У женщин (Помаскина, 1960), как и у самок павианов, концентрация и масса протеинов в сыворотке крови были невысокими в первой половине цикла, увеличивались скачкообразно на границе фаз (день максимального набухания половой кожи, овуляции) и оставались высокими в лютеиновой фазе цикла, снижаясь к менструации.
В первой половине цикла преобладали альбумины, во второй - глобулины.
В плазме крови женщин концентрация и масса фибриногена изменялась таким же образом, как и общие протеины.
В плазме у самок павианов чакм определили высокую фибринолитическую активность, большое количество фибрина и усиление синтеза мукопротеидов в фолликулярной фазе, с максимумом к овуляции. В секре-
46
торной фазе эти показатели снижались до минимальных к менструации (Cohen, Berg, 1951; Gillman et al., 1960; Schulz, 1954).
Достоверное увеличение йода, связанного с белком (СБЙ), после овуляции, с максимумом в середине лютеиновой фазы и последующим снижением к регулам у женщин установили Садыков М. Ш. с соавторами (1968) и у самок павианов (van Zyl, 1957).
Электролиты и микроэлементы
Количество калия в крови у женщин повышено в фолликулярной фазе цикла, максимально к овуляции и в начале лютеиновой фазы, а затем снижается, давая еще один пик в дни менструации (Левинсон, 1967; и др.).
Отчетливо выражены и колебания натрия. Содержание его в крови достигало максимума к овуляции и было минимальным во время регул. В фолликулярной фазе уровень калия был достоверно более высоким, а натрия -— более низким, чем в лютеиновой (Thomas, 1953; Левинсон, 1967). Увеличение кальция крови у женщин в лютеиновой фазе и перед менструацией отчетливо показали Bell, Hick (1903) и др. В. Б. Левинсон (1967) обнаружил самый низкий уровень хлоридов у женщин в начале фолликулярной фазы, резкое увеличение к овуляции, высокий уровень в лютеиновой фазе и еще более высокое содержание в дни менструации (статистически достоверно).
В связи со сменой фаз менструального цикла у женщин описаны колебания в содержании железа, мышьяка, брома, меди, йода, магния, цинка и кобальта, которые, различаясь в деталях, все в общем характеризовались одной динамикой - до овуляции уровень их был более низким, чем во второй части цикла (Атауллаханов, 1964; Иоскович, 1970; и др.).
Обмен
Обмен веществ и энергии
В основе разнообразных форм проявления жизни лежат превращения энергии. Энергетические процессы живых существ складываются из газообмена, теплопродукции и теплоотдачи.
Для суждения об энергетическом обмене, для характеристики интенсивности обмена веществ оказалось достаточным определять минимальное количество энергии, необходимой для поддержания основных жизненных процессов, или основной обмен.
Под этим термином понимают величину расхода энергии в состоянии полного покоя при расслаблении мышц, при возможно более полном отсутствии внешних раздражителей, натощак и при оптимальных температуре и влажности воздуха.
Уровень основного обмена весьма непостоянен - он изменяется не только в течение суток и по сезонам, но зависит от климатических условий, величины атмосферного давления, газового состава воздуха, питания, физиологического состояния испытуемых, их возраста, пола, индивидуальных особенностей. Кроме того, в реакциях энергетического обмена на все эти условия ведущую роль играет кора головного мозга. Известно, что можно выработать условные обменные рефлексы. Исследовали основной обмен у кондукторов товарных вагонов, находящихся зимой на открытых площадках вагонов. При одной и той же температуре воздуха нашли различные энергетические траты в зависимости от того, ехали кондуктора из дому или возвращались домой (Понучаева, I960).
Несмотря на большую лабильность энергетических трат, половые различия при правильном методическом подходе выступают достаточно
47
отчетливо даже на таком неустойчивом фоне. Правда, литературные данные по вопросу о времени появления половых различий в основных энергетических тратах у человека и о направленности этих различий весьма пестры. Большое количество наблюдений свидетельствует о возникновении половых различий в основном обмене у детей с 3-6-месячного возраста и о том, что у девочек до периода созревания (10-12 лет) основной обмен оказывается более высоким в среднем, чем у мальчиков, как, впрочем, и вес тела. У взрослых женщин основной обмен на 7-10% ниже в среднем, чем у мужчин. К старости половые различия обмена сглаживаются.
По данным многих авторов величины основного обмена у женщин варьируют в связи с различными внешними воздействиями в значительно более широких пределах, чем у мужчин. Е. М. Беркович (1964) дает соответственные границы изменений: от 3,5 до 4% у мужчин и от 4,7 до 15% для женщин. На более широкие границы в изменениях основного обмена у женщин указывают также Goetza (1962) и др.
Что касается закономерных волнообразных изменений основного обмена у женщин по фазам половых циклов, то хотя данные по этому вопросу противоречивы, большинство авторов такие колебания обнаружили.
В пролиферативной стадии цикла основной обмен у женщин невысок и наименьшие величины его совпадают со временем овуляции. После чего начинается увеличение основного обмена, а следовательно, и энергетических трат. Они держатся на высоком уровне в секреторной стадии цикла и снижаются перед или во время менструации.
Ряд авторов обнаружили повышение основного обмена в предменструальном периоде и снижение после него. Наиболее детальная работа была выполнена Е. М. Говорухиной (1964), которая исследовала основной обмен (изменения газообмена по методу Рида с помощью аппарата Крога) у здоровых женщин и у женщин с аменореей: определения производились утром натощак после 2-х суток безбелковой диеты, на 8, 14, 20, 24 и 28-й дни цикла. У всех 10 женщин основной обмен снижался в первой половине цикла, повышался при переходе в секреторную стадию, был высоким в течение недели и снижался перед концом цикла. При аменорее яичникового типа никаких циклических изменений газообмена не прослеживалось, а при аменорее маточного типа и нормальном двухфазном цикле яичников отмечались циклические изменения основного обмена, сходные с установленным в циклах у здоровых женщин. Некоторые авторы подтвердили увеличение потребления 02 при дыхании у женщин перед менструацией, которое еще больше усиливалось, если испытуемые не лежали, а стояли или выполняли какую-нибудь работу (Schulz, 1954; Grollman, 1935).
Изучение газообмена у обезьян впервые производилось на базе Сухумского питомника А. Д. Слонимом и его сотрудниками. Камерным методом Реньо в модификации Шатерникова были установлены видовые, возрастные и половые различия основного обмена. Авторы показали, что величина теплопродукции у обезьян, как у людей, изменялась в течение суток, по сезонам, в зависимости от температуры среды, от пищи.
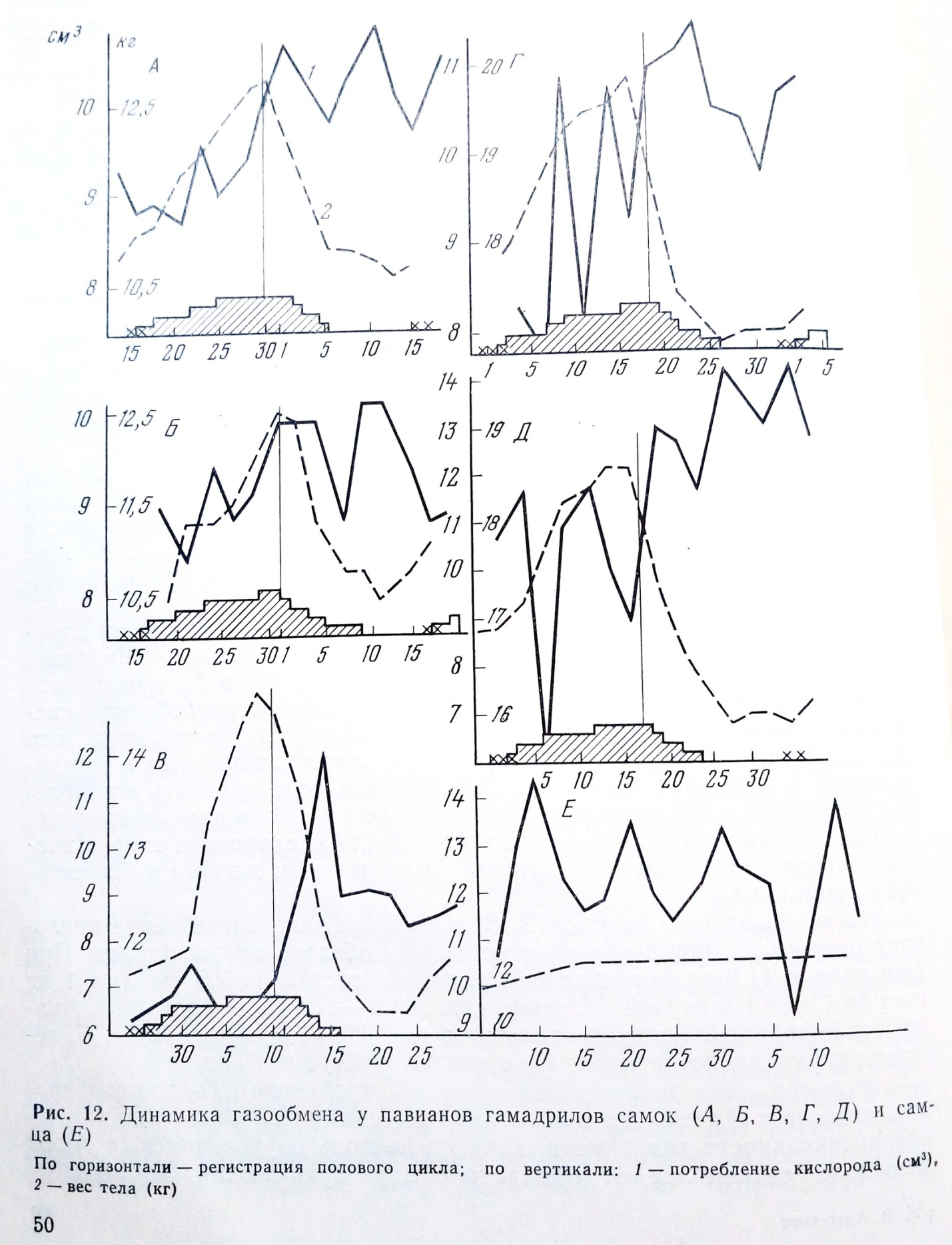
Исследование основного обмена у самок павианов гамадрилов в связи с половыми циклами проводилось (Алексеева, Авджиан 1961) на трех половозрелых здоровых самках. Взрослый самец этого же вида был взят в опыт в качестве контроля. Определение газообмена производилось утром натощак.
В течение первого месяца обследования обезьяны приучались к процедуре эксперимента - к перевозке в лабораторию, взвешиванию в специальной клетке, помещению в камеры и двухчасовому пребыванию в них. Результаты определения газообмена на протяжении двух последующих циклов были включены в обработку. Половые циклы регистрировались по принятой в питомнике схеме (Алексеева, 1974). Потребление О2 и
48
выделение СО2 в литрах за 2 часа определялось по Шатерникову (табл. 4).
У всех трех самок потребление О2 (см3) за 1 мин. на 1 кг веса в первых фазах цикла было ниже, чем после овуляции. В динамике это выглядело так: потребление О2 и выделение СО2 у обезьян было низким в начале пролиферативной стадии цикла и еще более снижалось к овуляции. В секреторной стадии цикла увеличивалось и потребление О2 и выделение СО2 (рис. 12). Различия оказались статистически достоверными.

Энергетические траты (в больших калориях за 1 час на 1 кг веса тела) в пролиферативной стадии у всех трех самок были много ниже, чем в секреторной. Так, у Маркитантки этот показатель в первой половине цикла колебался от 2,3 до 1,95 ккал, а во второй фазе цикла повышался до 2,7-3,0 ккал.
Потребление кислорода (см3/кг/мин) у контрольного самца Имама, который обследовался одновременно с самками, изменялось в несколько более узких пределах, чем у самок. Подъемы и снижения интенсивности потребления О2 у Имама носили неопределенный характер: в течение месяца наблюдений у Имама 4-5 раз увеличивалось, а затем снижалось потребление кислорода (см. рис. 12).
Ранее было установлено, что энергетический расход у обезьян изменяется пропорционально весу тела - чем крупнее обезьяна, тем ниже энергетические затраты на 1 кг веса тела. Можно было бы думать, что у самок обезьян, наблюдавшихся нами, изменения в потреблении кислорода и энергетические траты также определяются изменениями веса тела. Действительно, у всех трех самок вес тела увеличивался к середине цикла и снижался после овуляции. Однако, если сравнить потребление О2 у одной и той же особи при равном весе тела, то оказывается, что в пролиферативной стадии цикла потребление кислорода и энергетический расход будут менее интенсивны, чем при таком же весе в секреторной стадии цикла.
Это хорошо видно на рис. 12, В, где изображено потребление кислорода обезьяной Маркитанткой в первом из обследованных циклов. При весе тела в 11,9 кг потребление кислорода составило 7,5 см3 (на 1 кг веса за 1 мин.) в первой половине цикла и 9-11,9 см3 - во второй. Аналогичные сравнения можно сделать по рис. 12, Г, Д и рис. 12, А, Б, демонстрирующим потребление О2 в циклах у Кето и Мрии.
Непрямая связь между весом тела и потреблением О2 хорошо прослеживается у Имама, вес тела которого, несмотря на подъемы и снижения интенсивности газообмена, сначала несколько увеличился, а затем длительно сохранялся от опыта к опыту практически постоянным
49
(рис. 12, Е), в то же время потребление кислорода значительно колебалось. Из приведенных данных видно, что у самок обезьян наблюдаются определенные волнообразные изменения потребления О2 и выделения СО2 в связи со сменой фаз половых циклов и что эти изменения сходны с описанными для женщин.
Известно, что интенсивность основного обмена зависит от количества и качества потребляемой пищи и в свою очередь определяет потребность в пищевых продуктах.
Имеются отдельные сообщения об изменениях аппетита и вкуса у женщин в связи с менструацией. Увеличение аппетита совпадало с секреторной стадией цикла. Перед менструацией увеличивались аппетит и жажда. Женщины вообще тоньше различают сладкое, горькое и кислое,
особенно в предменструальный период. Во время менструации притупляется вкус сладкого. Правда, имеются наблюдения за изменением в эти дни десен, а именно за набуханием и разрыхлением их, что также может влиять на вкусовые восприятия. Krohn, Zuckerman (1937) показали, что макаки резусы больше поедают сухих кормов во второй, чем в первой половине циклов.
В тщательно проведенном эксперименте Gillman, Gilbert (1956) на павианах чакмах установили, что потребление корма (в калориях) уменьшалось с 6-го по 11-й дни цикла, оставалось низким до овуляции, затем увеличивалось и достигало максимальных величин за 2-7 дней до менструации. В первой фазе цикла потребление пищи за один день в больших калориях составляло 43-53 кал/кг/день, во второй фазе при равном весе тела - увеличивалось до 75 кал/кг/день, т. е. в полтора раза.
В этом опыте самки получали пищу два раза в день без ограничения. В составе рациона были корма животного и растительного происхождения, что обеспечивало свободу выбора по вкусу и по потребности. Было отмечено увеличение потребления белковых кормов в секреторной стадии цикла. Однако при этом обезьяны не уменьшали потребления углеводных и витаминных кормов. Попытка ограничить кормление самок обезьян только белковым питанием (маисом) закончилась гибелью 6 самок из 21 до перевода их на общий рацион и еще двух через некоторое время после перевода.
При этом у всех самок наблюдались расстройства половых циклов - аритмия, многовершинные набухания половой кожи, длительные паузы, аменоен (Gillman, Gilbert, 1956).
Оба исследования наглядно демонстрируют взаимосвязь, существующую между питанием, обменом и половыми циклами у обезьян. Специфическое динамическое действие углеводов и белков исследовалось в опытах на обезьянах (Слоним, Макарова, 1949). Авторы отметили значительные колебания основного обмена в ответ на кормление углеводной и белковой пищей, особенно у самки макака резуса, у которой изменения дыхательного коэффициента были выражены значительно сильнее, чем у самца, что оказалось связанным с протекавшим у нее во время обследования половым циклом.
Что касается изменений основного обмена у самок других видов животных в связи с циклами, то таких данных почти нет. Имеющиеся работы показывают половые различия обмена в нормальных условиях и при нагрузках. Так, Ю. М. Гефтер с соавторами (1948) констатировали факт существования половых различий в реакции на голодание: самцы быстрее теряли в весе, чем самки (на сроки в 70-77 час. и 100-110 час. соответственно), и у них определялись различия в количестве продуктов белкового и других видов обмена: ацетоновых тел у самцов в крови было меньше, кетоновых тел больше, сахара меньше, чем у самок, и т. п.
О зависимости газового обмена у самок белых мышей от функционального состояния половых желез сообщил в 1961 г. В. В. Полынин. Он определял газообмен в камере закрытого типа по методу Миропольского. У неполовозрелых мышей потребление кислорода в среднем не превышало 100,2 мл на 100 г веса. Введение 50 ед. масляного раствора синэстрола понижало газообмен на 13-14%. Наибольшее количество кислорода поглощалось самками во время течки. Кастрация снизила газообмен на 21%. Введение синэстрола кастратам не повысило потребления О2.
Наиболее универсальной и важной функцией человека и животных является мышечная деятельность. И. М. Сеченов в 1863 г. писал: «...окончательным фактом является мышечное движение. Все внешние проявления мозговой деятельности могут быть сведены на мышечное движение».
51
Известно три пути снабжения сокращающихся мышц энергией - анаэробное фосфорилирование, окислительное фосфорилирование и прямое окисление. Основным источником энергии в организме оказывается окисление углеводов, на втором месте стоят жиры и меньше всего на энергетические траты расходуется белков.
Установлено, что такой жизненно важный процесс, как окисление также волнообразно изменяется по фазам половых циклов. Okamoto (1960) приводит данные об изменении тканевого дыхания в связи с половыми циклами у крыс: в большинстве тканей (промежуточный мозг матка, вагина, щитовидная железа) потребление О2 увеличивается и достигает максимума в фазах предтечки и течки. В надпочечниках - то в стадии течки, только интенсивность окисления начинает увеличиваться несколько позже. Во всех обследованных тканях потребление кислорода и интенсивность тканевого дыхания уменьшается в стадии диэструса. Удивительным в работе Okamoto явился факт практически неменяющегося на всем протяжении цикла тканевого дыхания в яичниках.
У самок по фазам полового цикла и у самцов в параллельных опытах определяли окислительные процессы и гликолитическую активность передней и задней областей гипоталамуса. В переднем отделе гипоталамуса наибольшее значение О2 определялось у самок крыс в течке и наименьшее - в диэструсе. У самцов потребление кислорода тканями и гликолитическая активность переднего отдела гипоталамуса оказалось близким с величинами, характерными для самок в диэструсе и ниже, чем у самок, в течке (статистически достоверно).
В заднем отделе гипоталамуса потребление О2 и гликолитическая активность у самцов были сходны с таковыми у самок в фазе диэструса и выше, чем у самок в проэструсе и эструсе (статистически достоверно) (Mogilevsky et al1968). Энергозависимые реакции дыхания митохондрий в гомогенатах печени мышей в связи с фазами половых циклов определялись полярографически (Алексеева, Ахмеровой др., 1974). Метод позволяет различать скорости дыхания митохондрий в отрегулированном (покой, отдых) и активном (при добавке АДФ) состояниях, судить о времени фосфорилирования экзогенного АДФ и количестве требуемого на это кислорода, рассчитывать дыхательный коэффициент, отношение АДФ/О и соотношение скоростей дыхания состояний покоя (состояние 4) и активности (состояние 1). Оказалось, что энергозависимые реакции дыхания митохондрий изменяются по фазам полового цикла. В диэструсе дыхание отличалось высокой активацией на добавление АДФ и ДНФ и низкой скоростью дыхания в состоянии покоя. Присутствие в среде инкубации глютаминовой кислоты приводило к дополнительной активации дыхания активного состояния. В проэструсе и эструсе уровень дыхания покоя повышался, ослаблялась стимуляция дыхания глютаматом и укорачивалась способность дополнительного снижения скорости дыхания (состояния покоя) после последовательных циклов фосфорилирования (рис. 13, A).
В метаэструсе было получено два типа ответов. Один являлся как бы продолжением низкоэнергетического уровня проэструса и эструса. Продолжался рост скорости дыхания состояния покоя и затягивание времени рабочего цикла фосфорилирования со снижением его амплитуды (рис. 13, Б, а).
Ответ второго типа отличался высокой амплитудой и большой скоростью рабочего цикла, при этом добавление глютамата не стимулировало, а угнетало активное дыхание. Такое «синтезирующее» состояние митохондрий, по М. Н. Кондрашовой, обеспечивает активность процессов репарации с наибольшей полнотой (рис. 13, Б, б).
На электронно-микроскопических препаратах нативной суспензии митохондрий, выделенной из гомогенизированной печени самок мышей, забитых в разных фазах полового цикла, изучались различия среднего
52

объема (по диаметру) митохондрий трех различных типов строения и распределение морфологически неодинаковых митохондрий.
Были выделены следующие типы митохондрий: 1-й - округлые с тонким рельефом, 2-й - округлые с грубым рельефом и 3-й - полиморфные с грубым рельефом. Оказалось, что митохондрии 1-го типа численно преобладают в диэструсе, эструсе и позднем метаэструсе, 2-го типа - в раннем метаэструсе. В проэструсе все три типа митохондрий встречались в одинаковых долях. Средние диаметры (объемы) митохондрий 1-го и 2-го типов определялись одинаковыми в фазах диэструса, проэструса и позднем метаэструсе, достоверно были меньшими в эструсе и увеличенными в раннем метаэструсе (Алексеева, Стефанов, 1973; Алексеева и др., 1974).
Таким образом, было показано, что по фазам полового цикла изменяются строение и. функция митохондрий печени у самок мышей.
Углеводный обмен
Изменениям углеводного обмена в связи с циклами у самок посвящено большое количество работ. Но данные все еще противоречивы. Одни авторы не находят связи между углеводным обменом и фазами половых циклов Другие считают, что эндокринная деятельность яичника снижает содержание сахара в крови. Большинство же авторов обнаружили гликемическое действие фолликулина.
В литературе собрано достаточно наблюдении за нарушением овариально-менструальных циклов и репродуктивной функции при диабете и других расстройствах углеводного обмена. Л. И. Лобановская (1961) нашла расстройства половых циклов у 67,7% больных диабетом.
В 1926.г Eufinger и др. сообщили о повышении сахара в крови у голубей и уток во время овуляции Л. Г. Лейбсон и Р. С. Лейбсон (1940) изучали регуляцию сахара крови у собак с различными типами высшей нервной деятельности в зависимости от фаз полового цикла. Алиментарные гипергликемические кривые оказались различными у одних и тех же особей собак: в фазах диэструса, предтечки и особенно течки гипергликемические кривые намного превышали кривые, полученные в фазе послетечки.
Различия типов высшей нервной деятельности проявлялись в относительно более ярких кривых у возбудимых собак.
53
В. Я. Суетин и Ф. А. Чиркин (1955) установили факт повышения сахара крови (с 18-20 до 47-50 мг%) во время течки у самок белых мышей.
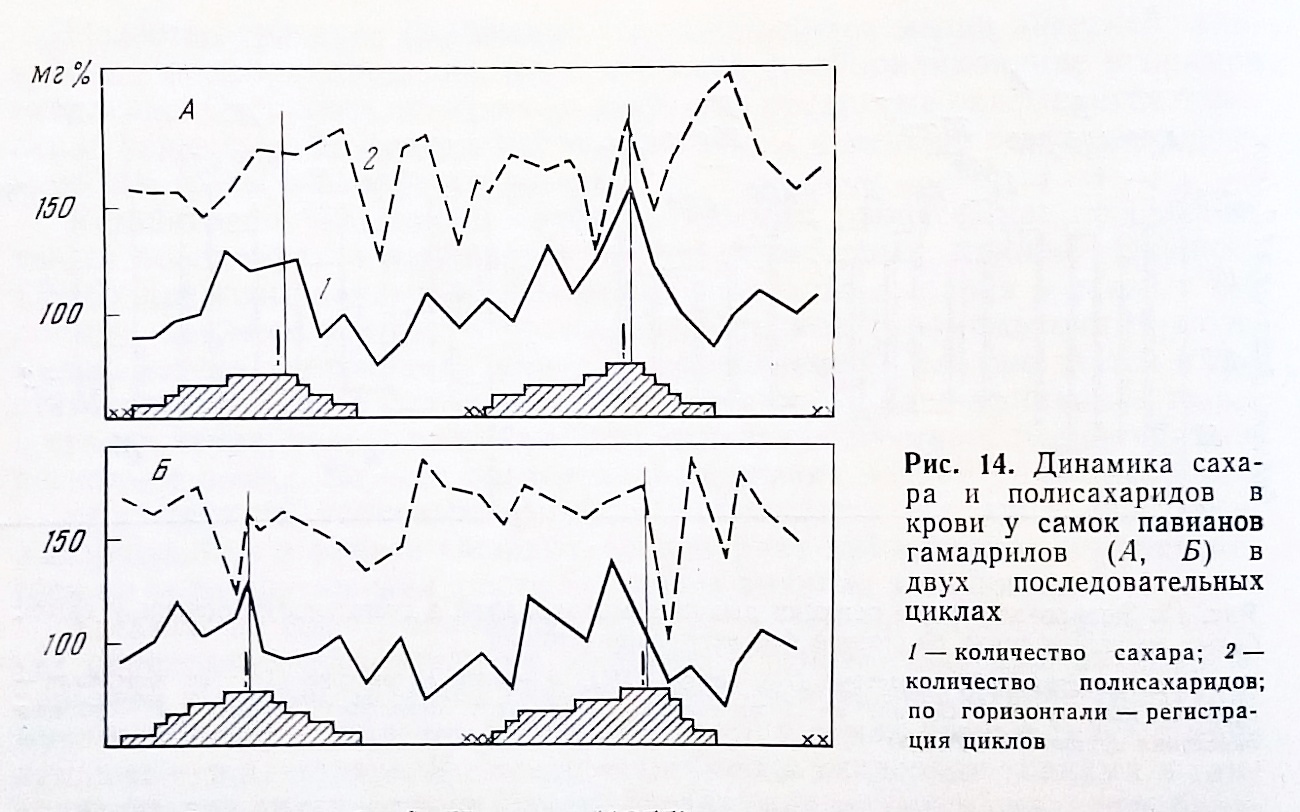
В 1960 г. Н. З. Майсурадзе исследовал углеводный обмен и половые циклы у самок крокодилов, морских свинок и белых крыс. У всех обследованных им самок уровень сахара крови изменялся в связи с фазами половых циклов. В покое он не превышал 70-80 мг%, в предтечке начинал подниматься, в фазе течки уровень сахара был самым высоким – 110-120 мг%. Различия в содержании сахара по фазам цикла оказались статистически достоверными. Менее определенные изменения уровня сахара и полисахаридов крови по фазам половых циклов были установлены нами у самок павианов гамадрилов (Алексеева, 1957).
В 18 из 30 половых циклов у 10 самок было обнаружено повышение уровня сахара и снижение полисахаридов крови ко времени овуляции. В секреторной фазе уровень сахара оставался высоким. Он снижался перед менструацией (рис. 14).
Неудачу следует объяснить высокой лабильностью этого показателя у обезьян. В первом цикле, пока обезьяны привыкали к процедуре обследования (подаче, взятию крови из уха), волнообразных изменений сахара по фазам цикла почти не прослеживалось ни у одной из самок. Оказалось, что уровень сахара в крови у обезьян быстро изменялся всякий раз, как что-нибудь изменялось в окружающей обезьяну среде во время обследования или даже перед ним. Каждое изменение очередности подачи, более или менее энергичной фиксации служителями обезьяны, даже такая «мелочь», как перемена головного платка у служительницы, которая фиксирует голову обезьяны, или нарисованная на щеке у знакомой служительницы родинка, - все вызывает ориентировочно-оборонительную реакцию обезьяны. С полным основанием можно включить сахар крови в число компонентов ориентировочного рефлекса обезьян, смысл которого в мобилизации организма к тому неизвестному, что может повлечь за собою замеченное обезьяной изменение.
Более определенные изменения сахара крови в цикле проследил Л. А. Фирсов (1963) у трех самок шимпанзе. Сахар крови нарастал в фазе пролиферации, достигал максимума ко дню овуляции, держался некоторое время на высоком уровне, а затем снижался перед менструацией. Л. А. Фирсов определял также гликемические кривые у шимпанзе после нагрузки 100-200 г сахаром. Полученные кривые значительно различались в зависимости от фазы цикла, когда ставилась функциональная
54

проба. В фолликулярной стадии цикла гликемические кривые имели наиболее выраженную форму: резкое увеличение сахара в крови через 15-60 мин. и снижение через 120 мин. В дни овуляции не только не было подъема уровня сахара, но уже после 15 мин. начиналось снижение содержания сахара, которое продолжалось через 60-90 и 120 мин. наблюдения. Во время менструации кривые были ломанными, уровень сахара крови то поднимался, то снова снижался, но к 120-й минуте наступала гипогликемия.
У женщин по фазам цикла четкую картину волнообразных изменений в содержании сахара крови установил Artner (1954). Он определял уровень сахара крови в циклах у 30 женщин и нашел, что в стадии пролиферации содержание сахара в крови в среднем было равно 89,06%, а в стадии секреции - 102,34% (статистически достоверно).
Переход от низкого уровня к высокому был резким и совпадал с днем овуляции. Сходные данные получили и другие авторы.
Изучение углеводного обмена проводилось на клеточном уровне.
Ю-Ай и Чжан Цзяо-чен (1958) гистохимическим методом исследовали содержание гликогена в клетках яйцепроводов, вагины у белых мышей и нашли, что его количество увеличивалось в фазах течки и в начале послетечки. Они же сообщили, что под действием эстрогенов (имитация пролиферативной фазы) повышается содержание гликогена в вагинальном эпителии у самок обезьян, морских свинок, мышей и сумчатых крыс и происходит резкое увеличение содержания глюкозы в слизи канала шейки матки к овуляции. Под действием прогестерона (гормон секреторной фазы) повышается содержание гликогена в эндометрии у приматов Lapan и Friedman (1950) нашли повышенное содержание гликогена и глюкозы в слизистой оболочке вагины у женщин в секреторной фазе цикла. К. А. Крутикова описала изменения pH влагалищного секрета у 140 женщин в связи с фазами циклов. pH было низким в первой фазе, ко времени овуляции увеличивалось, держалось на высоком уровне во второй половине цикла и снижалось перед менструацией. Отмечались большие индивидуальные различия в величинах pH, но направление изменений по фазам цикла совпадало почти у всех обследованных.
Л. Д. Тварчрелндзе (1964) производил окраску фиксированных мазков на гликоген и получил для пролиферативной стадии цикла гликогеновый индекс 40-50%, а для секреторной стадии - 70-80%. Во время менструации - снова снижение до 40-50%. В ановуляторных циклах таких волнообразных колебаний не прослеживалось. Автор предложил использовать определение гликогена на мазках в качестве теста для
55
суждения о функциональном состоянии яичников в цикле у женщин. Сходные результаты получили Lapan, Friedman (1950).
Нами (Юдина и др., 1968) проводилось исследование лактатдегидрогеназы (ЛДГ) в тканях органов у крыс в связи со сменой фаз половых циклов. Активность ЛДГ и ее изоэнзимов определялась электрофоретическим методом на полиакриламидном геле и последующим калориметрированием и денситометрированием. Было установлено, что в фазе течки увеличивается активность ЛДГ в гипофозе, гипоталамусе, надпочечниках и яичниках самок белых крыс (рис. 15). По фазам цикла во всех обследованных органах перестраивалась также активность изоферментов ЛДГ. Ко времени наступления течки увеличивалась активность изоэнзимов ЛДГ4 и ЛДГ5, т. е. наблюдался сдвиг в сторону преобладания М-субъединиц над Н-субъединицами в тетрамерах ЛДГ (рис. 15, Б).
Увеличение активности ЛДГ в фазе течки свидетельствует о том, что в стадии пролиферации активизируется анаэробный гликолиз, он достигает наибольших величин в конце стадии, когда наступает течка.
Для характеристики особенностей углеводного обмена в связи с половыми циклами изучали изменения в выделении с мочею у женщин лимонной кислоты - одного из своеобразных катализаторов в механизме окисления основного субстрата клеточного дыхания. Оказалось, что выделение лимонной кислоты уменьшается в течение пролиферативной стадии цикла, достигает наименьших величин в середине цикла и возрастает в секреторной стадии.
Не менее четкие изменения по фазам цикла были получены и при изучении фенольных веществ мочи. Концентрация фенольных соединений мочи (по оптической плотности) была высокой в первой половине овуляторного цикла и снижалась во второй. Если цикл был ановуляторным, различий в выделении фенольных веществ на протяжении цикла не наблюдалось. Было предложено использовать этот тест для определения овуляции в цикле (цит. по Schulz, 1954).
Белковый обмен
Уровень общего белка и его фракционный состав изучали в сыворотке крови у женщин в связи с циклом.
Для обобщенной характеристики белкового обмена исследовали азотистый баланс. Установили уменьшение выделения азота в дни менструации. В предменструальном периоде и после менструации у женщин отмечался положительный азотистый баланс. В дни регул выделение небелкового азота достигало 5-15 мг% (Schulz, 1954). Более полных сведений в литературе по этому вопросу нет.
Выделение серы у женщин в цикле проследил Senninger (1928). Он обнаружил вполне определенное выделение увеличения серы перед регулами. К сожалению, автор не следил за выделением серы на протяжении всего цикла, что делает полученную им картину неполной. Данная публикация до сих пор остается единственной (цит. по Schulz, 1954).
Обмен креатинина у женщин в связи с половыми циклами изучали Ryan и др. (1961). Они отметили изменения по фазам цикла, а именно высокий уровень креатинина мочи в пролиферативной стадии резко снижался в середине цикла и был более низким в секреторной стадии. Волнообразные изменения секреции креатинина были настолько яркими, что авторы предложили использовать их в качестве теста для определения овуляции в цикле. Имеются данные об увеличении мочевины к середине цикла и резком снижении ее во второй части цикла.
Изменения по фазам цикла в содержании коллагена в тканях матки у самок белых крыс определяли Erikson и Okey (1931) и обнаружили самый низкий уровень коллагена непосредственно перед овуляцией, когда вес матки был наибольшим. После овуляции увеличение общего колла-
56
гена сопровождалось возрастанием его концентрации и достигало пика* в стадии послетечки, а затем уменьшалось параллельно регрессу желтого тела. Отмечена значительная лабильность коллагена - изменения скорости аккумуляции коллагена наступали раньше, чем сдвиги в содержании остальных протеинов (цит. по Schulz, 1954).
У самок мышей по фазам астрального цикла значительно изменялось общее содержание белка в тканях гипофиза и яичников, увеличиваясь от диэструса к эструсу и снижаясь в метаэструсе, и надпочечников и тимуса, в которых количество белка достигало наибольших значении в метаэструсе. Были обнаружены различия в распределении водорастворимых белковых фракций этих же органов мышей, но здесь еще нужно уточнение и специальное изучение (Юдина и др., 1970).
С развитием представлений о нуклеиновых кислотах, осуществляющих синтез белков, появились работы, посвященные изучению нуклеиновых кислот в ядрах клеток, выстилающих половой тракт у самок, и изменений РНК и ДНК в связи с фазами половых циклов.
Ю-Ай и Чжан Цзяо-чен (1958) гистохимическим путем установили, что у самок белых крыс (половозрелых, 2-3-месячных) в фазе предтечки увеличивается содержание РНК в яйцепроводах, в эпителии вагины и в эндометрии матки, достигающее максимума в фазе течки, ко времени овуляции. ДНК повышается в фазах предтечки и течки. Изменения РНК в ганглионарных клетках вагины и шейки матки в разные физиологические периоды у самок крыс наблюдал Н. В. Оноприенко (1965). В эстральном цикле в фазе предтечки усиливалась интенсивность окраски пиронином. В течке она была наиболее выражена. В это же время на фоне сетчатой формы расположения РНК в протоплазме клеток обнаруживались зерна и глыбки РНК. Увеличение концентрации РНК в течке отмечалось не только в телах нейронов, но и в глиальных элементах. Накопление РНК свидетельствовало об активации белкового обмена.
Содержание РНК в клетках желез эндометрия в различных фазах цикла изучала И. Г. Патрикян (1967). Она определяла цитоплазматическую РНК и объемы цитоплазмы клеток в биопсиях на 10-13-й и 24-28-й дни цикла. Как и в работах других авторов, средняя концентрация РНК оказалась более высокой в пролиферативной стадии цикла (статистически достоверно). Но при этом объем цитоплазмы клеток эндометрия в пролиферативной стадии был небольшим, а в секреторной - намного увеличивался, а следовательно, и количество РНК было фактически почти вдвое большим в секреторной стадии, несмотря на низкую концентрацию его к цитоплазме. Обмен нуклеиновых кислот в эндометрии у женщин также изменялся на протяжении цикла. На 13-14-й и на 23-24-й дни цикла (перед овуляцией и во время максимальной активности желтого тела) достигали наибольшей концентрации растворимый в кислотах фосфор и фосфор рибонуклеиновой кислоты. Содержание ДНК-фосфора было наибольшим на 15-16-й дни цикла (во время овуляции) Активность РНК усиливалась к 19-20-му дням и становилась максимальной на 23-24-й день. Что касается метаболических сдвигов в различных фосфатных фракциях, то их пики также выпадали на 15-16-й и 23-24-й дни цикла (совпадали с пиками секреции эстрогенов). Синтез рибонуклеиновых кислот заметно возрастает в стадии пролиферации. В секреторной стадии сначала усиливается синтез дезоксирибонуклеиновой кислоты, а затем идет разложение ее.
Исследованиям ДНК в связи с половыми циклами посвящено несколько работ Drasher (1952) установил высокий уровень ДНК в матке в период течки и объяснил это увеличением числа клеток на единицу объема. Он нашел минимальное количество ДНК в ядрах клеток базального слоя эпителия вагины в стадии диэструса, увеличение в проэструсе, максимальное содержание в эструсе. Отметил, что ороговевание эпителия сопровождается уменьшением ДНК.
57
А. Е. Попова (1964), в отличие от многих авторов, обнаружила, что содержание ДНК может быть различным в одних и тех же клетках у разных индивидумов и в разных клетках одной и той же ткани. Оно изменяется в зависимости от физиологического состояния клетки. Цитоспектрофотометрическим методом А. Е. Попова определяла содержание ДНК в ядрах клеток и оптическую плотность ДНК.
Среднее содержание ДНК в ядрах клеток эндометрия у женщин изменялось по фазам полового цикла. В пролиферативной стадии содержание ДНК было более низким (0,92 х 10-7), чем в секреторной стадии (112 х 10-7). Оптическая плотность ДНК повышалась в первой половине цикла, снижалась перед овуляцией и снова увеличивалась во второй половине цикла. Изменялась и плотность ядер клеток. В стадии пролиферации в ядрах преобладали диплоидный и триплоидный уровни количества ДНК. В стадии секреторной преобладали ядра с три- и тетраплоидным количеством ДНК, появлялись клетки и с пятиплоидными уровнями.
Все это свидетельствует об интенсивном делении клеток в пролиферативной стадии цикла и замедлении деления клеток в секреторной стадии. Сходные изменения в содержании ДНК, оптической плотности ДНК и площади ядер обнаружила А. Е. Попова и в эндометрии крольчих.
Несмотря на некоторые расхождения, все эти данные говорят об интенсификации пролиферации ткани эндометрия, сопровождающейся усиленным синтезом ДНК. Смена фаз полового цикла обусловливает не только морфологические изменения эндометрия, но и регулирует те тонкие биохимические процессы на молекулярном уровне в клетке, которые предшествуют пролиферации ткани.
Электрофоретическое изучение белковых фракций слизи канала шейки матки в разные фазы менструального цикла показало наличие изменений в соотношении альбуминов, кислых и нейтральных мукопротеинов (Баблидзе, 1964).
Некоторые данные об изменении белков в плазме в связи с циклами были получены при изучении ретикулоэндотелиальной системы (РЭС) путем нагрузки краской (конгорот). Способность к поглощению краски изменялась в связи с половыми циклами. Она увеличивалась в первой половине цикла, снижалась во второй, была наименьшей перед менструацией. Возможно, что изменения поглотительной способности РЭС находятся не в связи с изменениями состояния РЭС, а в связи с изменением белкового состава плазмы. Уменьшение количества альбуминов и увеличение грубодисперсных белков перед менструацией и во время нее затрудняют поглощение краски РЭС.
Жировой обмен
Морфологический метаболизм липидов в вагинальных мазках в связи с циклом исследовал Д. К. Пападов (1965). При окраске мазков Суданом черным липиды отчетливо выделялись как в клетках эпителия, так и внеклеточные. Общее количество липидов увеличивалось в пролиферативной стадии цикла, достигало максимума к овуляции, снижалось после нее и снова увеличивалось к середине секреторной стадии. Перед стадией регенерации количество липидов снова снижалось. Изменения были настолько четкими, что автор предложил использовать их как дополнительные к дифференциальной окраске мазка, принятой в клинике для оценки функционального состояния яичников.
Ю-Ай и Чжан Цзяо-чен (1958) обнаружили, что у сам<






