Базальные ганглии представляют собой комплекс нервных клеток, которые впервые описаны английским анатомом, физиологом, врачом Томасом Уиллисом Thomas Willis, 1621-1675) и сравнительно давно получили название базальных или подкорковых ядер (nucl. basales), так как располагаются ближе к основанию мозга. Это старое топологическое название сегодня не может в полной мере отражать современные представления о морфофункциональной организации, локализации и функции, входящих в состав базальных ганглиев анатомических и функционально близких к ним структур мозга. Двигательные ядра базальных ганглиев подразделяют на несколько анатомически и функционально различных групп. Базальные ганглии с анатомических позиций включают хвостатое ядро (nucl. caudatus), скорлупу (putamen), бледный шар (globus pallidus, имеющий внутренний и наружный сегменты) и ограду (claustrum). Указанные 4- е ядра анатомически составляют полосатое тело (corpus striatum). Термин striatum (стриатум) приобрел другое содержание, так как в его состав вошли хвостатое ядро (nucl. caudatus) и бледный шар (globus pallidus, имеющий внутренний и наружный сегменты). Скорлупа и бледный шар объединяются в чечевицеобразное ядро (nucl. lentiformis).Функционально базальные ганглии связаны с ядрами таламуса с передним вентральным ядром (nucl. ventralis anterior) и латеральным вентральным ядром (nucl. ventralis lateralis). Кроме этого базальные ядра имеют важные функциональные связи с субталамическим ядром (nucl. subthalamicus) и черной субстанцией, в состав которой входят компактная и сетчатая части (sub. nigra, pars compacta et retina) (рис...).
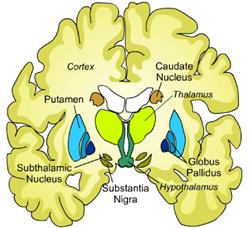
Рис.... Поперечный срез головного мозга
Базальные ганглии являются парными образованиями и распространяются в толще лобных долей больших полушарий по обе стороны от таламуса над стволом (уровень промежуточного мозга). При этом хвостатое ядро, состоящее из головки, тела и хвоста из лобных долей в виде буквы "С" пронизывает теменные и затылочные доли, заворачивается снова вперед и в полюсе височной доли образует миндалевидное ядро. Каудальная стенка головки хвостатого ядра граничит с таламусом, отделяясь от него полоской белого вещества (stria terminalis). (рис. см два рис ниже их объединить).. С функцией базальных ганглиев тесно связана черная субстанция, ее нейрохимические пути проецируются на нейроны стриатума. Но по местоположению она находится в стволовой части среднего мозга. C функциональных позиций к базальным ганглиям, а значит и к участию в управлении движениями следует отнести и участки коры большого мозга: префронтальную ассоциативную, премоторную и дополнительную моторную, первичную моторную, сенсомоторную и фронтальную глазную области (рис....).Аксоны кортикоспинального и кортикобульбарного путей от пирамидных клеток коры большого мозга, минуя базальные ганглии образуют полоску белого вещества - внутреннюю капсулу. В состав внутренней капсулы входят и другие нисходящие и восходящие к коре больших полушарий проводниковые системы. Неврологи перечисленный комплексанатомических образований называют экстрапирамидной системой.
Базальные ганглии как составная часть экстрапирамидной системы прежде всего участвуют в организации и регуляции моторики.
Интегратором и распределителем перехода одного вида движения в другое, а также триггерным органом запуска программы совершения действий является стриатум. Стриатум функционирует как «ворота» для входа глутаматергических возбуждающих химических сигналов из коры. Кроме этого стриатум является и коллектором афферентных входов, идущих от неспецифических ядер таламуса и дофаминэргических путей от черной субстанции. Стриатум вовлечен и в абстрактные программы теменной и затылочной частей мозга благодаря связям с лобными долями. Благодаря этому в моторных программах стриатума решается вопрос, «какое движение было бы целесообразнее сделать из тех, которые заложены в программе».
Скорлупа (как часть стриатума) получает входы от моторных и сенсорных областей, соседствующих с первичной моторной корой и выполняет специфическую функцию по организации пищевого поведения, поскольку ее активность прямо связана с двигательной системой.
Хвостатое ядро имеет прямые связи с фронтальной корой и поэтому оно вовлечено в когнитивные процессы высокого порядка, участвует в реализации эмоций, вегетативных реакций.
♦ Базальные ганглии участвуют в выполнении сложных двигательных программ совместно с кортикоспинальной системой. Передача химических сигналов по комплексу базальных ганглиев имеет четкий вектор: от премоторных зон коры с возвратом сигналов вновь в кору, но в первичную моторную область.
Иными словами, работает принцип отрицательной обратной связи ограничения числа выходящих двигательных команд. Таким образом между корой больших полушарий и базальными ганглиями сформировался кольцевой контур для управления приобретенными движениями (рис....).
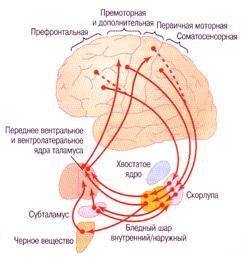
Рис.... Кольцевой контур премоторная кора - базальные ганглии - моторная кора для управления движениями (из А. Гайтон).
Основной функциональной задачей моторной функции базальных ганглиев является модуляция (корректировка) замысла таких параметров движения, как сила, амплитуда и направление.
Причем, сам замысел движения возникает в премоторной коре (см. рисунок). Вспомните! именно из этого отдела базальные ганглии (стриатум) получает двигательную команду. Поэтому благодаря такой связи базальные ганглии работают как механизм выбора включения двигательных поведенческих программ еще до их совершения. На самом деле программы в базальных ганглиях уже заложены, но базальные ганглии "принимают стратегическое решение" к какому действию прибегнуть в конкретный момент времени: осмотреться, вслушаться или произвести какое-либо действие. Коррекция движения (замышленного в премоторной коре) осуществится в структурах базальных ганглиев еще до того как движение исполнится первичной корой. Например, несмотря на изменившуюся конкретную обстановку, ряд сложных дискретных движений, будут выполнены в определенном правильном порядке, путем активации обычных движений и исключении излишних, например, при ежедневном одевании одежды, или выполнении последовательных действий при игре на музыкальном инструменте. Роль базальных ганглиев при этом сводится к двум системам обучения выше указанным действиям: неосознанном и осознанном.Причем описанные и другие действия при участии базальных ганглиев совершаются с определенной индивидуальной последовательностью, пластикой, ритмом и гармонией всех частей тела конкретного человека.
Непосредственную модуляцию (подправку) сложных двигательных актов, инициируемых корой больших полушарий базальные ганглии осуществляют с помощью 2- х нервных путей: прямого и непрямого. КОЛЯ сюда кольцо-два контура
Начало двух путей идентично: от премоторной коры - к стриатуму. Далее аксоны от нейронов стриатума разделяются на 2- а направления. Первое направление (оно получило название прямого пути) связывает стриатум с внутренним сегментом бледного шара и сетчатой частью черной субстанции. Нейроны бледного шара (внутренний сегмент) обладают высокой спонтанной активностью и приходящие от нейронов стриатума аксоны уровень этой активности снижают, поскольку их окончания образуют на них ГАМКергические тормозные синапсы. Второе направление (непрямой путь) также связывает стриатум с внутренним сегментом бледного шара и сетчатой частью черной субстанции, но как вытекает из названия двигательные команды из стриатума приходят туда не напрямую, а сначала следуют в наружный сегмент бледного шара, затем через субталамическое ядро возвращаются в бледный шар.
Прямой путь. Суть функции этого пути состоит в том, что базальные ядра облегчают запускаемые корой движения и подавляют лишние сопутствующие движения. Инициатива (замысел) сложного двигательного акта для прямого пути возникает в премоторной коре ( и в соматосенсорных областях). Пучки нервных волокон от премоторных областей лобной коры распределяются на нейронах скорлупы в виде отдельных полосок (отсюда и произошло название -стриатум - полосатое тело). Только моторные нервные клетки премоторной области имеют прямое отношение к регуляции движений, так как отправляют глутаматергические, возбуждающие проекции в стриатум, минуя хвостатое ядро. Нейроны стриатума посылают аксоны к внутреннему сегменту бледного шара и сетчатой части черной субстанции, на которых они образуют ГАМК- ергические синапсы (первое последовательное тормозное переключение). В свою очередь нейроны внутреннего сегмента бледного шара и сетчатой части черной субстанции с помощью аксонов образуют тормозные ГАМК-ергические синапсы (второе последовательное ГАМК-ергическое тормозное переключение) на переднем и латеральном вентральных ядрах таламуса. Нейроны бледного шара тонически активны, посылая до 50 потенциалов действия в секунду на нейроны ядер таламуса в кору что приводит к постоянному торможению прохождения возбуждения от таламуса к коре. От нейронов переднего и латерального вентральных ядер таламуса к первичной моторной коре следуют главные эфферентные возбуждающие глутаматергические волокна (их пути совпадают с проекциями от мозжечка). Таким образом, при торможении нейронов внутреннего сегмента бледного шара и сетчатой части черной субстанции возбуждающее влияние таламуса на кору усиливается - что и способствует облегчению запускаемого корой движения. Напротив, при возбуждении нейронов внутреннего сегмента бледного шара и сетчатой части черной субстанции возбуждающее влияние таламуса на кору подавляется - что приводит к торможению излишнего движения. Прямого выхода на спинной мозг базальные ядра не имеют. Прямой путь от стриатума к внутреннему сегменту бледного шара и сетчатой части черной субстанции образован тормозными ГАМКергическими волокнами. Функция тормозной ГАМК- ергической передачи как раз и заключается в усилении возбуждающего влияния ядер таламуса на отделы первичной моторной коры, которые и будут исполнять необходимое в конкретной ситуации действие по активации скорректированных движений. В итоге нервные волокна прямого пути от ядер таламуса возвращаются к первичной моторной коре больших полушарий (исполнитель сложного движения) и к областям премоторной коры и дополнительной коры.
СЮДА рис прямого пути
Посредством нейронов, формирующих прямой круг базальные ядра вовлечены в регуляцию двигательных программных команд, исходящих из премоторных областей коры: если стриатум тормозит активность бледного шара и черной субстанции возбуждающее действие моторных ядер таламуса на нейроны первичной области коры больших полушарий повышается - движения совершаются в полном объеме. В случае возбуждения нейронов внутреннего сегмента бледного шара и сетчатой части черной субстанции активирующее воздействие таламических ядер на корковые нейроны угнетается и излишние движения тормозятся (исключаются из программы движения). В итоге базальные ганглии с помощью функции прямого замкнутого круга выбирают из предложенного корой ряда двигательных программ адекватную для реального осуществления движений в данный момент времени и облегчают ее реализацию. Функциональная значимость прямого пути состоит в облегчении запускаемых корой движений то есть активации необходимых (функционально обоснованных) компонентов биологических, бытовых, профессиональных, спортивных сложных движений, например, резания ножницами, правильного написания букв, забивания гвоздей молотком, бросков баскетбольного мяча в кольцо, ведения и финтов мяча в футболе, движений лопатой при копании земли, вокализации, сознательных управляемых движений глазами, практически любых точных, выполняемых автоматически движений ит.д. Смысл двух последовательных тормозных переключений в прямом пути нивелирование одного другим и в итоге прямой путь является возбуждающим.
Непрямой путь. Функция его заключается в подавлении возбуждающего влияния таламуса на другие отделы моторной коры. Начало непрямого пути сперва также исходит из премоторной и соматосенсорных областей коры, откуда в скорлупу, минуя хвостатое ядро, следуют аксоны, выделяющие в своих окончаниях глутамат. Из нейронов скорлупы аксоны следуют к наружной части бледного шара, где и контактируют с его нейронами с помощью тормозных ГАМК- ергических синапсов (первое тормозное переключение). Затем путь от наружного сегмента бледного шара продолжается к нейронам субталамического ядра, где заканчивается также тормозными ГАМК- ергическими синапсами (второе тормозное переключение). От нейронов субталамического ядра следуют возбуждающие глутаматергические волокна, причем часть их возвращается к наружному бледному шару, другие направляются к внутреннему сегменту бледного шара и сетчатой части черной субстанции. От нейронов сетчатой части черной субстанции аксоны этих нейронов передают тормозные ГАМК- ергические сигналы на нейроны таламуса (третье тормозное переключение). Ядра таламуса далее активируют первичную моторную кору с помощью глутамата. Таким образом, непрямой путь оказывается более сложным, не только конструктивно, но и функционально: если активация прямого пути от стриатума усиливает возбуждение моторной коры, то наличие у непрямого пути - трех тормозных переключений, способствует подавлению возбуждающего влияния таламуса на другие отделы моторной коры и относит непрямой путь к тормозному пути.
Активность прямого и непрямого путей регулирует компактная часть черной субстанции, посылающая к стриатуму дофаминергические волокна. Эти волокна возбуждают прямой эфферентный путь стриатума (через нейроны с D1-рецепторами) и тормозят - непрямой (через нейроны с D2-рецепторами). Полосатое тело посылает также тормозные ГАМКергические волокна к компактной части черной субстанции, образуя тем самым с ней отрицательную обратную связь. Наконец, активность стриатума модулируют его холинергические вставочные нейроны - антагонисты дофаминергических нейронов.
СЮДА рис непрямого пути
Функциональная значимость непрямого пути состоит в уменьшении и подавлении излишних компонентов и действий сопутствующих сложным движениям, например, ограничение размашистости движений рук, ног, других частей тела, быстрых подергиваний лица, кистей, в написании букв и цифр и т. д.
♦♦ Важно также в связи с вышеизложенным: Базальные ганглии не имеют прямых (непосредственных) эфферентных связей с мотонейронами спинного мозга. Базальные ганглии управляют альфа- и гамма- мотонейронами мышц-разгибателей через красные ядра, мышцами-сгибателями - через вестибулярные ядра, а также через ретикулярные ядра продолговатого мозга и моста.






