Жизненный цикл рыб, т.е. вся жизнедеятельность организма в течение онтогенеза от момента оплодотворения до естественной смерти, распадается на различные периоды, каждый из которых характеризуется определенными морфологическими и физиологическими особенностями. В жизненном цикле рыб выделяют следующие периоды: эмбриональный (зародышевый), личиночный, мальковый, ювенильный (юношеский), взрослого организма, старческий.
Эмбриональный (зародышевый) период характеризуется эндогенным питанием (за счет собственного желтка), длится от момента оплодотворения до момента перехода на внешнее (экзогенное) питание и подразделяются на 2 подпериода: собственно эмбриональный, когда развитие совершается внутри яйцевой оболочки, а у живородящих рыб протекает внутри тела самки; предличиночный (свободного эмбриона), когда выклюнувшиеся свободноживущие особи питаются за счет желтка (рис. 50).

Рис. 50. Развитие воблы:
1 – икринка; 2 – предличинка; 3 – личинка; 4 – малек
Личиночный период характеризуется переходом на активное внешнее питание с сохранением личиночных органов и признаков. По внешнему и внутреннему строению организм еще далек от взрослого. В частности – плавниковая складка еще не дифференцирована на зачатки парных и непарных плавников. Мальковый период характеризуется тем, что по внешнему строению организм приобретает сходство со взрослым. Непарная плавниковая складка дифференцируется на плавники. Закладывается чешуя, дифференциация пола началась, но половые органы неразвиты. Малек – это молодая сформировавшаяся рыбка данного года рождения.
Ювенильный (юношеский) период характеризуется усиленным развитием половых желез, но рыбы еще неполовозрелые. Начинают развиваться вторичные половые признаки (если они имеются).
Период взрослого организма начинается с момента наступления половой зрелости, и рыбы обладают всеми признаками, характерными для полностью сформировавшегося организма.
Старческий период характеризуется замедлением роста или полным прекращением его. Рыба теряет способность размножаться.
Теория этапного развития рыб, позволившая изучить периоды, этапы, стадии развития и их продолжительность у многих видов рыб, была разработана В. В. Васнецовым, затем дополнена и развита другими исследователями.
Периоды – это довольно длительные интервалы индивидуального развития, слагающиеся из последовательных этапов.
Этапами называются такие интервалы в развитии рыбы, в течение которых происходит рост и медленные, постепенные изменения организма, однако никаких существенных изменений ни в строении, ни в физиологии, ни в поведении рыбы не совершается. Сохраняются ведущие взаимоотношения организма со средой, остается тот же способ движения, захвата пищи и т.д. (Васнецов).
Только после достижения определенной меры количество переходит в новое качество, и организм вступает в следующий этап. Смена этапов у рыб происходит в течение всей жизни. В первые периоды этапы обычно короткие, продолжающиеся от нескольких часов до нескольких суток, а у взрослых рыб длительность этапов может измеряться годами. Переход от одного этапа к другому осуществляется скачкообразно с резкими изменениями в строении и обычно происходит при достижении определенных размеров.
Скачки (морфологические изменения) совершаются очень быстро, иногда менее чем за 3-4 ч, и неразрывно связаны с изменением биологии. Незначительные морфологические изменения, отражающие каждый момент развития организма, называются стадиями.
У трески, например, период собственного эмбрионального развития подразделяют на несколько стадий (рис. 51).
Ia стадия – начало дробления (от 2 до 8 бластомеров).
I стадия – дальнейшее дробление, обрастание желтка зародышевым слоем - гаструляция. Длится до появления зародышевой полоски.
II стадия – образование зародышевой полоски. Продолжается дифференцировка туловищных миотомов. Стадия заканчивается отделением хвостовой почки от желтка.
III стадия – неоформившегося эмбриона. Начинается от момента обособления хвоста и продолжается до охвата эмбрионом всего желтка.
IV стадия – оформившегося эмбриона. Начинаются пульсация сердца и подергивание эмбриона. Длится от момента охвата эмбрионом всего желтка до выхода его из икринки. На этой стадии у эмбрионов многих рыб глаза пигментированы.
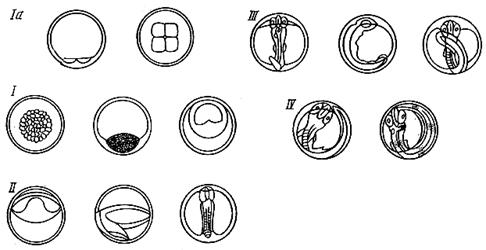
Рис. 51. Стадии эмбрионального развития трески:
Ia – начальная стадия дробления (от 2 до 8 бластомеров); I – стадия дробления и обрастания; II – стадия образования зародышевой полоски; III – стадия неоформившегося эмбриона; IV – стадия оформившегося эмбриона
Однако не все авторы вкладывают в термины «стадия», «этап» одинаковое содержание, что затрудняет сравнение данных о развитии рыб. К тому же у разных рыб наблюдается разное количество этапов развития, причем содержание их также различно. У щуки, например, три личиночных этапа развития, у литофилов – четыре, у фитофилов – шесть.
Знание этапов развития рыб необходимо для совершенствования биотехники разведения и перевозки икры и личинок, понимания причин колебаний численности популяций.
Контрольные вопросы и задания
1. Способы размножения у рыб.
2. Единовременное и порционное икрометание.
3. Шкалы зрелости половых продуктов.
4. Коэффициент и индекс зрелости половых продуктов.
5. Плодовитость (потенциальная, конечная, индивидуальная или абсолютная, относительная, рабочая и др.). Способы определения плодовитости.
6. Жизненный цикл рыб (периоды, этапы, скачки).
VII. МЕТОДЫ ОПРЕДЕЛЕНИЯ ЧИСЛЕННОСТИ РЫБ
Абсолютные методы определения численности стада
(на единице площади или в единице объема)
Одной из серьезных методологических задач современной экологии в целом и гидроэкологии в частности остается проблема количественного учета населения и его динамики под воздействием факторов среды. В ихтиологии она актуальна с момента становления ее как науки до настоящего времени. Уже накоплен огромный опыт определения абсолютной и относительной численности рыб, описано большое число методик и способов расчета плотности ихтиомассы в водоемах разного типа и на разных биотопах одного водоема, однако проблему еще нельзя считать решенной.
Существующие в настоящее время методы определения общей или абсолютной численности рыб условно могут быть разделены на 2 группы. К первой из них относятся все методы прямого учета числа особей, находящихся над определенной площадью дна или в единице объема водной толщи. Этот учет осуществляется путем отлова икры, молоди или взрослых рыб различными орудиями лова, по данным наблюдений с воздуха и аэрофотосъемки, путем расшифровки эхолотных записей и подводного фотографирования, прямыми подсчетами рыб, проходящих через фиксированный створ реки или рыбохода и особей, погибших в процессе химической очистки водоемов.
Вторую группу составляют расчетно-аналитические методы и методы математического моделирования численности популяций, основанные на прямых и косвенных показателях пополнения и убыли (естественной и промысловой) и данных мечения.
Основным недостатком существующих методов определения численности рыб в водоемах оказывается то, что они могут быть применимы или только в строго определенных экологических ситуациях (например, все методы прямого учета), или по принципу применимы широко, но неточны.
Основные трудности использования прямого счета заключаются в технической сложности облова за короткие сроки значительной акватории и в непостоянстве коэффициентов уловистости существующих орудий лова, связанном с пятнистостью распределения скоплений рыбы, а также с возрастными и сезонными изменениями жизнедеятельности образующих популяции группировок особей.
Общий недостаток существующих расчетно-аналитических методов определений и математических моделей численности заключается в том, что все они пока построены на основе весьма условных представлений о фактическом ходе пополнения и убыли особей в изучаемых популяциях и не учитывают в нужной мере комплексного воздействия на динамику численности факторов среды.
Расчет предполагаемого улова при применении методов определения относительной величины запаса зависит не только от динамики возрастного состава, то есть от урожайности поколений и темпа их изъятия промыслом, но и от степени изменения биологических свойств популяции (темпа роста, созревания, продолжительности жизненного цикла и др.). Эти изменения регулируются условиями внешней среды и зависят от численности популяций, ежегодно меняющейся в соответствии с эффективностью воспроизводства, в некоторых случаях – с интенсивностью промысла.
Расчет промыслового запаса ведется в следующей последовательности: пополнение, рост, естественная убыль, промысел.
Пополнение рассчитывается на основании количественного учета молоди и экстраполяции полученных данных с аналогичными результатами прошлых лет.
Количественный учет молоди проводится по средним уловам выметанных икринок с учетом этапов и стадий развития, по средним уловам личинок в период перехода их на активное питание, по средним уловам сеголеток, по средним уловам младших возрастных групп (до наступления половой зрелости), после того как прошел массовый отход икринок и личинок.
При этом преимущественное значение имеет учет сеголетков (или младших возрастных групп), так как между относительной величиной поколения, учтенной по промысловому возврату, и средним уловам сеголеток данного поколения имеется тесная связь.
Первые попытки определения численности рыб по пелагической икре были сделаны на треске в 1892 г. Гензеном и Апштейном (цит. по Никольскому Г.В.) (Hensen, Apstein, 1895).
Облов пелагической икры производился планктонными сетями с мельничным газом N3.Определив сроки икрометания, район и глубины распределения икры и этапы развития икры и личинок, подсчеты производили умножением количества икры, приходящейся на 1 м2, на общую площадь моря, где встречалась икра.
Затем, принимая во внимание плодовитость одной самки, определялось количество отнерестившихся самок: по соотношению самцов и самок вычислялась величина нерестового стада (уравнение 39, 40)
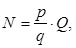 (39)
(39)
где N – общее количество выметанных икринок;
р – среднее число икринок в улове;
q – обловленный объем;
Q – общий объем воды в исследуемом районе.
 (40)
(40)
где St – величина нерестового стада;
S – соотношение полов;
n – средняя плодовитость (ИАП);
N – общее число выметанных икринок.
Для получения относительных величин численности трески этот метод, где он был впервые использован, вполне применим, однако для определения абсолютной численности нерестового стада этим способом требуется учет столь многих поправок, что практически этот метод оказывается применимым лишь в редких случаях.
Погрешности этого метода связаны с недоучетом гибели развивающейся икры, с неравномерностью распределения ее по глубинам, недостатками в конструкциях орудий лова, что отражается на их уловистости (отношении икринок, находящихся в единице объема, к числу икринок попавших в орудие лова). (Уравнение 41, по Гензену, Апштейну, 1897).
 (41)
(41)
где Р – величина запаса;
N – количество икры;
S – соотношение полов;
n – средняя индивидуальная абсолютная плодовитость (ИАП);
k 1 – смертность икры до вылупления;
d 1 – время между нерестом и взятием проб;
Т – период инкубации в сутках.
Попытки определения численности сельди у берегов Норвегии по донной икре предпринимались Руннстремом (Runnstrom, 1941).
Регулярные обследования нерестилищ охотоморской сельди, откладывающей икру на донную растительность в прибрежье, с целью определения в начале нерестового запаса, а затем и численности проводились Охотской лабораторией ТИНРО с конца пятидесятых годов прошлого столетия (Тюрнин, 1965).
Нерестовый ареал охотской сельди только по западному побережью Охотского моря составляет более 500 миль. Учет отложенной икры проводился на подконтрольных нерестилищах от зал. Шельтинга до Аяна. Средством передвижения были самолеты АН-10 и малогабаритные суда (катера и рыболовные боты). Непосредственно на нерестилищах работали с весельных лодок. На каждом участке для обнаружения и оконтуривания обыкренных площадей тралили треугольной ручной драгой.
На наиболее типичных по мощности обыкрения участках нерестилищ выбирали контрольные площадки 20×20 см, с которых брали и фиксировали в формалине пробы облепленного икрой субстрата.
При камеральной обработке материалов в каждой пробе путем навесок определяли количество икринок, а затем по этим данным подсчитывали всю икру, отложенную сельдью на учтенной площади нерестилищ.
Зная плодовитость сельди в конкретном году, высчитывали количество отнерестившихся самок, по соотношению самцов и самок определяли величину нерестового стада.
Суть дальнейших расчетов состояла в следующем: годовой коэффициент естественной смертности охотской сельди по П.В. Тюрину (1963) оказался равным 35%, при этом промысловое изъятие могло быть порядка 25-30%.
Охотская сельдь созревает к 4 годам, незначительная часть достигает половой зрелости в 2-3 года. Определив соотношение половозрелой и неполовозрелой рыбы в каждой возрастной группе в нагульном стаде или перед зимовкой, можно посчитать весь промысловый запас по формуле 42.
Количество рыбы в нагульный период (Тюрнин Б.В., 1965) (Уравнение 42).
 (42)
(42)
где К – количество рыбы в нагульный период;
Пн – % неполовозрелой рыбы перед зимовкой;
Кв – количество рыбы, выловленной при нересте;
Ко – количество отнерестившейся рыбы, учтенной по отложенной икре;
nn – % половозрелой рыбы перед зимовкой.
Учет пелагической икры (белого амура), согласно устному сообщению проводился Фарышевым Н.И. в русле Амударья в семидесятые годы прошлого столетия.
Станции облова выбирались в потоке основных струй, желательно их делать по всему русловому сечению.
Ловушки длиной 2,5 м с площадью входного отверстия – 0,25 м2 крепились фалом к лодке, поставленной на якорь.
Продолжительность экспозиции составляла 5 мин, для сравнения брались данные за 4 часа, количество воды в м3/сек, прошедшее через русло, предоставляла Гидрометеостанция.
Коэффициент уловистости ловушек определялся специально и был принят равным 0,7.
Затем среднее количество икры в сутко-станцию перемножалось на объем воды, прошедшей через русло.
Оценка биомассы минтая в Беринговом море по пелагической икре и личинкам с учетом смертности производилась Булатовым О.А. (1982) в восьмидесятых годах прошлого столетия с использованием следующих уравнений (43.1, 43.2, 43.3):
 , (43.1)
, (43.1)
N♀♀= 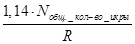 , (43.2)
, (43.2)
N♀♂=N♀♀ ·  +N♀♀, (43.3)
+N♀♀, (43.3)
где N общ – общее количество икры;
γ – уровень выживания от N0 до N лич;
N1, N2, N3 – численность икры на разных стадиях;
R – индивидуальная абсолютная плодовитость;
Y/X – соотношение самцов (♂) и самок (♀).
По данным учета икры и личинок можно судить об относительной мощности поколений, в меньшей степени эти данные могут быть использованы для абсолютных расчетов. Чаще всего по учтенной икре и личинкам можно составить суждение о тенденциях изменения численности поколений и использовать эти данные для изучения причин, обуславливающих выживание эмбрионов и личинок на ранних стадиях их развития.
Установлено, что наиболее точные результаты получают при учете икры на стадии дробления, позже количество живой икры отражает влияние условий среды на ее выживание.
Сопоставление количества живых икринок с количественными параметрами основных факторов среды обитания помогает получить в полевых условиях данные о требованиях организма к факторам среды и установить оптимальные условия для выживания эмбрионов.
Количественный учет икринок и личинок представляет известные трудности, при этом сеголетки считаются менее активными, чем последующие возрастные группы, и облавливаются равномернее.
Прежде чем начинать количественный учет молоди, необходимо установить ареал распространения ее, убедиться в том, что он относительно постоянен. Затем необходимо выявить распределение молоди по сезонам в соответствии с глубиной, соленостью, распределением кормовых организмов и другими факторами.
В исследовательской практике на внутренних водоемах для определения численности разновозрастных рыб наиболее широко применяется метод площадей, принцип которого достаточно прост и заключается в том, что количество рыбы, пойманной на определенной площади или в определенном объеме воды, относится ко всей площади (объему воды) водоема или его части, занятой скоплением рыбы.
Расчет ведется по формуле (44):
 , (44)
, (44)
где Р – площадь района учета;
m – улов в единицу времени;
Р 1 – площадь, обловленная орудием лова;
КК 1 – коэффициент уловистости, или количество пойманных рыб к количеству рыб, находящихся в зоне облова.
Величина контрольного улова определяется весьма точно, так как вылов учитывается по каждому виду рыб поштучно.
Объем процеженной тралом воды и площадь облова рассчитываются по величине раскрытия трала и расстоянию его прохождения в толще воды. Погрешности в определении облавливаемой площади зависят от выбора галсов, от точности измерения скорости и направления движения судна с тралом относительно дна. Обычно эта погрешность составляет 5-10%.
Наиболее сложно определить коэффициенты уловистости трала (К и К 1).
Как показывают наблюдения, уловистость любого орудия лова зависит от факторов биотического и абиотического характера. К первым следует отнести состояние самих рыб: их подвижность, активность, реакцию на орудие лова, стайность, особенности распределения и условия нагула, ко вторым – неровности дна, конструкции орудий лова, изменения температуры, погодные условия.
Таким образом, практически невозможно установить единый коэффициент уловистости даже одного трала при различных экологических ситуациях. При этом надо помнить, что орудия лова имеют четко выраженную селективность относительно размеров и вида рыбы.
Коэффициент вертикальной уловистости в значительной степени определяется горизонтом траления. Донный трал в 2-3 раза уловистее пелагического, что, вероятно, обусловлено тем, что у дна рыбе труднее его избежать. Для учета вертикального распределения иногда вводят поправку на соответствие высоты изучаемого скопления (h скопления), что регистрируется по показаниям эхолота, к вертикальному раскрытию трала (h траления). В этом случае коэффициент поправки (К) равен (45):
 , (45)
, (45)
В рыбохозяйственной практике, особенно на внутренних относительно мелководных водоемах, часто не производят деления уловистости на вертикальную и горизонтальную составляющие, вводя общий показатель уловистости.
Вариации уловистости одного и того же трала даже в сходных условиях весьма значительны. Например, при траловом лове леща на Цимлянском водохранилище К колебался от 0,28 до 0,104.По другим данным уловистость трала в этом же водоеме имела размах 0,22-0,47, а при облове ряпушки в озере Плещеево в ночное время варьировала в пределах 0,2-0,75 (в среднем 0,4). Днем рыба совершенно избегала трала.
Исходя из многолетней практики, погрешность уловистости трала оценивается в 60%. Она в основном определяет точность расчета плотности рыб по траловым уловам.
Еще более существенную погрешность получают при экстраполяции полученных данных по плотности на весь водоем, где распределение рыбы не равномерно. Для уменьшения дисперсии оценки применяют метод изолиний, заключающийся в разделении изучаемой акватории на участки с одинаковой плотностью рыбы. При этом, чем больше частота выборки плотности, тем меньше дисперсия и соответственно погрешность оценки.
На внутренних водоемах не все участки могут быть одинаково интенсивно облавливаемы тралом. Более того, при высокой подвижности рыб необходима синхронная съемка всей изучаемой акватории, что не всегда осуществимо. В этих случаях приходится по 10-30 разрезам, где определена плотность, рассчитывать численность рыб на акватории, в десятки тысяч раз большей.
Так, например, при определении численности леща в Каховском водохранилище траления были проведены по 31 разрезу с разрывом в 10 км, а эти же расчеты для озера Ильмень основаны всего на 23 тралениях. Поскольку внутренние водоемы характеризуются большим разнообразием биотопов, где плотность рыб варьирует в десятки и сотни раз, погрешность в оценке общей численности рыб может достигать 250-300%.
Однако во многих ситуациях при возможности охвата исследуемой акватории достаточно плотной сеткой станции и экспериментальном определении уловистости конкретного трала для основных видов рыб расчетная оценка величины стада может быть вполне удовлетворительной. Более того, в ряде случаев, например, при учете донных рыб, подобный метод оценки является наиболее удобным и точным.
С появлением рыбопоисковой техники метод площадей резко повысил объективность. Уже накоплен большой опыт применения гидроакустических методов оценки рыбных запасов в морях. В последние годы эти методы широко внедряются во внутренних водоемах. Их применение позволяет в расчетах полностью исключить ошибку уловистости орудий лова, анализировать весь столб воды (высоту исследуемого скопления рыбы), обследовать любые участки водоема, где глубины соответствуют режиму работы рыбопоисковых приборов, обеспечив нужную быстроту съема информации с большой акватории и внедрив автоматизацию расшифровки показаний приборов, уменьшить субъективную ошибку подсчета эхомишений.
Численность рыб гидроакустическим методом находят следующим образом: выполняют детальную эхометрическую съемку исследуемого района по системе определенных разрезов, разбивают каждый из них на более дробные участки и находят плотность рыб на каждом участке, зная число «высвеченных» рыб и объем зоны действия прибора. Далее, проведя интерполяцию данных на «непросвеченную» область и просуммировав все, находят численность рыб в скоплении или в водоеме.
Расшифровка показаний приборов проводится либо визуально, либо автоматически с помощью эхосчетчиков или эхоинтеграторов, которые суммируют показания прибора в заданный отрезок времени.
Визуальная расшифровка в настоящее время проводится в основном не для исследования численности, а при изучении распределения рыб, но она вполне применима и для количественной оценки разреженных скоплений, если в зоне действия прибора на одном горизонте находится не более одной рыбы.
Численность рыб гидроакустическим методом рассчитывается по следующей формуле Ю.Т. Сечина (46):
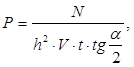 (46)
(46)
где Р – плотность скопления, шт./м3;
N, t – число отметок на эхолотной ленте за время t, часах;
H – глубина водоема, м;
a – угол излучения вибратора, берется по паспорту эхолота.
Так при определении численности осетровых в р. Волга (ширина зоны действия гидролокатора «Лещ» составляет 5-6 м на глубине 15-20 м) подсчет количества эхомишений оказался наиболее точным (погрешность составила менее 10%) только в том случае, если между ближайшими рыбами по горизонтали расстояние было не мене 5-6 м. Нередко плотность рыб такова, что сигналы от отдельных особей визуально различить трудно, погрешность учета эхомишений в таком случае достигает 50%. Так при определении численности осетровых под плотиной Волжской ГЭС им. XXII съезда КПСС в 1981 г. плотность варьировала в пределах 0,02-0,1 шт./м2. Это обусловило сравнительно небольшую точность при гидроакустической оценке мощности подплотинного скопления осетровых, которая была оценена как 57±24 тыс. производителей. Однако эти данные уже можно было использовать в целях рыбохозяйственного прогнозирования.
Для стайных рыб выделить отдельные эхосигналы визуально нельзя. В этом случае численность рыб рассчитывается по количеству и размерам стай на эхограммах, а плотность рыбы в стае находится по контрольным уловам, то есть неизбежно вводится коэффициент уловистости трала. Более точно плотность стай определяется с помощью кино- и фотосъемки и погрешность в расчетах здесь оценивается в 10-15%. Но фотосъемка возможна только в высокопрозрачных водах. Идентификация вида рыб осуществляется по анализу контрольных уловов, проводимых в местах эхосъемок. Ясно, что для разновидовых скоплений точность определения соотношения рыб различного вида будет обусловлена возможностями облова. При этом возникает погрешность, связанная с неодинаковой уловистостью даже одного и того же орудия лова для различных видов. Опыт показывает, что некоторые виды рыб, например судак в дневное время в трал попадают значительно реже, чем ночью. Это необходимо учитывать при оценке численности рыб в смешанных скоплениях по видам. Наименьшая разница уловистости трала для различных видов рыб отмечается в темное время суток.
И.Л. Калихманом (1982) показано, что систематическая ошибка в определении плотности пелагических скоплений рыб составляет 33%. Однако при выполнении требований к повышению точности, ошибка в определении плотности может быть уменьшена до 20%, при этом погрешность в расчетах гидроакустическим методом может быть гораздо меньшей.
Гидроакустические методы, повышая класс точности определения величины рыбных запасов, имеют область ограничений, которые обусловлены особенностями распределения рыб. Значительная масса рыб, преимущественно бентосоядных, сосредотачивается в придонных слоях воды. Минимальное расстояние от дна, при котором рыба обнаруживается с помощью рыбопоисковых приборов, определяется длительностью импульса и шириной «высвечивания». Например, гидролокатор «Лещ» регистрирует только рыб, которые находятся в 0,3-0,5 м выше дна. Для регистрации рыб вблизи дна требуется существенное увеличение разрешающей способности рыбопоисковой аппаратуры.
Большие возможности в регистрации придонных скоплений дают знания особенностей их суточных вертикальных миграций, позволяя приурочить проведение гидроакустических съемок к периоду массового подъема рыб от дна в толщу воды.
Существенные ограничения применения рыбопоисковых приборов связаны также с тем, что во внутренних водоемах значительная масса пелагических рыб сосредотачивается в приповерхностном слое воды, находящемся в «мертвой» зоне гидролокатора. Это ограничение полностью снимается при буксировании приемно-передающих устройств на глубине и при просвечивании толщи воды не от поверхности, а, наоборот, от дна к поверхности.
При работе на малых глубинах неизбежны погрешности в учете рыб, связанные с их отпугиванием от идущего за эхолотом судна и уходом из зоны действия прибора. В этом случае помогут существенно снизить погрешность буксируемые приемно-передающие устройства, так как плотность рыб будет рассчитываться не под судном, где количество рыбы падает в десятки раз, а в стороне от него.
Диапазон применения гидроакустических методов значительно расширяется при их комбинировании с биотелеметрией, контрольными обловами и средствами наблюдения с подводных аппаратов.
Методы определения численности движущихся рыб
Для учета численности стада проходных и полупроходных рыб существенное значение имеют способы оценки количества рыбы, прошедшей за определенный отрезок времени через поперечное сечение участка реки, лежащего ниже нерестилища. Впервые в России этот метод оценки численности мигрирующего стада был предложен Ф.И. Барановым (1933, 1960) для учета численности мигрирующей воблы. Для учета пользовались закидным неводом. Ф.И. Баранов принимал, что общее количество рыбы, прошедшее через облавливаемый участок реки, равно (47):
N=n·U·S·T, (47)
где N – общее число рыб, прошедших через облавливаемый участок реки;
n – число рыб на единицу площади;
U – скорость хода рыбы;
S – ширина реки;
Т – время между двумя заметами.
Однако таким способом лишь в редких случаях удается установить абсолютную численность прошедших рыб. Это связано с неравномерностью распределения движущихся рыб поперек русла, разной уловистостью невода в отношении рыб, встречающихся с неводом в разные фазы притонения. Этот метод более применим для учета относительной численности мигрирующих рыб.
Точность этого метода проверялась на примере расчета абсолютной численности нерестовой популяции осетровых, мигрирующих из Каспия в реку Волга. Отлов производителей производился на экспериментальной тоне «Мужичья», расположенной в 65 км выше г. Астрахани и действующей весь период открытой воды. Определялось количество рыб на всем сечении реки во время контрольных неводных обловов, и полученная величина экстраполировалась на период, когда облова не было.
Наибольшая погрешность расчета возникает при определении количества рыб, проходящих через все сечение реки за весь день работы невода (уравнение 48).
 (48)
(48)
где N – количество рыб;
K – коэффициент уловистости;
q – улов;
S 1 – облавливаемая площадь;
S 2 – необлавливаемая площадь.
Наиболее вариабельным оказался коэффициент уловистости. Коэффициент уловистости (К) для закидных неводов на промысле осетровых составляет 0,18-0,75 (в среднем 0,38) или 0,17-0,56.
Следующий источник ошибок связан с определением общей облавливаемой неводом площади сечения реки. Погрешность определения площади облова оказалась равной 26%, а после пересчета количества рыб на все сечение реки – 30% при допущении, что мигрирующие рыбы распределяются по нему равномерно. Однако, телеметрические наблюдения показывают, что осетровые преимущественно мигрируют вблизи русловых склонов, что вносит в расчет дополнительную погрешность еще на 30%.
На втором этапе расчетов численности рыб по контрольным обловам величина экстраполируется на весь период нерестового хода с учетом времени действия невода, суточной и сезонной ритмики миграции. Первый показатель легко поддается строгому учету и его погрешность невелика. При большей повторности измерений (экспериментальная тоня работает весь период открытой воды) сравнительно точно и характеризует особенности сезонного хода рыбы.
Однако анализировать суточную ритмику активности рыб по уловам трудно. По одним данным дневные уловы рыб не отличаются от ночных, по другим наибольшая интенсивность хода приходится на ночное время, по данным телеметрических наблюдений в ночное время двигательная активность осетров в среднем на 20% выше и в этот период лова больше их количество успешно минует тоневый участок. За счет неравномерности суточной активности рыбы погрешность в оценке численности на створе тоневого участка за сутки равна 20%. В итоге общая погрешность в расчете численности мигрирующих рыб методом контрольных обловов за весь сезон составляет не менее 50%.
Наиболее широко применяется количественный учет проходных дальневосточных лососей во время их нерестовой миграции. Этим способом удается в течение ряда лет учитывать численность отдельных локальных стад, заходящих для нереста в определенные реки. Многолетние материалы накоплены в отношении стад Камчатки, Сахалина, Приморья, Охотского побережья. Обычно просчет проводится визуально путем учета рыбы, периодически пропускаемой через специальные окошки в перегораживающих реку сплошных заграждениях.
На Аляске проходящих к нерестилищам рыб просчитывают путем периодической киносъемки рыб, пересекающих выложенные на дне белые панели, и путем экстраполяции пересчитывают на сутки (Mathisen, 1962а).
Затем, используя соответствующий эмпирический переводной коэффициент, подсчитывают абсолютное количество рыб, прошедших за определенный промежуток времени данное сечение реки. В 1955 г. в день снимали в среднем 1440 кадров.
Разница между данными, полученными путем непосредственного просчета и путем пересчета рыб с кинокадров, оказалась очень небольшой, она составляла в среднем 1,91%.
В.Р. Протасов и Ю.А. Митрохин (1960) сконструировали прибор, позволяющий просчитывать количество и измерять длину рыб, проходящих через определенный участок (рис. 52).
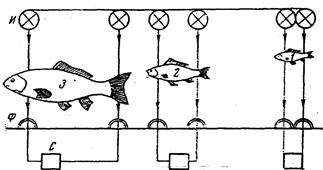
Рис 51. Схема размещения фотоэлементов и источников света, позволяющая регистрировать рыб по размерам (по Протасову и Митрохину, 1960): Ф – фотоэлементы; И – источник света; С – счетчики; 1, 2, 3 – учитываемые размерные группы
Рыбы, проходящие через определенный отсек, прерывают свет, тем самым отмечаются как длина рыбы, так и число рыб, прошедших через определенное отверстие. Недостаток метода заключается в том, что он не позволяет учитывать видовую специфику проходящих рыб; кроме того, в некоторых случаях две, проходящие непосредственно одна за другой рыбы могут сливаться. Основная сложность заключается в обеспечении раздельного учета при совместной миграции нескольких видов рыб (Протасов и Неймарк, 1964).
Точность учета в таких условиях в основном зависит от длительности работы наблюдателя или качества счетных устройств. Ошибка может быть сведена к минимуму, если учет производится круглосуточно на протяжении всего нерестового периода. Однако при этом необходимо учитывать, что не все производители заходят в ловушки, часть рыбы отпугивается и скатывается назад или длительное время передвигается перед ловушками. Этого недостатка легко избежать, если использовать электронные счетчики, устанавливаемые прямо на дне реки без перекрытия ее заграждениями. Например, разработано счетное устройство, в основе которого лежит прокладка по дну поперек реки кабелей с интервалом, соответствующим размерам учитываемых рыб. Проплывающие над кабелями рыбы вызывают в них изменение электропроводности, что отмечается регистрирующим прибором высокой точности (Kristinsson, et al., 1978).
Однако экстраполяция точных данных, полученных для нескольких малых рек на весь нерестовый район, может быть отягощена большими ошибками из-за сильной изменчивости роли этих рек в воспроизводстве рыбных запасов.
Так же, как учет рыб при нерестовой миграции, в ряде случаев возможен и учет скатывающейся молоди проходных рыб, особенно молоди лососей. Редко удается осуществить абсолютный учет всей скатывающейся молоди. Обычно же учет покатной молоди ловушками дает лишь относительные величины (Таранец, 1939) (49).
 , (49)
, (49)
где t – время экспозиции;
N – поймано молоди за ночь;
M – количество обловов;
Т – покатный период;
К – коэффициент неравномерности распределения;
S – п






