На этой основе у млекопитающих животных развились разнообразные половые циклы, которые отчетливо объединяются в две группу: а) астральные циклы, б) менструальные циклы.
а) Эстральные циклы (Asdelle, 1946; Marschall, 1910, 1936. Киршенблат, 1955; и др.) характерны для животных до-приматов. Внешним признаком циклов этой группы служит течка.
Эстральные циклы у женских особей различных видов млекопитающих могут иметь неодинаковую продолжительность течки и периодов покоя, могут отличаться по структуре цикла: известны циклы «‘простые» и «двойные». Овуляция в циклах животных может быть спонтанной и может наступать только после контакта с самцом. Нервно-физиологическую стимуляцию, необходимую для овуляции, можно в условиях эксперимента заменить другими раздражителями, например электромеханическим. Такие циклы выделяются в группу «циклов с зависимой овуляцией» (зависимые циклы). Несмотря на такие различия, во всех эстральных циклах сохраняется в основных чертах структура, близкая к изображенной на схеме (рис. 1).
В случае астральных циклов фолликулярная фаза овариального цикла соответствует стадии проэструса, овуляция - стадии течки или эструсу, лютеиновая фаза - стадии метаэструса и, наконец, стадии диэструса соответствует состояние относительного покоя в яичниках, когда возникающие волны роста примордиальных фолликулов угасают без того, чтобы часть их перешла к развитию и достигла зрелости. Уровень секреции половых гормонов в фазе покоя не превышает подпороговых величин.
Приведем несколько примеров различных астральных циклов.
Половой цикл опоссума (и других сумчатых животных). Относится к «простому» типу, так как развивающиеся во время фолликулярной фазы пролиферативные процессы переходят без какого-либо перерыва в секреторные. Овуляция наступает спонтанно и не сопровождается никакими деструктивными изменениями эндометрия. Течка имеет место и выражается в брачном поведении самки, в ее готовности к встрече с самцом. Если произошло оплодотворение, секреторный эндометрий превращается в эндометрий беременности. Если нет, то после овуляции наступает стадия ложной беременности, которая почти не отличается от истинной беременности ни продолжительностью, ни состоянием половых проводящих путей.
Овуляция у сумчатых (опоссум, сумчатые куницы и кенгуру) происходит во время течки, которая длится 5-7 дней. Короткая беременность сумчатых заканчивается рождением недоношенных детенышей, которые самостоятельно преодолевают расстояние до сумки, вползают в нее, присасываются к млечным железам. Здесь завершается их развитие до того времени, когда они станут способными к самостоятельному существованию.
Половой цикл крольчихи. Как и у сумчатых, у крольчих пролиферативный эндометрий превращается в секреторный и далее в эндометрии беременности без перерывов. Течка не сопровождается никакими видимыми выделениями или морфологическими перестройками проводящих путей. Она выражается в изменении поведения самки, в проявлении ее половых рефлексов.
10
Однако цикл крольчихи имеет особенности. Даже у виргинных самок матка может находиться в состоянии длительно сохраняющейся, как бы заторможенной, пролиферации, и в яичниках у крольчих при этом можно обнаружить созревающие и готовые к овуляции фолликулы. Такое состояние может длиться неопределенно долгое время, без того чтобы наступила овуляция. Для того чтобы овуляция произошла, необходим контакт с самцом. Если койтус будет стерильным, у крольчихи разовьется стадия ложной беременности. Именно у крольчих была впервые обнаружена и изучена стадия ложной беременности, которую затем нашли в половых циклах многих животных. Сходно протекают циклы у хорьков, кошек, норок, некоторых сусликов, землероек, хотя у некоторых из них есть внешние признаки течки (хорек, кошка, норка) - набухание «петли», выделения.
В циклах такого типа овуляция наступает не спонтанно, а после контакта с самцом. В таких случаях принято говорить о «зависимой» овуляции. Время наступления овуляции неодинаково: у кроликов, например, через 8-10 час после контакта с самцом, у кошек - через 24-30 часов, у норки - через 50 часов.
В «простых» циклах со спонтанной овуляцией последняя также наступает в различные по отношению к течке сроки: перед окончанием течки, через несколько часов после нее, а у некоторых, преимущественно у животных, впадающих в спячку на зимний период, течка может иметь место осенью, а овуляция происходит только весной, когда и оплодотворяются яйцеклетки (летучие мыши, куница и др.).
Половой цикл мыши и некоторых других мышевидных грызунов. В отличие от описанных выше типов циклов, у мышевидных грызунов половые циклы могут протекать в различных условиях по-разному.
1. Если самки содержатся в изоляции от самцов, начало фолликулярной фазы в яичниках совпадает, как обычно, с началом пролиферативных изменений в половых проводящих путях самок (стадия проэструс).
Когда яйцеклетки созревают, наступает стадия течки (эструс), во время которой происходит спонтанная овуляция, сопровождающаяся деструкцией эндометрия матки и обильным отторжением ороговевающих клеток эпителия вагины. Течка длится недолго (часы), затем её
11
проявления затухают, деструкция эндометрия матки и эпителия вагины сменяется атрофическими процессами (метаэструс), возвращающими слизистые оболочки проводящих путей к состоянию покоя (диэструс). После диэструса наступает новая пролиферация (проэструс).
На воле, где общение животных разного типа неограничено, фактически каждая течка самки, во время которой она подпускает самца, кончается беременностью. В этих случаях пролиферативные изменения проводящих половых путей сменяются у мышей секреторными, обеспечивающими прикрепление и выращивание оплодотворенной яйцеклетки. Новый цикл наступает после окончания лактации, хотя может начаться и раньше. В летние периоды грызуны нередко одновременно кормят одних детенышей и вынашивают следующих. В зоотехнике даже рекомендуется практиковать спаривание кормящих самок, например кроликов, для увеличения их продуктивности.
Описанное течение полового цикла также является примером «простого» цикла грызунов.
2. Иная картина наблюдается, если крысу или мышь подсадить к вазэктомированному самцу. Можно получить «двойной» половой цикл который будет существенно отличаться от «простого».
В циклах такого типа после контакта со стерильным самцом течка закончится, произойдет овуляция, разовьется стадия инволюции и наступит пауза - диэструс, как если бы цикл закончился так же, как «простой». Но следом за короткой паузой начнется новая пролиферация, более глубокая, продолжающаяся в секреторную и далее в стадию ложной беременности, которая и продлится некоторое, различное у разных видов грызунов время (около 2/з продолжительности нормальной беременности). Стадия ложной беременности завершится быстрой инволюцией. После паузы (диэструс) наступит новая пролиферация.
Во время «простого» цикла (без самца) на месте фолликула развивается малоактивное желтое тело, которое быстро угасает. В стадии ложной беременности желтое тело тоже малоактивно, оно слабо влияет на эндометрий матки, и секреторные изменения не достигают сколько-нибудь значительного развития. Но все же желтое тело ложной беременности грызунов, в отличие от желтого тела «простого» цикла, существует более продолжительное время и производит более глубокие перестройки эндометрия. Если же наступает беременность, желтое тело достигает полного расцвета, при этом секреторные изменения в матке протекают наиболее активно. Желтое тело беременности ответственно за формирование и течение беременности, во всяком случае до какого-то этапа в развитии плода.
Половой цикл собаки и самок горностая, соболя, лисы, африканской виверры и некоторых других животных. Пролиферативная стадия в эндометрии матки и вагины развивается синхронно фолликулярной фазе яичников. Течка (эструс) начинается в конце пролиферативной стадии и после спонтанной овуляции продолжается некоторое время в секреторной стадии. В циклах этого типа течка сопровождается деструкцией слизистой: при этом отторгается некоторая часть слизистой оболочки, разрываются мелкие кровеносные сосуды, кровь из них изливается в полость матки и выделяется наружу.
В яичниках фолликулы замещаются желтыми телами. Эндометрий матки после непродолжительной инволюции снова усиленно разрастается, в нем развиваются секреторные прегравидные изменения, и наступает стадия ложной беременности, которая продолжается несколько недель. Затем мукоза матки дегенерирует, разрушение ее сопровождается обильной секрецией и сильным слущиванием отмирающего эпителия. Все отторгающиеся образования выделяются в виде «белой менструации», достигающей значительного развития, например, у собак. Половые циклы собак, коров и сходных с ними животных всегда имеют
12
В своей структуре стадию ложной беременности, а в период течки на границе пролиферативной и секреторных стадий, у них отмечаются более или менее энергичные процессы деструкции слизистой оболочки половых проводящих путей, которые сопровождаются кровотечением (внутренним и внешним). Между эндометрием пролиферации и секреторным нет постепенного перехода. Конструктивные процессы пролиферативной стадии обрываются и начинаются снова после деструкции, когда закончится течка. Циклы, в структуре которых отмечается такой разрыв между конструктивными процессами в эндометрии, относятся к «двойным».
Следует отметить, что одомашнивание накладывает отпечаток на половую функцию животных, на ее периодичность и структуру. У домашних копытных переход от пролиферативной стадии к ложной беременности более сглажен, чем у свободноживущих животных.
б) Менструальные циклы. Циклы этого типа характерны для отряда приматов. Внешним проявлением их являются менструальные кровотечения.
Общая схема менструального цикла. Менструальный цикл женщины привлекал внимание многочисленных исследователей и изучен достаточно полно. Хуже изучены половые функции и половые циклы у других представителей отряда: полуобезьян, низших обезьян, человекообразных. Все же существующих данных достаточно для того, чтобы представить себе общую схему менструального цикла приматов.
Как и астральные циклы животных, менструальные циклы приматов начинаются ростом фолликулов в яичниках. Параллельно развивается пролиферативная стадия слизистой оболочки, выстилающей половые проводящие пути. К концу этой стадии матка гиперемируется, ее эндометрий утолщается, железы удлиняются, эпителий становится многослойным, в нем появляется много делящихся клеток. После спонтанной овуляции развиваются и функционируют желтые тела, а в матке продолжаются более интенсивные конструктивные процессы, начатые в первой фазе цикла. В результате слизистая матки заметно утолщается (в 8-10 раз). Ее капилляры расширяются, переполняются кровью, сквозь их стенки выпотевает жидкая часть крови, усиливающая отек стромы. Клетки стромы превращаются в округлые, богатые протоплазмой децидуальные клетки. Железы сильно разрастаются, углубляются, извиваясь, в толщу стромы и обильно секретируют. Исчезают реснички выстилающего полость матки эпителия. В отличие от циклов до-приматов, слизистая матки приматов в секреторной стадии дифференцируется на поверхностный, или функциональный, слой, в котором происходят все изменения, и базальный, остающийся почти неизменным на протяжении всего цикла.
В конце секреторной стадии через стенки сосудов мукозы выпотевают форменные элементы крови и в строме образуются мелкие лакунарные кровоизлияния.
В случае наступления беременности в подготовленную слизистую имплантируется яйцеклетка и происходит формирование характерной для приматов гемохориоидальной плаценты. Нередко этот процесс сопровождается маточным кровотечением, достаточно интенсивным для визуального наблюдения. Впервые об этом кровотечении сообщил Hartman (1932, 1939), который обнаружил его у самок макаков резусов на 14-37-й день после оплодотворения и дал ему наименование. Позднее знак плаценты был обнаружен у павианов, у шимпанзе и у женщин. Если беременность не наступила, функциональная слизистая матки быстро отторгается. Этот процесс сопровождается кровотечением. Работами Mаrкеé (1940), который наблюдал циклические изменения эндометрия на пересаженном в переднюю камеру глаза обезьяны кусочке слизистой матки, было показано, что в конце секреторной стадии, когда
13
начинает отслаиваться функциональный слой слизистой, кровеносные спиралевидные сосуды эндометрия матки (сосуды Дарона) резко скручиваются и так же резко распрямляются. При этом многие из них лопаются, выделяя кровь в полость матки. К этому кровотечению прибавляется кровь из очагов кровоизлияний стромы.
Но, как выяснилось позже, не у всех обезьян имеются спиральные артерии эндометрия, а у женщин кровотечение наступает после того как спиральные артерии выпрямятся. Поэтому была выдвинута новая теория менструации. В основу ее положено два факта, увеличение резистентности капилляров у женщин во второй половине цикла и полимеризация мукополисахаридной (гиалуроновой) кислоты в строме эндометрия (и в половой коже павианов) под действием эстрогенов в пролиферативной стадии цикла с последующим разрушением и деполяризацией после овуляции (Zachariae, 1959). Автор считает, что менструальное кровотечение - результат снижения резистентности капилляров, возникающего, когда из стенок сосудов уходят биофлавоноиды и стенки становятся дряблыми, проницаемыми; когда же под влиянием эстрогенов в стенках сосудов образуются эти соединения, резистентность сосудов восстанавливается.
Уже в конце менструации слизистая матки снова начинает покрываться функциональным эпителием, который достигает полного восстановления в пролиферативной стадии следующего цикла.
Несмотря на обилие гипотез, до настоящего времени нет полной ясности в представлении о механизмах регуляции менструаций.
Обобщая имеющиеся сведения, можно сказать, что менструация - самостоятельный процесс, не стоящий в преимущественной связи ни с овуляцией, ни с желтым телом. Различают два типа менструаций: в циклах без овуляции, когда кровотечение сопровождает отторжение пролиферативных образований; в циклах овуляторных, не перешедших в беременность, когда во время менструации отторгается сложное двойное разрастание слизистой.
Яичники находятся в теснейшей связи с менструацией, так как у овариектомированных обезьян и женщин кровотечение имеет место только один раз после операции, а затем прекращается и вызвать его можно только введением половых гормонов. Учитывая, что в циклах обезьян набухание половой кожи, как и пролиферативные изменения проводящих половых путей, развивается под действием половых гормонов, надо принять тезис П. В. Бочкарева «без пролиферации нет менструации», который был проверен нами более чем на 1.800 циклах самок павианов гамадрилов (Алексеева, 1948).
До недавнего времени менструальное кровотечение считали гомологичным течке животных. Изучение структуры половых циклов обезьян показало, что маточное кровотечение является концом цикла, тогда как течка связана с овуляцией, происходящей приблизительно в середине цикла. Пока не было известно о менструациях у обезьян, противники эволюционного учения ссылались на этот процесс как на особенность человека, противопоставляющую его всему животному миру.
Появление менструаций ставили в связь со сменой фаз луны, с особым «гормоном менструации», с «гипофизарным фактором кровотечения». С менструацией связывали появление в крови у женщин особых менотоксинов. Кстати, это представление - наиболее древнее. Оно породило обряды «очищения» женщин, бытовавшие у многих племен и народов.
Синхронно овариальному и эндометриальному циклам протекает и вагинальный цикл приматов. Но в вагинальном цикле у приматов менее ярко, чем у грызунов, выражено деление на стадии: в мазках и лаважах на всем протяжении цикла присутствуют одновременно все клеточные
14
элементы - лейкоциты, эритроциты, эпителиальные клетки базального, смешанного и ороговевшего типа.
Для установления по мазкам стадии полового цикла у самок приматов, в особенности у обезьян, нужно подсчитывать большое число клеточных элементов и устанавливать их количественные соотношения. Значительно более точны данные лаважей с последующим подсчетом всех клеточных элементов в них (Бочкарев, 1937; Арист, 1940).
От схематического описания менструального цикла приматов почти не отличается цикл женщин (см. рис. 1). Характерным для этого типа циклов является переход конструктивных процессов пролиферативной стадии без перерыва в секреторные. Овуляция не сопровождается деструкцией эндометрия. Эти черты позволяют отнести половые циклы приматов к циклам «простой» формы.
Половые циклы обезьян. Половые циклы обезьян протекают в соответствии с общим схематическим описанием менструального цикла приматов, но отличаются некоторыми особенностями (Алексеева, 1947).
Одной из основных особенностей являются рудиментарные следы течки на рубеже фолликулярной и лютеиновой фаз цикла. В это время отмечается частичная деструкция эндометрия,.сопровождаемая в день овуляции отторжением эпителиальных чешуек, а иногда и кровянистыми выделениями. Следы течки прослеживаются и в поведении обезьян. Д. Лавик-Гудолл (1967, 1970) для шимпанзе, Н. А. Тих (1950) для павианов гамадрилов, Loy (1970) для резусов отметили повышенное стремление самок к контакту с самцами именно в дни овуляции. Рудиментарная течка, как по морфофизиологическому, так и по психосексуальному компоненту, выражена у различных видов обезьян неодинаково интенсивно.
Открытие следов течки в половых циклах обезьян помогло понять некоторые остававшиеся долгое время непонятными особенности полового цикла женщин, а именно межменструальные кровотечения и межменструальные боли. Степень выраженности этих явлений подвержена значительным индивидуальным колебаниям. То, что эти симптомы появляются как раз на границе фолликулярной и лютеиновой фаз цикла, когда происходит овуляция, позволило причислить их к проявлениям течки, которая была присуща половым циклам животных предков человека, сохранилась в сглаженном виде в менструальном цикле обезьяньих предков человека и современных обезьян и, наконец, была обнаружена в рудиментарном виде в менструальном цикле женщин.
Вторым рудиментом в половом цикле обезьян оказалась стадия покоя, или пауза - гомолог диэструса животных. Она располагается между концом одного цикла (последним днем кровотечения) и началом последующего цикла (началом пролиферативной стадии).
Стадия покоя, или диэструс-пауза, сохранилась и в структуре женского менструального цикла (Алексеева, 1948).
Несмотря на значительные видовые и индивидуальные особенности, у половых циклов разных приматов имеется много общего. Различия ограничиваются продолжительностью циклов, степенью выраженности течки, менструации.
Кроме того, среди узконосых обезьян Старого Света многие виды обладают половой кожей - так названа совокупность оголенных участков кожи на теле обезьян, которые реагируют покраснением и набуханием на повышение уровня фолликулина в организме (естественное или искусственное). Основной участок обычно располагается в генито-анальной области (промежность и седалище). Однако у некоторых видов обезьян участки половой кожи могут находиться на хвосте (у его основания), бедрах, спине, груди, шее и даже на лице. Наиболее выразительные (изменения наблюдаются на участке половой кожи промежности. Как только в яичниках начинается фолликулярная фаза, половая
15
кожа краснеет, ее складки и морщины разглаживаются, наполняются и растягиваются, пока не образуется подушка интенсивного красного цвета. Размеры, форма и окраска набухания имеют видовые и индивидуальные различия. Перед овуляцией размеры набухания достигают максимальных, а поверхность его отличается блеском и сильным натяжением. Сразу после овуляции набухание становится тусклым, половая кожа покрывается сетью морщин, приобретает синюшный оттенок. В начале лютеиновой фазы набухание еще сохраняет свои размеры, а затем сморщивается, опадает и за несколько дней до конца фазы половая кожа возвращается к состоянию покоя. От остальной кожи половая кожа отличается почти полным отсутствием волос, даже рудиментарных. В редких случаях на ней можно обнаружить одиночные волоски. В половой коже не обнаружено скоплений пигмента. Ее красный цвет образуется за счет крови, наполняющей ее сосудистую сеть.
Природа и механизм изменений, происходящих в половой коже в связи со сменой фаз полового цикла, еще неясны. Набухание расценивали как локальный воспалительный процесс. Отсюда его названия «опухоль» или «менструальная опухоль».
Гистологическое строение половой кожи отличается богатством и сложностью кровеносных сосудов, легко наполняющихся кровью под действием различных раздражителей - температурных, механических. Покрывающие ее эпидермальные слои усиленно слущиваются, особенно когда набухание приближается к предельным размерам. Дермальный слой половой кожи разрыхляется, образуя подобие сети, переполненной межклеточной жидкостью и форменными элементами крови. Было подсчитано, что число ядер в 1 мм3 ткани половой кожи в состоянии покоя приближается к 200.000, а в дни максимального набухания их количество уменьшается до 60.000. Очевидно, набухание половой кожи осуществляется не за счет увеличения числа клеточных элементов. Klenerman (1955) показал, что клеточный состав половой кожи павианов гамадрилов изменяется в связи с ее физиологическим состоянием. С развитием набухания в половой коже увеличивается количество фибробластов и лимфоцитов, в состоянии покоя значительно увеличивается количество тучных клеток (mast cells).
Набухание половой кожи определяли и как регулируемый организмом физиологический отек. Но исследование химического состава жидкой части набухания половой кожи павианов показало, что протеины в ней составляют 3,4%. Жидкость из отечных очагов обычно содержит не более 1% протеинов. Это говорит о том, что жидкость набухания не является простым транссудатом. По содержанию протеинов она вдвое беднее плазмы крови и приближается к лимфе (Schall, 1955).
Половая кожа отличается пониженной чувствительностью к боли, почти полным отсутствием кровотечений при порезах и разрывах набухания. На разрезах ткань набухания становится белой, обильно выделяет бесцветную жидкость с небольшим количеством клеточных элементов. Повреждения набухания заживают очень быстро, и образующийся рубец впоследствии рассасывается бесследно. Если повреждение было достаточно большим, то следы его остаются в появлении добавочных долек, развитии асимметрии. Все эти свойства набухшей половой кожи объясняются особенностями ее строения и повышенной свертываемостью крови. Они имеют важное биологическое значение для обезьян. Набухшая подушка половой кожи, достигающая у павианов 1/6-1/5 величины тела обезьяны, легко травмируется. Не будь описанных свойств, повреждение набухания постоянно угрожало бы жизни самки.
Изучение половой кожи обезьян и ее изменений в связи с половыми циклами проводилось многими авторами. Наиболее подробное исследование было выполнено в Сухумском питомнике обезьян на десятках самок павианов, гелад, мартышек и макаков разных видов. Было уста-
16
новлено, что начало реакций половой кожи совпадает по времени с началом фолликулярной фазы цикла, максимум набухания - с днем овуляции. Нечто похожее на набухание половой кожи обезьян описано и у самок некоторых животных (хорька, лисы). В ходе развития пролиферативных изменений на половых органах самок формируется так называемая петля. В зоотехнике, особенно при разведении пушных зверей, состояние «петли» регистрируется и оценивается в баллах, что облегчает наблюдения за течением половых циклов, расчеты приблизительных сроков течки, установление ожидаемых сроков родов.
Набухание половой кожи у обезьян и «петля» некоторых животных - примеры конвергентного развития сходных по своему назначению особенностей.
Единственным безотносительным внешним проявлением в эстральных циклах является течка, которая не позволяет уточнить времени овуляции без помощи дополнительных сложных методов исследования (цитологических, оперативных и пр.). В менструальном цикле единственным точно регистрируемым проявлением оказывается менструальное кровотечение. Это привело к тому, что именно эти внешние проявления стали приниматься за исходные при изучении и описании половых циклов в зоотехнической практике и в клинике. Только в специальных морфологических работах речь идет о всех фазах половых циклов. Обычно при изучении тех или иных показателей жизнедеятельности, или синдромов, их оценивают только по отношению к явным проявлениям циклов — течке, менструации, что подчас затрудняет правильное понимание описываемых явлений.
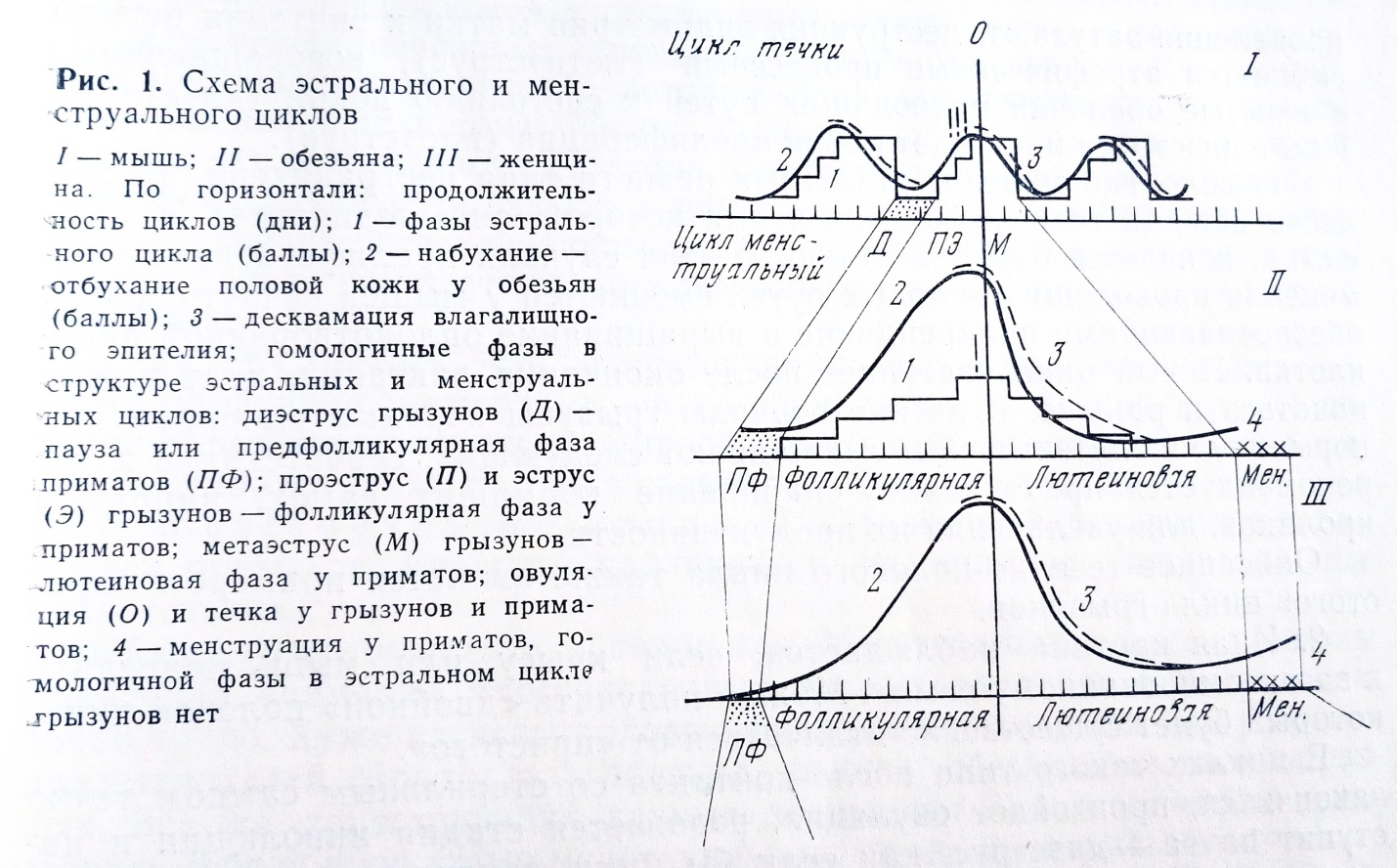
Благодаря наличию половой кожи половые циклы многих обезьян являются незаменимым объектом для изучения. В менструальных циклах обезьян легко и точно регистрируются все фазы цикла, становится доступным визуальное наблюдение за всей структурой цикла. Это позволило внести уточнения в представления о структуре не только циклов обезьян, но и циклов всех остальных типов животных и человека (рис. 1).
На рисунке отчетливо видно начало менструального цикла - день, когда начинается покраснение и набухание половой кожи, овуляция - день наиболее выраженного набухания, начало лютеиновой фазы - первый день инволюции половой кожи, конец лютеиновой фазы - последний день перед кровотечением. Менструальная стадия цикла продолжается от первого до последнего дня кровотечения. У обезьян в цикле обычно имеется предфолликулярная фаза, или пауза,- гомолог диэструса животных до-приматов. Пауза отмечается от конца менструации до начала следующего набухания.
Для проверки места каждого из внешних проявлений цикла в структуре цикла были проведены лапаратомии, гистологические и морфофизиологические исследования, эксперименты с воздействием фолликулина, прогестерона и других гормонов. Результаты исследований оказались бесспорными. Все, что в менструальном цикле многих приматов остается скрытым от глаз наблюдателя, у обезьян, имеющих половую кожу, становится доступным наблюдателю.
Так, в опытах с введением фолликулярных и фолликулостимулирующих гормонов была показана зависимость между уровнем гормона в организме самки и реакциями половой кожи. Совпадение дня овуляции с днем максимального набухания половой кожи было подтверждено на секционном материале и данными лапаратомий.
Наиболее яркое развитие набуханий половой кожи обнаружено среди приматов у шимпанзе, павианов разных видов, мандриллов и у некоторых видов макаков (яванские, лапундеры, тибетские). Значительно слабее набухание у гелад. У них основной участок половой кожи расположен на груди и шее. У макаков резусов половая кожа занимает большие
17
участки на седалище, бедрах, спине, лице. У многих видов мартышек (исключение - мартышка талапойн), у тонкотелых обезьян, равно и у обезьян Нового Света, половая кожа или отсутствует, или развита очень слабо (игрунковые обезьяны). Изучение таксономической ценности половой кожи показало, что этот признак имеет, скорее, вспомогательное значение для систематики приматов. Так, по развитию половой кожи можно отличить тибетских макаков от ассамских, у которых ее нет.
Нет половой кожи у полуобезьян (тарзиусов, руконожек, тупаии). Лишь у лемуров описано набухание наружных половых органов, позволяющее наблюдать смену фаз половых циклов (Petter, 1962, 1963), сходных с циклическими изменениями «петли» животных.
Сходство и различие астральных и менструальных циклов. Заканчивая описание половых циклов у самок приматов, следует отметить что по сравнению с половыми циклами течки менструальные циклы меньше различаются по структуре и среди них нельзя выделить различные типы циклов. Все они по структуре относятся к «простым» циклам со спонтанной овуляцией.
Сопоставление различных астральных циклов с менструальными циклами приматов позволяет отчетливо установить черты сходства и различия между ними (табл. 1).
Таблица 1.






