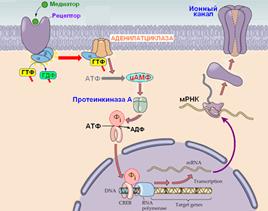 Рис. 809251517. Регуляция экспрессии белков путем активации метаботропных рецепторов.
Рис. 809251517. Регуляция экспрессии белков путем активации метаботропных рецепторов.
|
42. Нервно-мышечный синапс скелетного мышечного волокна (этапы передачи сигнала)
Нервно-мышечный синапс (neuromuscular junction, myoneural junction) - соединение концевой ветви аксона мотонейрона спинного мозга с мышечной клеткой.
Но концевой пластинкой (End-Plate) чаще называют постсинаптическую мембрану, отсюда возбуждающий постсинаптический потенциал (ВПСП) чаще называют потенциалом концевой пластинки (ПКП).
НМС – типичный химический синапс.
Холинергический, потому что медиатором является ацетилхолин.
Н-холинергический [эн, не аш и не эйч], потому что агонистом ацетилхолина в этом синапсе является никотин. Часто такие синапсы называют никотиновыми.
Н-холинергический, мышечного типа, потому что антагонистами ацетилхолина в этом синапсе являются кураре и курареподобные вещества.
Нервный отросток проходя по сарколемме мышечного волокна утрачивает миелиновую оболочку и образует сложный аппарат с цитолеммой мышечного волокна
|
Этапы передачи возбуждения в НМС
1. Деполяризация пресинаптической мембраны пришедшим по аксону потенциалом действия (ПД).
2. Открытие потенциалзависимых Сa++‑каналов на пресинаптической мембране и поступление Сa++ в пресинапс (пассивный транспорт).
3. Выход в синаптическую щель ацетилхолина путем экзоцитоза.
4. Диффузия медиатора к постсинаптической мембране.
5. Взаимодействие ацетилхолина с Н‑холинорецепторами постсинаптической мембраны мышечного волокна.
6. Открытие никотиновых рецепторных каналов постсинаптических каналов, пассивный вход Na+ в мышечное волокно.
7. Образование потенциала концевой пластинки - ПКП (возбуждающего постсинаптического потенциала - ВПСП) в области постсинаптической мембраны.
8. Электротоническое распространение ПКП в околосинаптическую область.
9. Формирование ПД мышечного волокна на сарколемме околосинаптической области.
|
|
Ацетилхолин в синаптической щели быстро разрушается ацетилхолинэстеразой (АХЭ), превращаясь в холин (он вновь захватывается для последующего синтеза) и ацетат.
|
Рис.. Схема чисто электрического синапса между дендритами мотонейронов лягушки (A), смешанного синапса между центральными окончаниями первичного афферентного нейрона и мотонейроном лягушки (B) и химического синапса между центральными окончаниями первичного афферентного нейрона и мотонейрона кошки (C).
Рис. 210211130. Схема саркомера.
Актиновые нити крепятся на Z‑мембране, миозиновые – на М‑мембране.
Участок между миофибриллы между Z‑мембранами называется саркомером
Саркомер принято считать структурно-функциональной единицей (СФЕ) миофибриллы скелетных и сердечных миоцитов.
Для иллюстраций мышечного сокращения удобно пользоваться упрощённой схемой саркомера.
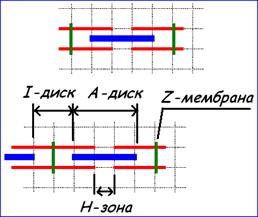
Рис. 709232249. Экстрафузальные (1) и интрафузальные (2) мышечные волокна Мышечное веретено (3).
Экстрафузальные мышечные волокна (рис. 709232249) образуют основную массу мышцы и выполняют всю работу, необходимую для движения и поддержания позы.
Двигательная единица
Мышечные волокна не являются функциональной единицей скелетной мускулатуры. Эту роль выполняет нейромоторная, или двигательная единица которая включает мотонейрон и группу мышечных волокон, иннервируемых разветвлениями аксона этого мотонейрона, расположенного в ЦНС
Число мышечных волокон, входящих в состав двигательной единицы, различно и зависит от функции, которую выполняет мышца в целом.
В мышцах, обеспечивающих наиболее точные и быстрые движения, двигательная единица состоит из нескольких мышечных волокон, в то время как в мышцах, участвующих в поддержании позы, двигательные единицы включают несколько сотен и даже тысяч мышечных волокон.
Все мышечные волокна каждой двигательные единицы (ДЕ) относятся к одному типу.
Композиция скелетных мышц
Каждая мышца уникальна по спектру входящих в её состав типов мышечных волокон. В зависимости от преобладания в мышцах конкретного типа мышечных волокон скелетные мышцы относят к «красным» и «белым» либо «быстрым» и «медленным».
Этот спектр генетически детерминирован.
Механизм мышечного сокращения и расслабления
Механизм мышечного сокращения объясняется моделью скользящих нитей, авторами которой принято считать Х.Хаксли и Дж.Хансона(1954 г.)
При микроскопии миофибрилл в расслабленном состоянии и состоянии сокращения было отмечено, что при сокращении длина А-диска не меняется, а I-диск и H‑полоска уменьшаются и даже исчезают. Отсюда был сделан вывод, что тонкие и толстые нити при сокращении не изменяют значимо свою длину, а скользят относительно друг друга
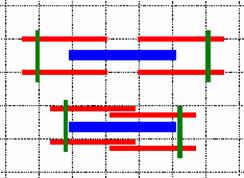
Рис. 709270838. Изменение саркомера при сокращении (упрощённая схема). Объяснение в тексте.
Рис. 709240356. Схема электромеханического сопряжения.
1. Образование потенциала концевой пластинки (ПКП).
2. Электротоническое распространение ПКП в околосинаптическую область.
3. Генерация потенциала действия мышечного волокна в околосинаптической области.
4. Распространение потенциала действия по поверхностной мембране мышечного волокна
5. Распространение потенциала действия по мембранам поперечных трубочек (Т‑системы).
6. Реакция рецепторов дигидропиридина [8] на изменение мембранного потенциала.
7. Передача сигнала дигидропиридиновыми рецепторами рианодиновым рецепторам терминальных цистерн саркоплазматического ретикулума.
8. Открытие каналов Ca2+, с рианодиновыми рецепторами и выход Ca2+ из саркоплазматического ретикулюма (цистерн) в саркоплазму (цитозоль).
9. Диффузия Ca2+ в межфибриллярное пространство.
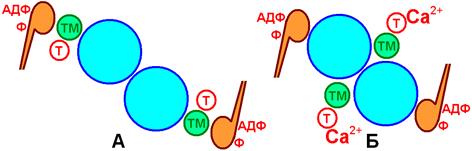
10. Связывание Ca2+ с тропонином, расположенным на тонком, актиновом, миофиламенте.
11. Конформационные изменения тропонина при связывании Са2+ ® «заталкивание» тропомиозина в канавки актиновых нитей
12. Открытие активного центра актиновых нитей.
13. Соединение головки миозина, каталитический центр которого связан с АДФ и фосфатом (продуктами гидролиза АТФ)
14. Начинается цикл миозиновых мостиков.
Типы мышечного сокращения
· Изометрическое- если оба конца мышцы неподвижно закреплены и при неизменной длине увеличивается напряжение
· Изотоническое – если один конец мышцы свободен. То в процессе сокращения длина мышцы уменьшится и при укорочении напряжение не изменится
· Ауксотонический(смешанный)- изменение длины и тонуса мышц, при сокращении которой происходит перемещение груза. Есть преодолевающий (концентрический) и уступающий (эксцентрический)
Режимы мышечного сокращения
Одиночные мышечные сокращения – возникают при низкой частоте электрических импульсов. Если очередной импульс приходит в мышцу после завершения фазы расслабления, возникает серия последовательных одиночных сокращений
При более высокой частоте импульсов очередной импульс может совпасть с фазой расслабления предыдущего цикла сокращения. Амплитуда сокращений будет суммироваться и возникает зубчатый тетанус – длительное сокращение, прерываемое периодами неполного расслабления мышцы.
При дальнейшем увеличении частоты импульсов каждый следующий импульс будет действовать на мышцу во время фазы укорочения, в результате чего возникает гладкий тетанус – длительное сокращение, не прерываемое периодами расслабления
Сила и работа мышц
Сила – способность за счет мышечных сокращений преодолевать внешнее сопротивление. При ее оценке различают абсолютную и относительную мышечную силу
Работа – энергия, затрачиваемая на перемещение тела с определенной силой(F) на определенное расстояние(S) формула – A=F*S
Если сокращение мышцы совершается без нагрузки, то механическая работа равна 0 (А=0)
Если при максимальной нагрузке не происходит укорочения мышцы, то работа также равна 0 (А=0). В этом случае химическая энергия полностью переходит в тепловую

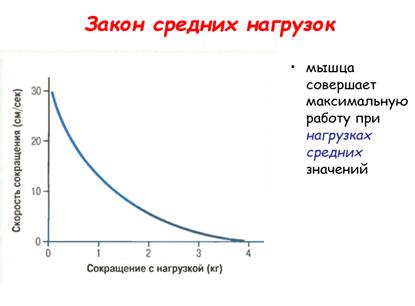
Закон средних нагрузок
Вопросы к итогу №1
Понятие «биомембрана»
МЕМБРА́НА, -ы, ж. от лат. membrana — кожица, перепонка.
Мембрана - упругая перепонка, тонкая пленка или пластинка, способная совершать колебания. Гибкая тонкая пленка, находящаяся в состоянии натяжения и поэтому обладающая упругостью.
Принцип построения всех биологических мембран одинаков, независимо от того, какой клетки (растительной или животной) или клеточной органелле она принадлежит.
Под мембраной чаще подразумевают перегородку, разграничивающую пространства, компартменты. Считается, что плазматическая мембрана вместе с ядром и цитоплазмой является одним из основных компонентов клетки.
Ядро и органеллы клетки также отделены от цитоплазмы мембранами, которые предупреждают свободное движение воды и растворенных в ней веществ из цитоплазмы в них и наоборот. Это создает условия для разделения биохимических процессов, протекающих в различных компартментах внутри клетки.
Гук действительно был первым человеком, увидевшим клетки. В 1663 году, пытаясь понять, почему пробковое дерево так хорошо плавает, Гук стал рассматривать тонкие срезы пробки с помощью усовершенствованного им микроскопа. Он обнаружил, что пробка разделена на множество крошечных ячеек, напомнивших ему монастырские кельи, и он назвал эти ячейки клетками. Р.Гук видел оболочку, стенку, образованную целлюлозой. Но не биомембрану.
Эволюция представлений о биомембране
Гук видел не мембраны, а оболочки (стенки) растительных клеток. Это надмембранные структуры.
Мембрану и сейчас плохо видят. Даже при внимательном рассмотрении фотографий ультратонкого среза живой ткани, выполненных с помощью лучших электронных микроскопов, мы можем увидеть только тонкие двойные линии, которые "вырисовывают" контуры клетки и внутриклеточных органелл.
 Рассмотреть тонкую структуру биомембраны даже на электронных микрофотографиях невозможно.
Рассмотреть тонкую структуру биомембраны даже на электронных микрофотографиях невозможно.
О том, что содержимое клетки отделено от окружающей её среды мембраной догадались.
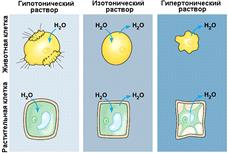
1855 год. К. фон Негели обнаружил, что неповрежденные клетки изменяют свой объем при изменении осмотического давления окружающей среды.
У растительных клеток и бактерий при их погружении в гипертонический раствор наблюдается плазмолиз - отставание плазматической мембраны от клеточной оболочки при уменьшении объёма протопласта. Фон Моль и К. фон Негели ввели термин «мембрана» для объяснения плазмолиза.
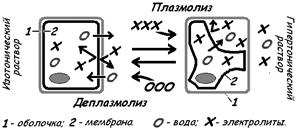 Эти опыты показали наличие на границе клетки полупроницаемой мембраны. Проницаемой для воды и плохо проницаемой для ионов натрия, хлораа
Эти опыты показали наличие на границе клетки полупроницаемой мембраны. Проницаемой для воды и плохо проницаемой для ионов натрия, хлораа
1895-1902 годы. Э. Овертон на основании того, что в его опытах неполярные молекулы легче проходили через клеточную мембрану, чем полярные соединения, делает вывод о липидной природе биомембран.
1925 год. Е.Гортер и Ф.Грендел сравнили площадь мономолекулярной пленки на поверхности воды, образованной липидами, экстрагированными из мембраны эритроцитов и поверхности этих эритроцитов. Получили соотношение значений площадей 2:1. Отсюда был сделан вывод, что мембрана эритроцитов состоит из липидных молекул, расположенных в два слоя.
1930-40 годы. В связи с необходимостью объяснить явное расхождение между поверхностным натяжением на границах раздела масло/вода и мембрана/вода Дж. Даниелли предполагает, что с мембранами связаны белки.
Внимание! «Бутербродная» модель Даниелли-Давсона устарела!
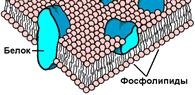
Была создана «бутербродная модель» Даниелли и Давсона мембрана состоит из двойного липидного слоя, а белок располагается на её поверхности.
Вопрос о взаимном расположении липидов и белков в мембране стал предметом многочисленных дискуссий, так как обнаружилось, что мембраны выполняют разнообразные функции.
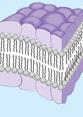
1972 год. С. Сингер и Дж. Никольсон предложили жидкостно‑мозаичная модель биомембраны.
К этому времени электронно-микроскопические показали, что в мембраны встроены глобулярные частицы. Данные спектральных исследований указывали, что для мембранных белков характерно высокое содержание α-спиралей и что они, вероятно, образуют глобулы, а не распределены в виде монослоя на поверхности липидного бислоя. Неполярные свойства мембранных белков наводили на мысль о наличии гидрофобных контактов между белками и внутренней неполярной областью липидного бислоя.
С. Сингер и Дж. Никольсон свели воедино все эти идеи, создав жидкостно-мозаичную модель.
В последние годы жидкостно-мозаичная модель также подверглась модификации. Выявляются новые функции цитоскелета. Не все мембранные белки свободно диффундируют в липидном бислое. Имеются данные о существовании в мембране липидных доменов. Обнаружены динамические ассоциаты липидов, обладающие более плотной упаковкой (рафты). Выявлен специфический класс амфифильных белков, которые под влиянием внеклеточных сигналов меняют свою гидрофобность и обратимо диссоциируют от мембраны.
В рамках этой модели мембрана представляется как фосфолипидный бислой, в который погружены свободно диффундирующие белки.
Тем не менее, жидкостно‑мозаичная модель в её разных модификациях все еще служит в качестве концептуальной основы для объяснения многих мембранных феноменов.