Оптимальное сопряжение на обменном участке параметров архитектоники, гемодинамики и кислородного обеспечения имеет важнейшее значение для адекватного снабжения кислородом сердечных клеток по «качеству» и «количеству». К аспекту «качества» кислорода следует отнести разность его напряжений на концах обменной артериолы, аспектом «количества» является объем кислорода, исходящий из микрососуда за единицу времени.
Установлено [68, 247], что кислород начинает поступать в ткани из прекапиллярных артериол. Наибольшее значение имеют генерации артериол с диаметром менее 40 мкм; из них в сердечную ткань поступает почти 12% от общего количества кислорода. «Вклад» сосудов с. d>40мкм в снабжение тканей незначителен. Для «кислородного» анализа нами выбрана цепочка из 10 обменных микрососудов вплоть до капилляра, двигаясь по которой эритроцит отдает кислород в окружающие ткани (см. рис. 3.7). Обозначим диаметры сосудов, начиная от капилляра и далее по направлению к аорте: m=1, 2,…10. Диаметры в этом направлении изменяются следующим

образом:
dm+1( bЗС) = dm( bЗС)/ 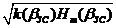 , (3.17)
, (3.17)
где dm(bЗС), dm+1(bЗС) - диаметры m- и m+1-сосудов цепочки, начиная от капилляра, в золотом режиме гипертензии. Рассмотрим «работу» обменной цепочки в сердце собаки. Диаметр сердечного капилляра собак dK(bЗС)=5,5 мкм [233]; эта величина для различных видов млекопитающих практически одинакова [78] Расчетные значения диаметров первых 10 звеньев приведены в Табл. 4.
Время пребывания эритроцитов в терминальных сосудах можно определить по формуле [203]:
tm+1( bЗС) = [С m+1( bЗС)/С m( bЗС)] tm( bЗС). (3.18)
Продолжительность прохождения эритроцита через артериальный участок сосудистого тракта составляет (15-20)% от общего времени [169]. В покое общее время пребывания эритроцита в коронарном русле собак при нормальном среднем давлении в аорте составляет 4 с [236]. По нашим расчетам количество артериальных генераций в сердце собак N=28 [203]. Примем время движения эритроцитов по артериальному руслу от аорты до капилляра равным 600 мсек (15% от общего времени). Выражение (3.18) справедливо для любых двух соседних звеньев цепочки с d<40 мкм, по которой движется отдельный эритроцит. Для сосудов с d> 40 мкм коэффициент конструкции «стандартный»: Cm(bЗС)=3 (см. Табл. 4, столбец 3). При приближении терминальных артериол к капилляру отношение Сm+1(bЗС)/Сm(bЗС), как нам уже известно, последовательно возрастает в пределах 1,33¸2. В соответствии с представленными величинами время пребывания в каждом «стандартном» сосуде tm(bЗС)=18,5 мcек. Количество генераций на обменном участке у всех млекопитающих практически одинаково, поскольку у всех животных диаметр обменных микрососудов изменяется в пределах от 40 до 7 мкм.
Исходя из представленных величин, рассчитаем время пребывания эритроцитов в 10 терминальных артериолах сердца млекопитающих (см. Табл. 4, столбец 4). В соответствии с параметрами, известными ранее и
Табл. 4. Величины конструктивных и кислородных параметров
в терминальных артериолах сердца [203]
| m
| dm(bЗС),
мкм
| Сm
| tm(bЗС),
мсек
| Um(bЗС),
%/сек
| DHbO2m(bЗС),
%
| DpO2m(bЗС),
мм рт. ст.
|
| 1
| 6,9
| 6
| 37,0
| 138,3
| 5,04
| 11,0
|
| 2
| 8,7
| 6
| 37,0
| 87,0
| 3,10
| 10,1
|
| 3
| 11,0
| 5
| 30,9
| 54,7
| 1,66
| 6,9
|
| 4
| 13,9
| 5
| 30,9
| 34,4
| 1,05
| 4,9
|
| 5
| 17,5
| 5
| 30,9
| 21,6
| 0,667
| 4,0
|
| 6
| 22
| 4
| 24,6
| 13,6
| 0,30
| 3,5
|
| 7
| 27,8
| 4
| 24,6
| 8,5
| 0,14
| 2,4
|
| 8
| 35,0
| 4
| 24,6
| 5,3
| 0,080
| 1,1
|
| 9
| 44,0
| 3
| 18,5
| -
| -
| -
|
| 10
| 55,6
| 3
| 18,5
| -
| -
| -
|
Примечания: m - номер обменного сосуда цепочки по направлению капилляр – аорта; tm ( b ЗС ) - время пребывания эритроцита в m -сосуде, Um ( b ЗС ) - скорость отдачи кислорода отдельным эритроцитом в поперечном сечении m -сосуда, D HbO2m ( b ЗС ) - количество кислорода, отданного отдельным эритроцитом за время пребывания в m-сосуде, D рО2m ( b ЗС ) – разность напряжений кислорода на концах m -сосуда в золотом режиме гипертензии.
вычисленными по (3.17) и (3.18), в первом от капилляра сосуде (m=1) ΔP1(bЗС)=3,5 мм рт. ст., t1(bЗС)=0,037 c, ξ1(bЗС)=1,45×10-2 Пз, d1(bЗС)=6,9×10-4 см. С учетом этих величин нами получены значения для других параметров: q1(bЗС)=41,9×10-9 см3×с-1, V1(bЗС)=1,53×10-9 см3, G1(bЗС) = 0,91×10-11 г-1×см5×с-1. Используя эти величины и С1(bЗС)=6 в качестве исходных, можно вычислить значения аналогичных параметров для других обменных звеньев цепочки.
Перейдем к непосредственному расчету кислородных параметров. Каждый эритроцит отдает кислород, перемещаясь по цепочке обменных артериол, включая капилляр. Следует заметить, что между капилляром и терминальными артериолами много общего. Стенки мельчайших артериол и капилляров имеют весьма сходное строение: эпителий, тончайшая базальная мембрана и сравнительно редкие и очень тонкие пучки гладких мышц. Все эти образования имеют такой же коэффициент диффузии для О2 и СО2, как и окружающие ткани. Эритроциты при перемещении в артериолах с диаметром менее 40 мкм так же, как и в капиллярах, принимают парашютообразную форму [292]. По расчетам Федершпиля и Попеля [252] эритроциты отдают кислород с одинаковой скоростью независимо от степени их отклонения от продольной оси сосуда. Экспериментально установлено [323], что количество кислорода, поступающее в ткани через единицу поверхности артериолы за единицу времени, не меняется при значительном изменении кровотока, величины гематокрита и степени насыщения кислородом движущейся крови. Другими словами, за единицу времени микрососуд всегда отдает постоянное количество кислорода независимо от представленных изменений в протекающей крови. Скорость отдачи кислорода обусловлена только величиной потребления кислорода окружающей тканью, а она по всей длине цепочки микрососудов, включая капилляр, одинакова. Следовательно, общее количество кислорода, поступающее за единицу времени из каждого поперечного сечения сосудов любого калибра, всегда одинаково. Естественно, «вклад» отдельного эритроцита в общую диффузию будет тем больше, чем меньше поперечная площадь сосуда. В капилляре отдельный эритроцит отдает кислород с максимальной скоростью. Очевидно, что соотношение скоростей отдачи кислорода отдельным эритроцитом в двух соседних микрососудах обратно пропорционально соотношению их поперечных площадей. Исходя из этих рассуждений, в m-сосуде по отношению к капилляру:
Um( bЗС) =U K( bЗС)S K( bЗС)/Sm( bЗС), (3.19)
где Um(bЗС), UK(bЗС), Sm(bЗС), SK(bЗС) - соответственно скорость отдачи кислорода отдельным эритроцитом и площадь поперечного сечения в m-артериоле и капилляре в золотом режиме гипертензии.
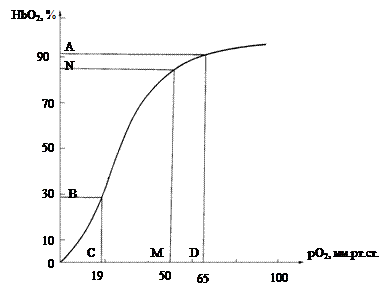
Установлено [37, 38], что напряжение кислорода на артериальном конце капилляров в тканях составляет 40-60 мм рт. ст. Примем для дальнейших расчетов в исходном режиме рО2(bЗС)=50 мм рт. ст., а в первой коронарной артерии рО2=95 мм рт. ст. На кривой диссоциации гемоглобина человека (см. рис. 3.8), взятой нами из книги Уайт и др. [176], этому интервалу соответствует разность насыщения кислородом ΔHbO2=12%. Показано [255], что в миокарде величины рО2 в крупных, средних и мелких артериях точно следуют величине рO2 в аорте. Диффузия кислорода начинается в артериолах диаметром менее 100 мкм. В микрососудах с диаметром более 40 мкм отдача кислорода отдельным эритроцитом очень мала и этой величиной при расчетах в дальнейшем можно пренебречь. При прохождении m-сосуда разность насыщения кислородом эритроцита кислородом в этом сосуде:
ΔHbO2( b ЗС) = U K( b ЗС)t m( b ЗС)[S K( b ЗС)/Sm( b ЗС)].
Рис. 3.8. Кривая насыщения гемоглобина человека [ 176 ]. По оси абсцисс – рО2 в мм рт. ст.; по оси ординат - HbO 2 в %. Точки A , B , C , D – границы логарифмического участка кривой (см. рис. 16). Интервал CD - pO 2 =19-65 мм рт. ст. интервал AB – HbO 2 =28,7-91,8%; точкам M и N соответствуют рО2=50 мм рт ст. и HbO 2 =85% на артериальном конце капилляра в золотом режиме гипертензии.
Для расчетов используем первые 8 генераций обменных сосудов (см. Табл. 4, столбец 6); при m>8 расчетные значения Um(bЗС) изменяются незначительно. Общий «разряд» эритроцита на 8 звеньях цепочки:
Δ HbO2( b ЗС) = UK( b ЗС) SK( b ЗС) 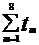 ( b ЗС)/ Sm( b ЗС). (3.20)
( b ЗС)/ Sm( b ЗС). (3.20)
Скорость отдачи кислорода эритроцитом в капилляре в золотом в режиме можно определить по формуле:
U K( b ЗС) = [ΔHbO2( b ЗС)/S K( b ЗС)]/ ( b ЗС)/Sm( b ЗС). (3.21)
Приняв ΔHbO2(bЗС)=12%, Sk(bЗС)=23,7×10-8 см2 и используя значения dm(bЗС) и tm(bЗС), приведенные в табл. 4, можно вычислить величину UK(bЗС), а также Um(bЗС) и ΔHbO2m(bЗС); последние приведены в табл. 4 (см. столбцы 5 и 6). Расчетная величина скорости отдачи кислорода UK(bЗС)=215%/с незначительно отличается от максимальной – 220 %/с. Эта величина была рассчитана и экспериментально установлена для движущихся эритроцитов в плазме с имитацией потребления кислорода [301, 211]. Наши расчеты подтверждают, что «в живых организмах скорость химических реакций всегда достигает предельного значения, которое определяется законами физики» [220, с. 96]. Для дальнейших расчетов будем использовать величину UK(bЗС)=220 %/с. В артериолах с диаметром менее 40 мкм скорость отдачи кислорода единичным эритроцитом быстро возрастает; растет и время пребывания эритроцитов в сосуде. Отметим, что экспериментально установлено [68, 70], что в артериолах мозга и мышц особо резкое падение рО2 происходит в сосудах диаметром 10-15 мкм и ниже. Эта экспериментально установленная тенденция совпадает с нашими расчетами (см. Табл. 4, столбец 7).
В покое на артериальном конце сердечного капилляра рО2(bЗС)»50 мм рт. ст. [38, 39], а на венозном рО2(bЗС)=20 мм рт. ст. [321]. Большое значение для равенства рО2 на артериальных концах капилляров имеет близость прекапиллярных тройников к полной симметрии [49]. Эритроциты в симметричных ветвях терминальных тройников отдают равное количество кислорода и входят в капилляры с приблизительно одинаковыми рО2, что соответствует одинаковому «качеству» кислорода. В противном случае, если бы вхождение и выход эритроцитов происходило со значительными отклонениями от величин рО2(bЗС)=50 мм рт. ст. и рО2(bЗС)=20 мм рт. ст., было бы нарушено условие одинакового «качества» кислорода, поступающего в сердечные ткани из капилляров.
Рассмотрим «количество» и «качество» кислорода, поступающего из обменных артериол. Как было показано выше, каждое поперечное сечение артериол независимо от их диаметра отдает такое же количество кислорода, что и сечение капилляра. Иными словами, количество кислорода, поступающее за единицу времени из микрососуда известной длины, равно объему кислорода из капилляра той же длины. «Качество» кислорода, поступающего из артериол в клетки, также равно «капиллярному». Это обусловлено тем, что чем больше диаметр сосуда, тем толще его стенка и, следовательно, больше падение величины рО2 на последней. Вследствие этого величины рО2 в ткани уже на незначительном удалении от стенки обменных артериол и капилляров приблизительно равны [71]. Представленное сходство условий диффузии кислорода в капиллярах и микрососудах позволяет сделать вывод, что с «кислородной» точки зрения последние ничем не отличаются от капилляров.Добавим к этому, что обменные артериолы, также как и капилляры, параллельны мышечному волокну, которое они обеспечивают кислородом [270, 55]. Таким образом, «кислородная работа» артериол ничем не отличается от капилляров за исключением одного обстоятельства. Разница заключается в «цене» кислорода. Энергетическая «стоимость» единичного объема кислорода, потребляемого из микрососудов, в артериолах выше, чем в капилляре. Эта «цена» тем больше, чем крупнее диаметр обменного микрососуда. Таким образом, с точки зрения дешевизны потребляемого кислорода, имеет место оптимальное сопряжение «противоположностей»: общее количество «дешевых» и общее количество «дорогих» «капилляров». Энергооптимальная гармония этих «противоположностей» обеспечивает оптимальные величины«количества» и «качества» кислорода, поступающего в кардиомиоциты, при минимальных затратах энергии.
Как будет показано в 4 главе (см. рис. 4.3), сегмент кривой диссоциации оксигемоглобина, соответствующий рО2=19-65 мм рт. ст., обладает особыми, логарифмическими, свойствами. Этими свойствами обусловлено оптимальное сопряжение «по кислороду» прекапиллярной и капиллярной частей обменного коронарного русла. Прекапиллярные сосуды являются своего рода переходной конструкцией, где происходит предварительная организация оптимальных условий для диффузии кислорода на важнейшей части обменного русла - капиллярах. Точка раздела «противоположностей» - «микрососудистой» и «капиллярной частей на кривой HbO2=f(pO2) - определяет оптимальное кислородное обеспечение миокарда по «количеству» и «качеству».
Отметим «золотую» особенность распределения скорости отдачи кислорода отдельным эритроцитом по цепочке обменных сосудов. Выше нами показано, что количество кислорода, отдаваемого всеми эритроцитами в каждом поперечном сечении любой артериолы за единицу времени, равно постоянной величине U(bЗС)=220%/с. Эта величина не зависит от диаметра микрососудов. Последовательность значений Um(bЗС)= 5,3; 8,5; 13,6; 21,6; 34,4; 54,7; 87,0; 138,3; 220 % с-1 (см. Табл. 4, столбец 5) близка к классическому ряду чисел Фибоначчи: 5, 8, 13, 21, 34, 55, 89, 144, 233. Представленная последовательность указывает на то, что скорость отдачи кислорода единичным эритроцитом в терминальных артериолах и в капилляре изменяется по аналогии с рядом чисел Фибоначчи. Эта аналогия сохраняется при любой гипертензии (см. раздел 4.5). Размеры и архитектоника терминальных микрососудов у различных видов млекопитающих практически одинаковы [76, 216]. Поэтому можно считать, что скорость отдачи кислорода отдельным эритроцитом в «одноименных» прекапиллярных сосудах сердца у млекопитающих (независимо от их размера и веса) одинакова. Скорости «кислородного разряда» эритроцита в последовательности терминальных артериол являются инвариантными оптимальными величинами, соотносящимися между собой по аналогии с рядом чисел Фибоначчи.