Выделив волнообразность жизненных функций, которая охватывает весь организм и определяется течением половых циклов, в качестве особого экстрагенитального ритма жизнедеятельности, мы поднимаем вопрос
80
о том, какие силы вызывают эту ритмичность, что управляет ею и каковы конкретные механизмы экстрагенитальных циклов.
Согласно фактическим данным, в течение половых циклов активно меняется деятельность нервной системы, общая реактивность и резистентность организма. С другой стороны, на течение и структуру половых циклов большое влияние оказывают внешние факторы, которые стимулируют половые циклы, тормозят их развитие в целом и течение отдельных фаз и, наконец, могут приводить к полному выключению.
Поскольку речь заходит о внешних влияниях на ход половых циклов, необходимо признать ведущую роль за центральной нервной системой, связывающей внешнюю среду организма с его внутренней средой. Непосредственными регулировщиками циклических изменений являются тропные и половые гормоны, и это тоже бесспорно, так как всякая цикличность у самок исчезает после овариектомии.
Нельзя считать, что механизм создания и регуляции волнообразности жизненных функций выяснен до конца, но обилие экспериментальных данных и обобщения, выполненные физиологами репродуктивной системы и эндокринологами, делают возможным получение общего представления о причинах и механизмах, лежащих в основе этого своеобразного биологического ритма.
В литературе имеется огромное количество работ, демонстрирующих изменения половой функции животных, и в частности половой цикличности самок, под влиянием меняющихся функциональных состояний желез внутренней секреции. Не меньше работ посвящено изменениям половой функции и половых циклов у самок под действием различных частей нервной системы, вплоть до коры больших полушарий.
Значительно меньше исследований было выполнено по вопросам изменений, происходящих в различных железах внутренней секреции в связи с половыми циклами самок.
Надпочечники. Нами была выполнена работа на крысах самках, у которых определяли динамику лактатдегидрогеназы в некоторых органах и попутно - вес самих органов в связи с половыми циклами (Юдина и др., 1968). Фазы полового цикла определялись у крыс по вагинальным мазкам, окрашенным раствором Люголя по Papanicolaou (1932).
В третьей фазе цикла, т. е. в течке, вес надпочечников у всех обследованных самок увеличивался по сравнению с фазами покоя и после течки. Видимо, специальные исследования, если они будут предприняты, обнаружат различия в связи с течением половых циклов в соотношении корковой и мозговой частей надпочечников, в соотношении слоев коры и их строения у самок разных видов животных, обезьян и у человека.
Отчетливые различия в строении коры надпочечников у коров были прослежены по фазам полового цикла (Лемешко, 1958). В межтечковом периоде клубочковая зона узкая, хорошо отграничена, липидов содержит мало. Еще меньше липидов в мозговом слое. Аскорбиновой кислоты мало в клубочковом слое и много в пучковом и сетчатом.
Предтечка характеризуется широкой клубочковой зоной, граница ее с пучковой сглажена. Липидов много, особенно в клубочковой зоне. Увеличилось и содержание аскорбиновой кислоты. В остальных слоях количество липидов и аскорбиновой кислоты уменьшилось. В течке все эти особенности усиливались. После течки - все изменения возвращались к исходным.
Вопрос о динамике выделения 17 КС у женщин в связи с течением половых циклов следует считать нерешенным. Большинство авторов не обнаружили фазовых изменений в выделении этих гормонов. Ряд авторов нашли волнообразные изменения экскреции 17 КС по фазам полового цикла у женщин. Как правило, изменения были невелики, увеличение
81
выделения обнаруживалось в середине цикла, к овуляции. Об увеличении экскреции 17 КС у женщин во вторую и третью неделю менструального цикла сообщил Е. И. Кватер (1967). Есть наблюдения, демонстрирующие нарастание экскреции 17 КС у женщин в пролиферативной фазе цикла с максимумами выделения на 6-9-й и на 15-19-и дни, считая от первого дня менструации (Нестерова, 1961). Циклические изменения 17 КС по дегидроэпиандростерону ДНА у женщин обнаружил Джеймс (James, 1961). ДНА был низким в начале и в конце цикла и возрастал до 1,9-3,7 мг/24 часа к середине цикла. Фракционирование мочевых С19 стероидов у женщин показало отсутствие связи в соотношении фракций с фазами полового цикла (Каримов, 1965) или меньшее количество андростерона в первой фазе цикла (Какушкина, Орлова, 1960).
Отсутствие определенных изменений в выделении 17 КС в связи со сменой фаз половых циклов у женщин данных, демонстрирующих влияние яичников на функцию надпочечников.
Волнообразная динамика выделения 17 КС у самок морских свинок была установлена Brook с соавторами (1960): количество 17 КС мочи отчетливо было наиболее высоким в фазе течки, во все остальные периоды цикла уровень выделения оставался невысоким. В выделении 17 кетогенных стероидов связь с циклом обнаружена не была.
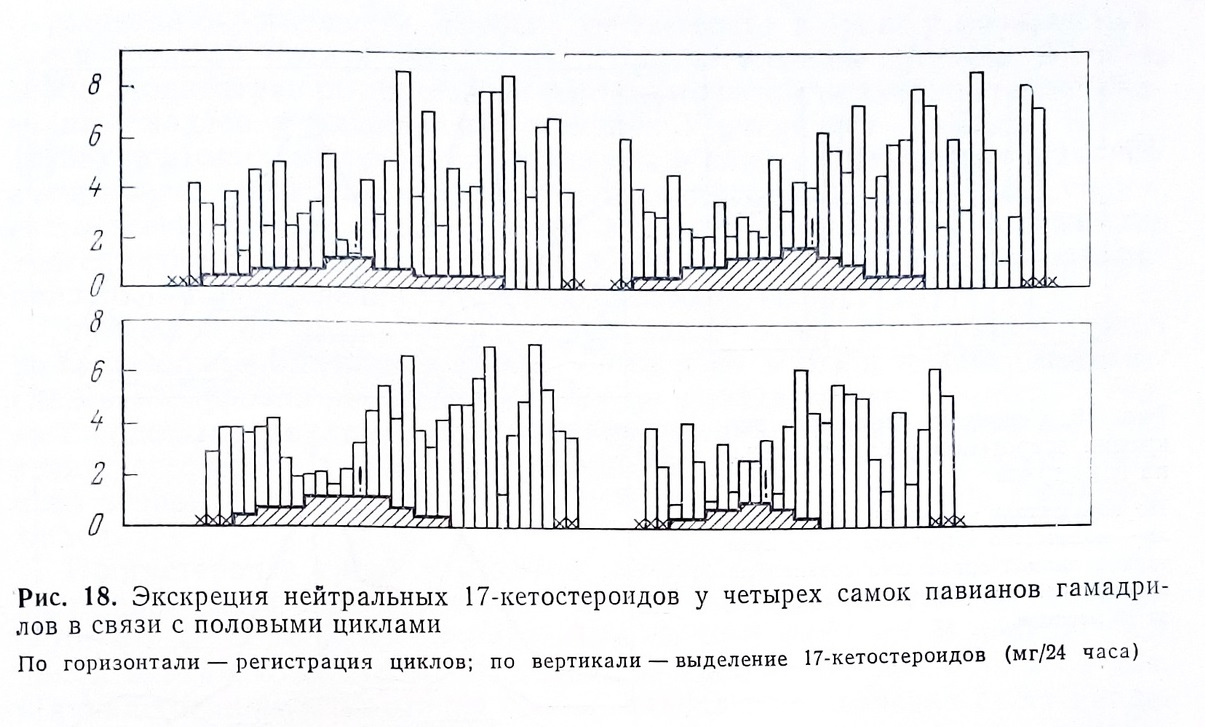
Тщательное, день за днем, определение выделения суммарных 17 КС у самок павианов гамадрилов позволило установить отчетливые изменения уровня экскреции этих метаболитов по фазам половых циклов. У четырех самок было обследовано по два полноценных цикла и установлено, что уровень выделения, несмотря на значительные колебания день ото дня, был более высоким во второй половине цикла (рис. 18) (Алексеева, 1974).
Яичники. У половозрелых женщин количество выделяемых эстрогенов находится в связи с развитием и сменой фаз половых циклов. Хотя абсолютные значения экскреции эстрогенов, полученные с помощью различных методов определения, значительно различаются, все же динамика выделения эстрогенов, как суммарных, так и каждой из фракций в отдельности, совпадают у разных авторов.
Некоторое разногласие существует лишь в отношении количества пиков экскреции. По одним авторам, их в женском цикле два - в период овуляции и в середине секреторной фазы. По другим - три пика; в этих случаях дополнительный пик экскреции определяется на 6-7-й день после первого дня менструации. В 28-дневном цикле этот пик соответствует началу пролиферативной стадии цикла. Установлено что эстрогены секретируются в основном яичниками - от 60 до 400 мкг/24 часа. Надпочечники в норме продуцируют лишь незначительную часть эстрогенов, не более 15-30 мкг/24 часа (Diczfalusy et al., 1961).
Близкие значения величины продукции эстрогенов яичниками у здоровых женщин получили с помощью изотопных методов.
Braun и др. (1958) вычисляли количество продуцируемых организмом эстрогенов за сутки по величине их суточной экскреции исходя из того, что при введении эстрогенов не более 16% их выводится с мочой. По его данным в фолликулиновую фазу цикла яичник продуцирует от 70 до 340 мкг/24 часа, а в фазу желтого тела - меньше, не более 160-200 мкг/24 часа. Методом изотопного разведения для фолликулиновой фазы были найдены величины секреции от 35 до 288 мкг/24 часа, а для лютеиновой фазы - от 60 до 187 мкг/24 часа
Сходство в структуре менструального цикла у самок различных приматов в большой степени определяется сходным течением овариального цикла. Доказано, что у самок обезьян яичники по своему строению и по функции весьма сходны с яичниками человека.
В яичниках у половозрелых обезьян, как и у женщин, растут и атрезируются примордиальные фолликулы, на фоне их несколько фоллику-
82
лов в каждом цикле созревают, но в норме овулирует лишь один из них. К слову можно напомнить, что многоплодие реже встречается у обезьян, чем у человека. Случаи рождения двойни почти в 10 раз более редки (по наблюдениям в Сухумском питомнике обезьян).
Выделение эстрогенов у макаков резусов с мочой биологическим тестированием изучали Dorfman и Wagenen (1941). Они установили, что у неполовозрелых самок и самцов различий в выделении эстрогенов и андрогенов нет. У половозрелых самок выделение эстрогенов с мочею достигает от 2 до 12 и. е. за 24 часа. У самцов - от 1.1 до 2,5 и. е. за 24 часа.
Отметили большую вариабильность выделения у самок, очевидно связанную с половыми циклами. У беременных самок количество выделяемых суммарных эстрогенов значительно увеличивается, достигая 10-460 и. е. за 24 часа (Алексеева, 1974).
Allen с сотрудниками изучали выделение тиелина у беременных шимпанзе (1935) и у шимпанзе небеременных (1936), определяя эстрогены в моче биологическим путем. Они показали, что у самок шимпанзе с мочой выделяются эстрогены. Количество их невелико в начале цикла, возрастает с развитием набухания половой кожи, достигает наибольших величин в дни максимального набухания половой кожи и снова уменьшается в секреторной стадии цикла. У беременных шимпанзе выделение эстрогенов с мочой во много раз больше, чем у небеременных.
У павианов гамадрилов выделение суммарных эстрогенов и их отдельных фракций изучалось химическими методами (Алексеева, 1961, 1963).
Было установлено, что уровень выделения суммарных эстрогенов у молодых обезьян самок и самцов не различается до приближения периода созревания. Перед первым набуханием половой кожи у молодых самок выделение эстрогенов приобретает волнообразный характер. Мы наблюдали это у неполовозрелой самки павиана Арапки и у других. У неполовозрелого самца Ларна за такое же время наблюдения отмечались незначительные колебания экскреции, без выраженных подъемов (Алексеева, 1974).
Прослежены были также суточные и сезонные колебания экскреции суммарных эстрогенов (рис. 19).
У четырех половозрелых самок павианов гамадрилов выделение эстрогенов суммарных и по фракциям определялось на протяжении 2-3 половых циклов. Установлено, что выделение эстрогенов повышается в
83
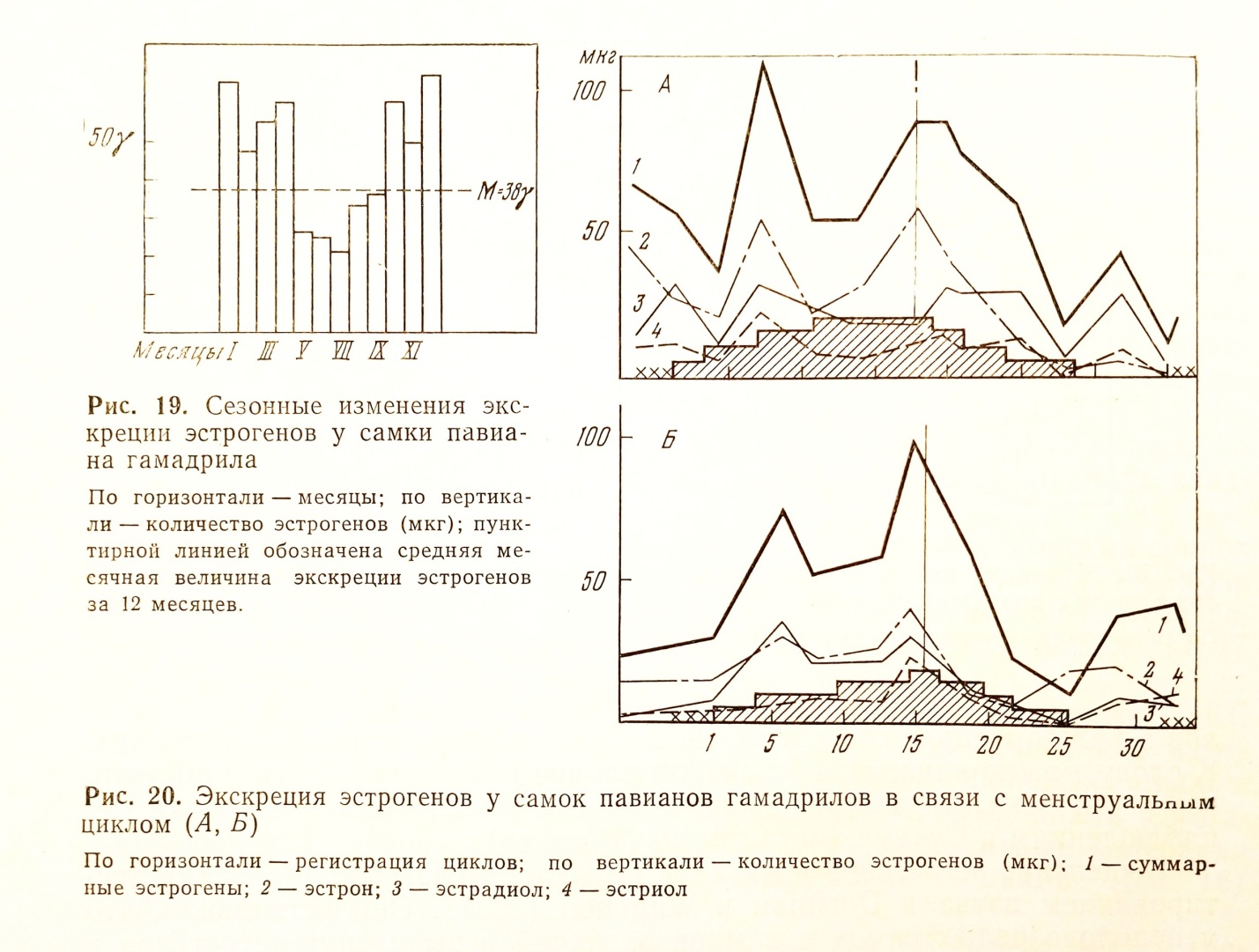
середине цикла, на границе второй и третьей трети секреторной стадии цикла. В ряде циклов определяется еще один пик эстрогенов - в начале пролиферативной стадии. Этот пик экскреции отмечался, по нашим наблюдениям, в случаях удлинения половых циклов за счет его первой половины.
Приводим две типичные кривые экскреции эстрогенов у самок павианов гамадрилов (рис. 20, А, Б).
У женщин с нормальным циклом количество прогестерона в фолликулярной фазе составляет 0,10-0,42 мкг/100 мг плазмы, к овуляции повышается до 1-2,06 мкг/100 мг, затем снижается до 0,70 мкг/100 мл в лютейновой фазе (Riondel et al., 1965).
У обезьян определение прегнандиола сначала дало отрицательные результаты, что послужило обоснованием для заключения о принципиально различных путях метаболизма прогестерона у обезьян и человека.
Первыми установили отсутствие прегнандиола и его эпимеров в моче у самок макаков резусов Westfals и Bukston (1939), Marker и Hartman (1940). Авторы пользовались весовым методом Элеоноры Веннинг, значительно менее чувствительным, чем современные методы, и не нашли даже следов прегнандиола в моче у самок макаков в цикле, при беременности и даже после нагрузки большими количествами прогестерона. Одновременно в параллельных условиях они определяли прегнандиол и находили его в моче у женщин. В 1961 г. Schort, Eckstein, не повторяя определения прегнандиола у макаков, предложили свои объяснения причин отсутствия прегнандиола в моче у макаков, что позволило резко выделить их из всех обследованных на прегнандиол мочи животных и послужило основанием для заключения о принципиальных различиях в метаболизме прогестерона у обезьян (макаков) и человека.
Факт представлялся тем более странным, что у макаков, как и у человека и у многих животных - у лошадей, коров, овец, собак и пр., удаление яичников не прерывает беременности. Очевидно, и у них прогестерон вырабатывается тканями плаценты в достаточных количествах для
84
сохранения беременности. Уровень прогестерона в крови у макаков оказался тоже сходным с установленным для женщин (Forbs, Hooker, 1950). Концентрация прогестииа в амниотической жидкости также оказалась сходной у макаков и у женщин. Уточняя эти данные, Schort (1958) повторил исследование плаценты и нашел в плаценте у макаков значительно более низкое содержание прогестерона, чем нашли упомянутые выше авторы. Он определил для плаценты макаков содержание прогестерона в 35-88 мг/кг ткани, а для плаценты женщины он при параллельном определении установил 2220 мг/кг ткани.
Более низкий уровень прогестерона нашел Schort и в плазме макаков по сравнению с плазмой человека. Forbs и др. в 1950 г. сообщили об исследовании уровня прогестерона в крови у самок макаков резусов в связи с половыми циклами. Во' время менструации и сразу после нее уровень прогестерона в крови был низким, он повышался в середине цикла и во второй фазе держался на высоком уровне, но в конце фазы снижался.
Прогестерон в крови у макаков резусов определяли в связи с половыми циклами и беременностью и нашли, что в плазме у макаков содержится от десятых долей микрограмма до 1-5 мкг/1 мл. В первой фазе цикла низкая концентрация, повышение происходит резко, за 2-3 дня, в начале секреторной стадии цикла, наибольшие значения были найдены на 18-20-й дни цикла (если за начало цикла считать начало набухания половой кожи). Если цикл переходил в беременность, то увеличение содержания прегнандиола продолжалось и концентрация его достигала 7-8 мкг/мл на первом месяце и далее изменялась волнообразно, то повышаясь, то понижаясь, на всем протяжении беременности (Ramirez, 1964).
В то же время в моче самок шимпанзе прегнандиол был обнаружен даже весовым методом (Fisch et al., 1942). У беременных шимпанзе авторы определили величину экскреции в 2 мг за сутки в среднем. Fisch выделил 48 мг свободного и 30 мг измененного прегнандиола из мочи кастрированной самки шимпанзе сразу после введения ей 3 гдезоксикортикостерона.
У других представителей низших узконосых обезьян (у медвежьих павианов) du Toit (1956) исследовал экскрецию прегнандиола ежедневно на протяжении полового цикла и нашел, что в первой фазе выделение не превышало 0,4 мг в сутки, в дни овуляции увеличилось до 0,7 мг и к концу секреторной стадии снова снизилось до 0,2 мг/24 часа.
Детальное изучение динамики выделения прегнандиола методом Клоппера, Мичи и Брауна, включающего ацетилирование и двукратное хроматографирование на окиси алюминия (1955) было проведено в Сухумском питомнике обезьян на самках павианов гамадрилов (Алексеева, Григоренко, 1964). В опыте было пять половозрелых самок с нормальными циклами, прирученных к содержанию в мочесборных клетках. Определения прегнандиола производились в дни, когда происходили изменения в течении полового цикла, регистрируемые по описанной выше схеме.
В фолликулярной фазе было проведено по четыре определения: в день.начала цикла - первый день набухания половой кожи, в дни, когда набухание достигало величины, оцениваемой двумя и тремя баллами, и непосредственно перед овуляцией. В лютеиновой фазе было сделано по 3-4 определения, последнее - перед менструацией.
Отмечены большие индивидуальные различия в уровне экскреции прегнандиола. Так, у обезьяны Неры выделение не превышало 60 мкг/24 часа, а у остальных выделение достигало 190 и 220 мкг/24 часа. Отчетливо проявилась связь между величиной экскреции и фазами половых циклов. В пролиферативной стадии цикла у всех трех самок выделение прегнандиола было сначала небольшим, в пределах 18-30 мкг, затем увеличи-
85
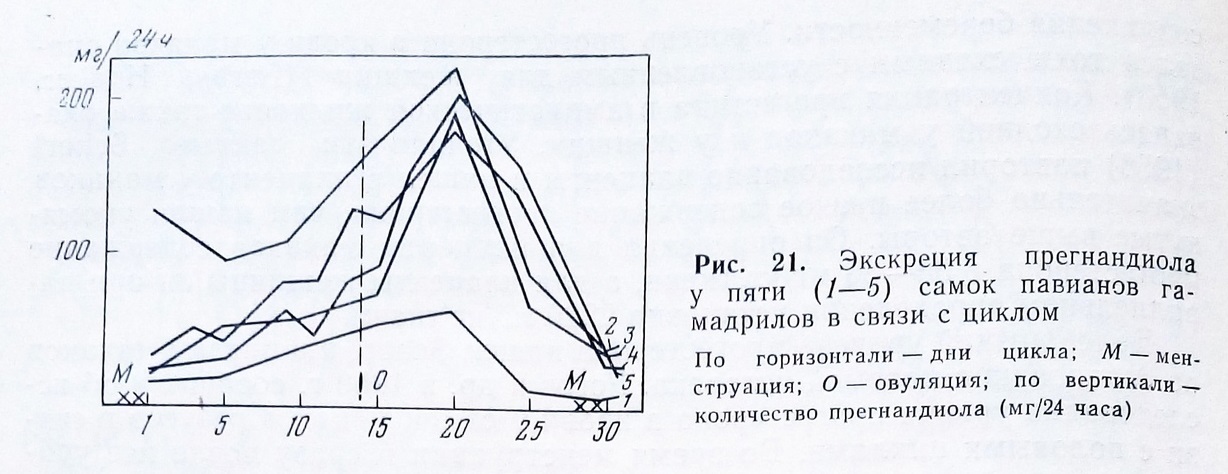
валось до 60-135 мкг к концу стадии. После овуляции выделение прегнандиола было более высоким, чем до нее, и в лютеиновой фазе цикла самки выделяли с мочею от 60 до 220 мкг в сутки. Самые большие величины выделения были получены у самок в дни, когда завершалось отбухание половой кожи. За несколько дней до конца лютеиновой фазы выделение резко снижалось.
Параллельно нами определялась экскреция прегнандиола у женщины и, как выяснилось, динамика выделения оказалась весьма сходной у женщины и у самок павианов. Но у женщины за сутки выделялось значительно больше прегнандиола - в первой фазе от 0,5 мг до 1,2 мг, во второй фазе от 4-5 мг до 1,9 мг. Самки павианов выделяли меньше прегнандиола не только абсолютно, но и относительно: на 1 кг тела обезьяны выделяли от 10 до 14 мкг, тогда как у женщины на 1 кг веса тела выделялось не менее 90 мкг (рис. 21).
Соматотропный гормон определялся (Алексеева, 1974) в гипофизах у забитых в разных фазах половых циклов крыс и мышей с помощью микроэлектрофореза на полиакриламидном геле последующим сканированием колонок на МУФ-4 и вычислением процентного соотношения дисков, образованных на колонке белковыми фракциями аденогипофиза. При этом мы определяли относительные количества соматотропного гормона и прослеживали изменения его по фазам полового цикла (рис. 22).
Как видно на графике, количество соматотропного гормона нарастало у крыс и у мышей в предтечке и было наибольшим в течке (статистически достоверно).
Установлено, что у самок крыс и мышей по фазам половых циклов изменяется относительное количество пролактина, а именно оно увеличивается в фазах течки и послетечки (рис. 22, Б) (Алексеева, 1974).
Приведенные данные об изменениях функций желез внутренней секреции в связи со сменой фаз половых циклов у самок животных и обезьян еще фрагментарны (см. рис. 26, Б).
Однако они указывают на принципиальное сходство с хорошо изученными фазовыми изменениями функций желез внутренней секреции у женщин в связи с менструальными циклами и свидетельствуют об активном участии желез внутренней секреции в формировании экстрагенитального ритма жизнедеятельности организма женских особей.
Остаётся открытым вопрос о существовании и локализации полового центра в ЦНС. Нами были проведены опыты, которые позволяют высказать некоторое суждение по этому вопросу.
В экспериментах по воссозданию половых циклов у павианов-кастратов было показано, что необходимым и достаточным условий для цикличности оказывалось введение половых гормонов. Получило подтверждение предположение о наличии особого высшего нервного полового центра, бисексуального в своих потенциях. Развитие в направлении женского циклического типа реакций определялось фолликулярным гормоном. В отсутствие этого гормона развивался мужской ациклический тип реакций.
86
Эти положения получили подтверждение в опытах на крысах (Gorski, Wagner, 1965). Авторы показали, что половые различия в типах секреции гипоталамических релизинг-факторов (мужской - ациклический и женский циклический тип секреции) формируются у крыс в первые 10 дней после рождения. Если у новорожденного крысенка приостановить резерпином секрецию тестикулов или кастрировать его и пересадить на 3-й день жизни яичники, то у него можно получить циклический женский тип секреции гипоталамических факторов и гормонов гипофиза.

В опытах на самках обезьян был проведен и получил теоретическое обоснование взгляд на одну из актуальных проблем женской клиники — на функциональные маточные кровотечения как на результат потери ритма во взаимодействии высшего нервного центра, гипоталамуса и гипофиза, с одной стороны, и половой сферой - с другой, как на нарушения цикличности выработки половых гормонов. Успешность опытов с длительным введением гормонов бесплодным самкам с ациклическими менструациями позволила обосновать рациональную гормональную терапию функциональных маточных кровотечений у женщин и дать практические рекомендации о безвредности повторения курсов гормонотерапии в течение 6 и более месяцев (Кватер, 1967).
Было показано, что регулярным введением половых гормонов у самок обезьян можно получить менструальные циклы заранее намеченной продолжительности. Длительным применением гормонов можно закрепить «воспитанный» ритм на более или менее длительный срок. Полученные в опытах циклы оказались полноценными, они сопровождались восстановлением экстрагенитальной цикличности и способности к зачатию и деторождению. В заключение следует обратить внимание на общебиологический аспект экстрагенитальной цикличности. По сути дела все рассмотренные выше волнообразные изменения жизнедеятельности в связи с половыми циклами сводятся к изменениям реактивности. Идет ли речь об изменениях проницаемости капилляров или pH различных тканей, о выделении и скорости секреции гормонов или возрастающей активности ферментов, динамике темповой адаптации зрительного прибора или морфологических изменениях в структуре энергетических фабрик клетки митохондрий и т. п. - результирующей всех проявлений экстрагенитальной цикличности, всех этих частных взаимосвязанных и взаимовлияющих из-
87
менений реактивности являются изменения общей реактивности и резистентности женских особей. В последние годы пробудился интерес к половым различиям в реактивности. В 1965 г. А. С. Султаналиев показал половые различия в чувствительности к повреждающему действию электротока. Из группы в 200 мышей по 100 каждого пола погибло 62 самца и 43 самки, причем чувствительность самцов (по степени поражения) оказалась более высокой.
Сообщают о более высоком и быстром подъеме уровня кортикостероидов в плазме крови у самок в ответ на стрессовые воздействия и о том, что период полураспада кортикостерона у самок крыс значительно короче, чем у самцов (13 мин. вместо 20 мин.) (Kitay, 1961).
Значительные различия в метаболизме гипоталамуса установили Моguilevski с соавторами (1968). Среди жителей Аляски у женщин обнаружили более низкую концентрацию цезия137 в тканях тела и более медленное накопление этого изотопа с возрастом (Boni, 1969). Половые различия в реакциях на внешние воздействия в том числе и на патогенные, давно известны в клинике.
Установлено, что женщины чаще заболевают тифами, скарлатиной,, дифтерией а мужчины - полиомиэлитом, гриппом, эпидемическими энцефалитом и менингитом. Из сердечно-сосудистых заболеваний для женщин более характерны анемии, тромбозы, эмболии, для мужчин - гипертония, инфаркт миокарда, эндо-мио-перикардиты, гемофилия, атеросклероз (Schulz, 1954; Горизонтов, 1966). Специальное изучение смертности от лейкозов в г. Минске и области за 1958-1967 гг. показало, что заболевания и гибель от острых форм лейкоза, от хронического лимфо-лейкоза и лимфогрануломатоза наблюдаются преимущественно среди мужского населения. Даже мальчики до 5-летнего возраста заболевали острым лейкозом вдвое чаще, чем девочки (Данилов, Ненадкевич, 1971). Почти все заболевания органов дыхания чаще встречаются у мужчин: гангрена легких, энфизема, пневмонии, бронхиты, лимфогрануломатоз, бронхоэкстазия, tbc. Правда, за последние годы случаи рака легкого участились и у женщин, видимо, в связи с курением. У женщин чаще, чем у мужчин, отмечаются холециститы, желудочно-кишечные заболевания, у мужчин - язвы желудка и 12-перстной кишки. Разнообразнее у женщин нервная клиника - неврозы, психозы, истерии. Больше случаев диабета, ожирения. Из эндокринопатий у мужчин встречаются чаще Адиссонова болезнь, карликовый нанизм и гигантизм, у женщин - базедовая болезнь, микседема.
К сожалению, лишь в отдельных случаях обращалось внимание на физиологическое состояние женщин и заболевания сопоставляли с половыми циклами. Оказалось, что такие преимущественно женские заболевания, как скарлатина и дифтерия, чаще начинаются в дни, когда наиболее снижаются бактерицидные свойства крови (третий пик экскреции эстрогенов?). Вероятно, имеется связь и между другими заболеваниями и экстрагенитальными циклами женских особей. Но таких наблюдений нет.
Половые различия ярко выражены и в таком интегративном показателе реактивности, как преимущественное выживание и большая продолжительность жизни женских особей. Широко известная для животных и обезьян, эта закономерность установлена и по отношению к человеку. Механизм формирования этих различий ещё непознан. Он, очевидно, связан с половыми различиями, с половым диморфизмом, а это значит, прежде всего, с экстрагенитальной циклической динамикой жизненных функций женских особей.
88
Глава III





