Нейроны ретикулярной формации, принимающие участие в регуляции движений занимают центральную часть продолговатого мозга и моста в виде скоплений, которые группируются в ядра. На уровне продолговатого мозга - это гигантоклеточное, мелкоклеточное и латеральное ретикулярные ядра, которые локализованы медиально относительно средней линии (см. рис....). В мостовой части ствола нейроны ретикулярной формации составляют каудальное, вентрикулярное и оральное ретикулярные ядра (см. рис....).
Заметьте! Двигательные ядра ретикулярной формации распределены в стволе таким образом, что на них поступает информацию от всех сенсорных (восходящих) и моторных (нисходящих) систем.
Впервые механизм воздействия ретикулярной формации на перераспределение тонуса мышц тела обосновал Р.Гранит (R.Granit). Нейроны ретикулярной формации моста (РФМ) и нейроны ретикулярной формации продолговатого мозга (РФПМ) в сущности являются функциональными антагонистами: нейроны РФМ возбуждают антигравитационные мышцы, а нейроны РФПМ расслабляют мышцы-разгибатели.
Нервные клетки ретикулярной формации продолговатого мозга связаны в нисходящем направлении с сегментарным аппаратом спинного мозга и формируют латеральный ретикулоспинальный путь продолговатого мозга. Аксоны этого пути заканчиваются на α- мотонейронах сгибателях передних рогов. Возбуждение нейронов ретикулярной формации продолговатого мозга активирует эти α-мотонейроны - усиливает тонус мышц-сгибателей и тормозит (расслабляет) антигравитационные мышцы-разгибатели. На нейронах ретикулярной формации продолговатого мозга заканчиваются аксоны: от пирамидного тракта (кортикоспинальный путь), от красного ядра (руброспинальный путь), приходящие по этим аксонам сигналы возбуждения на α-мотонейроны также повышают тонус мышц-сгибателей.
Нейроны ретикулярной формации моста посредством аксонов образуют медиальный ретикулоспинальный путь моста для связи с α-мотонейронами спинного мозга. Нейроны РФМ постоянно через альфа-мотонейроны спинного мозга направляют на осевую мускулатуру тела (разгибателям спины, шеи и нижних конечностей) потенциалы действия, которые сохраняют вертикальное положение тела путем сокращения антигравитационных мышц. Эти мышцы противодействуют силам земного притяжения и создают базисный осевой разгибательный мышечный тонус. Данный тонус позволяет в вертикальном положении совершат весь комплекс необходимых движений, присущих человеку. Нервные сигналы от РФМ возбуждают α-мотонейроны - усиливают тонус мышц-разгибателей и вызывают торможение α-мотонейронов - снижение тонуса мышц-сгибателей. Таким образом, ретикулярная формация моста является функциональным антагонистом нейронов ретикулярной формации продолговатого мозга. Нейроны ретикулярной формации моста находятся под сильным содружественным влиянием вестибулярных ядер и глубоких ядер мозжечка, от которых они получают возбуждающие стимулы. Большинство аксонов медиального ретикулоспинального пути заканчиваются на грудных сегментах спинного мозга, которые управляют мышцами распрямляющими позвоночник и мышцами - разгибателями конечностей. Таким образом, РФМ является мощным "стимулятором" выпрямительных рефлексов, так как активирует антигравитационные мышцы нижних конечностей,туловища и шеи.
Контроль работы мышц-разгибателей нижних конечностей и особенно осевой мускулатуры тела ретикулярной формацией ствола поддерживают вестибулярные ядра. Адекватными раздражителями вестибулярных ядер являются статическое положение головы в пространстве и изменения его, а также ускорения тела. Особенностью их управления указанными мышцами является поддержание равновесия тела в гравитационном поле Земли при разнообразных движениях, внешних возмущениях тела, например, толчках. Нейроны вестибулярных ядер возбуждают альфа-мотонейроны разгибателей и одновременно по механизму реципрокной иннервации тормозят альфа-мотонейроны сгибателей. Поэтому при раздражении вестибулярного аппарата тонус верхних и нижних конечностей меняется так, что равновесие тела сохраняется. Вестибулярные ядра образуют два вестибулоспинальных тракта: латеральный и медиальный, которые связывают вестибулярные ядра с α-мотонейронами, аксоны которых заканчиваются на мышцах-разгибателях крестцовых сегментов спинного мозга. Таким образом, вестибулярные ядра поддерживают возбуждающие влияния нейронов ретикулярной формации моста на антигравитационные мышцы туловища и нижних конечностей и увеличивают тонус мышц-разгибателей. Вестибулярное ядро Дейтерса находится под контролем мозжечка и связано с ним вестибулоцеребеллярным путем. Через этот путь мозжечок получает всю информацию от вестибулярного аппарата, где она перерабатывается, главным образом, в архицеребеллуме. Затем от него сигналы следуют в ядро шатра мозжечка, а затем вновь на вестибулярные ядра (ядро Дейтерса). Иными словами, вестибулярные ядра контролируют равновесие тела человека, положение шеи относительно головы и совместно с мозжечком корректируют любые отклонения туловища от положения равновесия. Так мозжечок контролирует деятельность вестибулярных ядер, поэтому поражения мозжечка проявляются схожими симптомами, что и при патологии вестибулярного аппарата и вестибулярных ядер.
Красное ядро расположено в стволе в области среднего мозга на уровне четверохолмия.
Двусторонние связи нейронов красного ядра с корой головного мозга, мозжечка (точнее с пробковидными и шаровидными ядрами) свидетельствуют об участии нейронов красного ядра в контроле положения тела в пространстве, о состоянии мышечной системы и кожи. Красное ядро (его нейроны) формируют руброспинальный тракт и преимущественно активируют альфа-мотонейроны сгибателей и тормозят активность альфа-мотонейронов разгибателей, поэтому совместно с вестибулярными ядрами красное ядро регулирует бессознательные, автоматические движения, например, перемена или восстановление позы тела.
Четверохолмие составляет часть ствола мозга, причем верхние холмики относятся к зрительной сенсорной системе, а нижние холмики - к слуховой. Нейроны, входящие в состав четверохолмия обеспечивают ориентировочные зрительные и слуховые рефлексы и координируют повороты головы и туловища в сторону световых и звуковых раздражителей. Четверохолмие формирует тектоспинальный путь.
Указанные ядра ствола мозга человека и соответствующие рецепторы осуществляют с татические и статокинетические рефлексы.
Статическими рефлексы называют так потому, что с их помощью поддерживается поза и равновесие тела. Статические рефлексы наблюдаются при относительном покое тела: стоянии, лежании и сидении в различных положениях, и выражается в рефлекторном перераспределении мышечного тонуса в зависимости от положения тела в пространстве
Поза - это закрепление частей скелета при самых различных положениях при выполнении работы и в покое, например, спокойное лежание, стояние и сидение.
С помощью статических рефлексов происходит перераспределение тонуса сгибательных и разгибательных мышц таким образом, что фиксируется неподвижное положение суставов для поддержания позы и сохранения равновесия и ориентации конечностей во время движения. Среди статических рефлексов различают позные или рефлексы положения и выпрямительные - установочные рефлексы. Выпрямительные рефлексы возникают при смене одной позы на другую, например, если придать животному неестественное положение, то оно быстро и точно вернется в типичную для него позу, причем при восстановлении позы животного выпрямительные рефлексы совершаются в определенной последовательности. Прежде всего восстанавливается положение головы под влиянием сигналов от вестибулярного аппарата. Поэтому эти выпрямительные рефлексы называются еще лабиринтными. Но выпрямление положения головы животного по отношению к туловищу (например, после запрокидывания ее) не остается "незамеченным", восстановление ее положения "улавливается" проприорецепторами шейных мышц. Такие выпрямительные рефлексы, происходящие с участием проприорецепторов шеи получили название шейные. Перечисленные статические двигательные рефлексы замыкаются на уровне ствола головного мозга и не требуют участия сознания (воли) человека. Выпрямительные рефлексы могут быть вызваны и с рецепторов других частей тела (например, с экстерорецепторов подошв стоп, которые передают ощущения давления на опору и распределения веса тела между двумя ногами, кожных экстерорецепторов), а также с рецепторов сетчатки глаз и проприорецепторов мышц глаз. Афферентная сигнализация от указанных рецепторов также замыкается в стволовой части мозга. Функциональная значимость выпрямительных рефлексов заключается в том, что ведущую роль в них играет голова, где как раз и располагаются рецепторы сетчатки, сенсорные клетки лабиринта, проприорецепторы глазодвигательных мышц.
Статокинетические рефлексы служат для поддержания позы во время движений и вызываются при действии на организм прямолинейного или углового ускорения. Статокинетические рефлексы компенсируют отклонения тела при ускорении или замедлении прямолинейного движения, а также при вращениях. К статокинетическим рефлексам относят повороты головы при вращении, принятие позы при свободном падении и так называемые "лифтные" рефлексы. При вертикальных ускорениях в момент начала движения лифта вверх у человека, который стоит на площадке лифта снижается тонус мышц-разгибателей и его ноги подгибаются, при движении же лифта вниз тонус мышц-разгибателей возрастает и ноги фиксируются в положении максимального разгибания. При вращении голова, тело и глаза отклоняются в сторону противоположную движению. При этом глаза медленно отклоняются в сторону противоположную движению и быстро возвращаются в исходное положение Такое движение глаз при вращении тела называется нистагм. Этот рефлекс обеспечивает сохранение изображения внешнего мира на сетчатке глаз и тем самым зрительную ориентацию.
В клинической практике статические и статокинетические рефлексы получили название - постуральные рефлексы.
Нисходящие влияния ретикулярной формации осуществляются через ретикулоспинальный путь, оказывающий облегчающее или тормозящее влияние на сегментарный аппарат спинного мозга. Тормозящее поле соответствует гигантоклеточному ретикулярному ядру продолговатого мозга. Облегчающие зоны локализованы менее четко, они захватывают большую зону — часть гигантоклеточного ядра, ядра моста, облегчающие влияния с уровня среднего мозга, осуществляются через полисинаптические связи. Нисходящие влияния ретикулярной формации воздействуют на α- и γ-мотонейроны, оказывающие влияния на мышечные веретена и вставочные нейроны.
Показано, что большинство волокон ретикулоспинального пути оканчиваются не ниже грудных сегментов и только вестибулоспинальные волокна прослеживаются до крестцовых сегментов.
Знание функциональных механизмов двигательных рефлексов позволяют невропатологу, нейрохирургу ставить топический диагноз повреждений, возникающих в структурах мозга и проводить соответствующие лечебные действия.
 Ствол мозга состоит из: мозга, и мозга.
Ствол мозга состоит из: мозга, и мозга.

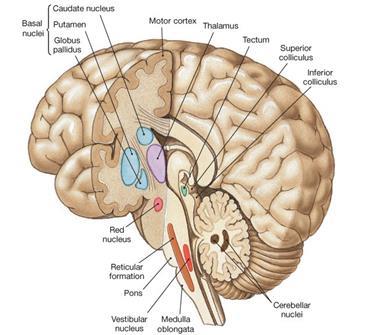
Рис. 16. Отделы ствола мозга, управляющие позой и движениями. ДИМА! мб ты облагородишь левый рисунок -сделаешь его похожим на правый рис: для чего Из правого рис убрать баз. гангл. а ретикул. формацию разделить как на левом рис. идр, а нарисовать две части РФ Нарисовать сегменты на уровне спинного мозга и в подписи к рис обратить на сегм. и ядерный принципы распределения нейронов в сп.и стволе.
Опишите функциональную роль областей ствола мозга, указанных на рисунке 16.
Верхние (передние) бугорки четверохолмия получают афферентные сигналы от сетчатки, нижние (задние) бугорки четверохолмия - от слуховых рецепторов и управляют движениями в ответ на зрительные, слуховые и соматические стимулы.
Ретикулярные ядра моста и продолговатого мозга функционируют по отношению друг к другу как антагонисты / синергисты (неправильное зачеркнуть). Ретикулярные ядра моста тормозят / возбуждают (неправильное зачеркнуть) антигравитационные мышцы. Ретикулярные ядра продолговатого мозга возбуждают / тормозят (неправильное зачеркнуть) антигравитационные мышцы. Эти ядра создают основной уровень тонического / фазического (неправильное зачеркнуть) сокращения для противостояния силам гравитации.
Все вестибулярные ядра вместе с ретикулярными ядрами моста управляют антигравитационными мышцами. Латеральные вестибулярные ядра возбуждают/ тормозят (неправильное зачеркнуть) позные мышцы , , верхних и
Медиальные вестибулярные ядра регулируют положение головы при воздействии на нее угловых ускорений.
Крупные нейроны магноцеллюлярной части красного ядра дают начало руброспинальному тракту, который заканчивается на вставочных и альфа - мотонейронах спинного мозга (рубро-спинальный путь). Нейроны красного ядра вызывают возбуждение альфа – мотонейронов сгибателей и усиливают их тонус.





