В общем виде для млекопитающих любого n-вида структуру сердечного цикла для всякого j-параметра (j-ССЦ), можно представить выражением:
C S j(nn,Wn) + CDj(nn,Wn) = Cj(nn,Wn), (2.1)
где nn, Wn- частота сердцебиений и вес животного n-вида, Csj(nn,Wn), CDj(nn,Wn), Cj(nn,Wn) - систолическое, диастолическое и суммарное значения некоторого j-параметра, соответствующие величинам nn и Wn . Выражение (2.1) является математическим представлением любой j-ССЦ.
Временную ССЦ каждого животного в соответствии с (2.1) можно представить выражением:
tS( n n ,Wn) + tD( n n ,Wn) = T( n n ,Wn), (2.2)
где tS(nn,Wn), tD(nn,Wn), T(nn,Wn) – соответственно длительности систолы, диастолы и сердечного цикла животного n-вида при частоте сердечных сокращений nn. Длительность систолы принята нами равной длительности электромеханической систолы, которая практически равна электрической систоле (см. рис. 2.1). Отметим, что временная ССЦ является «ведущей», определяющей, как будет показано ниже, структуру других сердечных параметров.
Важнейшее значение в организации эффективной деятельности сердца имеет синхронизация «противоположностей» всех рассматриваемых ССЦ. Синхронизация, как природное явление, известна давно. Еще в 17 веке Х. Гюйгенс (1629-1695) обратил внимание на удивительную синхронизацию часов, подвешенных на единое коромысло. Если даже вначале часы шли в разнобой, то через некоторое время у них вырабатывался единый ритм – они начинали работать синхронно. Как установил Гюйгенс, роль синхронизатора в этом случае играли слабые колебания коромысла, на котором подвешивались резонаторы – часы. Два резонатора в общей связующей среде (коромысло) всегда вырабатывали общий ритм в синхронизирующем процессе всей системы как единого целого. Можно привести биологический пример синхронизации. Н. Винер (1894-1964) заметил удивительное поведение светлячков в период весеннего хоровода. В вечерних сумерках деревья были облеплены этими насекомыми. Вначале световые мелькания светлячков были хаотичны. Но спустя некоторое время они становились организованными: множество живых существ синхронно выдавало световые вспышки в едином ритме. Отметим, в основе этого феномена лежит все та же резонансная синхронизация отдельных резонаторов (светлячков) в едином коллективном взаимодействии. В обоих примерах (механическом и биологическом) наблюдаются общие признаки: 1) независимая работа резонаторов (часов, светлячков), 2) наличие активной связующей среды (коромысло, воздух), 3) равноправная работа всех активных начал в едином синхронизированном процессе. Эти признаки являются сугубо функциональными. Они не зависят от природы резонаторов и связующей среды. И в то же время они являются факторами, обусловливающими резонансные основы функциональной гармонии целого. Универсальность явления синхронизации состоит в том, что оно не зависит от вида взаимодействия, не зависит от природы периодически движущихся систем (при этом отсутствует порог синхронизации). Синхронный режим (согласование периодических действий во времени и пространстве) достигается в живой природе при равенстве или соизмеримости частот рассматриваемых действий. При сколь угодно слабых взаимодействиях между элементами динамической системы их взаимное влияние в ходе эволюции обязательно приведет систему в синхронный режим. Добавим к этому, советский математик А.М. Молчанов показал, что «в диссипативных системах резонансы …неизбежны» [109,с. 196]. Эта общность становится фундаментальной при описании резонансных основ функциональной гармонии организма и его систем.
Было показано [217], что в сложных системах, имеющих в основе структуру золотой пропорции, возникают явления, близкие к резонансу. При отклонении от резонансного режима нарушается равновесие, начинают действовать затратные механизмы для поддержания нормального состояния системы. Резонанс, возникающий как следствие «золотой» гармонии «противоположностей», позволяет поддерживать устойчивость структур и процессов сердца с минимальными затратами энергии. С позиций синхронизации В.Н. Трифановым [172] была рассмотрена временная структура сердечного цикла. В режиме функциональной гармонии здесь можно выделить три активных центра, работающих совместно в едином хроноритме. В морфологическом плане – это резонаторы предсердий и желудочков в единой структуре сердца, порождают сердцебиения, в которых систола (два резонатора) и диастола (сердечная ткань) соотносятся между собой по золотой пропорции, и это является признаком нормального (здорового) режима сердцебиений. В функциональном (регуляторном плане) - это восходящие и нисходящие процессы между синусовым узлом и продолговатым мозгом (два резонатора) и возбуждение самого продолговатого мозга (среда). Между этими хронопроцессами в нормальном режиме функционирования устанавливается золотая пропорция, которая реализуется в сердечном ритме через синусовый узел. При резонансе синусовый узел порождает сердечный ритм, при котором систола и диастола миокарда соотносятся между собою по золотой пропорции (доля систолы составляет 38%, доля диастолы – 62% от длительности кардиоцикла). Два сердечных резонатора в равноправном взаимодействии между собой и средой в едином хроноритме создают гармонию, которая характеризуется золотыми «противоположностями». Установлено [57], что аналогичные соотношения между временными «противоположностями» - длительностями систолы и диастолы - имеют место и в предсердиях, и в желудочках. В дальнейшем нами будет показано (см. раздел 2.1.5), что «золотой» частоте сердцебиений соответствует минимальный расход энергии на исполнение деятельности сердца. Кроме того, при резонансе золотых «противоположностей» обеспечивается минимальность затрат энергии на поддержание стабильности сердечных систем.
(Отметим кстати, что К.П. Бутусовым [27] была показана связь золотого сечения с периодами обращения планет Солнечной системы. Было установлено, что Солнечная система находится вблизи максимального резонансного устойчивого состояния, а планеты движутся по орбитам с наименьшей концентрацией космической пыли, т.е. с минимальными затратами энергии.)
Экспериментально показано [285], что в покое у человека и млекопитающих (8 видов), несмотря на значительное различие в размерах и весе, соотношение между длительностью систолы и продолжительностью сердечного цикла определяется аналогичным по форме среднестатистическим выражением:
tS( n n, Wn)= kn 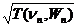 ,
,
где kn – постоянный коэффициент для всего диапазона сердцебиений животных n-вида, Wn – вес животного n-вида. Лепешкин установил, что для различных видов млекопитающих, включая и человека, в покое временнóе соотношение систолы и кардиоцикла находится в пределах 0,42 – 0,46, т.е. приблизительно одинаково. При известном kn, приравняв tS(nn,Wn)/T(nn,Wn)=0,382, можно для каждого вида определить среднестатистическую «золотую» частоту nnЗС, у которой длительности tS(nnЗС,Wn), tD(nnЗС,Wn) и T(nnЗС,Wn) соотносятся по золотой пропорции. Некоторое расхождение реальных частот в покое и вычисленных nnЗС объясняется тем, что лабораторные животные ведут менее активный образ жизни, чем их «вольные» сородичи. Поэтому у лабораторных
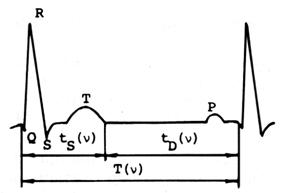
Рис. 2.1. Электрокардиограмма человека [193].
Q, R, S, Т, Р - зубцы электрокардиограммы, v –
частота сердцебиений; tS(ν), tD(ν), T(ν) - длительность
электрической систолы, диастолы и кардиоцикла при
частоте ν.
животных в покое частота сердцебиений выше «золотой». Отклонение «вверх» от золотого ритма у здоровых особей соответствует степени их детренированности (недостаточность или полное отсутствие регулярной физической нагрузки). Отметим, что у здоровых, физически активных людей и животных частота сердцебиений в покое и «золотая» частота nnЗС практически совпадают (например, для человека nnЗС=63 1/мин [190]).
В дальнейшем будем считать, что в покое у здоровых тренированных особей, ведущих естественный образ жизни, частота nnПОК » nnЗС; эту величину можно рассчитать, используя коэффициент kn.
Следует отметить, что у каждого i-животного n-вида имеет место «своя» золотая частота niЗС, которая может несколько отличаться от среднестатистической (niЗС @ nnЗС). Для отдельного i-животного нами были получены следующие изменения временной структуры кардиоцикла, связанные с изменением частоты сердцебиений ni по отношению к niЗС [193]:
tS( n i ,Wi) = tS( n iЗС ,Wi)/ 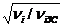 , (2.3)
, (2.3)
tD( n i ,Wi) = tD( n iЗС ,Wi)/( )3, (2.4)
T( n i ,Wi) = T( n iЗС ,Wi)/( n i / n iЗС ), (2.5)
где Wi – вес i-животного, niЗС, ni – «золотая» и другие частоты сердцебиений i-животного, tS(niЗС,Wi), tS(ni,Wi), tD(niЗС,Wi), tD(ni,Wi), T(niЗС,Wi), T(ni,Wi) – соответственно длительность систолы, диастолы и сердечного цикла при частотах niЗС и ni. Среднестатистическая ошибка в выражениях (2.3) и (2.4) во всех случаях не превышала предельной погрешности измерений. Это дает основание считать, что эмпирические выражения (2.3) и (2.4) подобраны правильно.
Таким образом, для любого i-животного при ni=niЗС выражение (2.2) можно представить выражением:
tS(niЗС,Wi)/ + tD(niЗС,Wi)/( )3 @ T(niЗС,Wi)/(ni/niЗС). (2.6)
Нас в дальнейшем будут интересовать изменения параметров, связанные с изменением отношения ni/niЗС, которое у различных видов млекопитающих изменяется приблизительно в одинаковых пределах. Эти изменения представляет универсальный показатель d =.ni/niЗС. Отметим, что показатель d имеет и другое «содержание». Для каждого животного при заданной физической нагрузке относительные изменения частоты сердцебиений и сердечного выброса за минуту аналогичны, поскольку объем выброшенной крови у млекопитающих за одно сокращение одинаков при любой нагрузке [193, 280, 313]. Следовательно, величину d можно представить в следующем виде:
d= n i / n i ЗС =S( n i ,Wi) /S( n iЗС ,Wi),
где S(niЗС,Wi,), S(ni,Wi) – соответственно сердечный выброс за минуту в золотом режиме кровоснабжения и при нагрузке, ni - частота сердцебиений i-животного при фиксированной нагрузке. Величина d для различных видов животных изменяется приблизительно в одних и тех же пределах (d»1-4) и не зависит от веса и размеров особи. В дальнейшем показатель d будет использоваться нами как универсальный показатель относительного изменения кровоснабжения сердца и всего организма млекопитающих в условиях физической нагрузки. Показатель d=dЗС=1 соответствует «золотому» режиму кровоснабжения животных.
Выражение (2.6) приобретает обобщенный вид:
tS( d , Wi)/  + tD( d ,Wi)/( )3 @ T( d ,Wi)/
+ tD( d ,Wi)/( )3 @ T( d ,Wi)/ 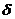 . (2.7)
. (2.7)
Выражение (2.7) с учетом «золотого» соотношения «противоположностей»:
0,382 T( dЗС, Wi)/ + 0,618 T( dЗС, Wi)/( )3 @ T( dЗС, Wi)/ . (2.8)
В обобщенном выражении (2.8) представлена гармония «противоположностей» во всех временных ССЦ различных видов млекопитающих. При d=dЗС=1 выражение (2.8) приобретает «классический» вид «золотой» структуры сердечного цикла:
0,382T( dЗС ,Wi) + 0,618T( dЗС ,Wi) = T( dЗС ,Wi).
Перейдем к анализу механической ССЦ. Показано [139], что по форме кривые артериального давления у человека и различных видов млекопитающих совершенно идентичны (см. рис. 2.2). Этот феномен позволяет нам исключить отдельный анализ этих кривых для различных видов животных.

Ри c . 2.2. Фазовая структура давления в аорте [ 190 ]. ν- частота сердцебиений. tS(ν), tD(ν), T(ν) - соответственно длительность электромеханической систолы, диастолы и сердечного цикла; РS(ν), PD(ν), p(ν,t) - соответственно среднее за систолу и среднее за диастолуи мгновенное значения давления в аорте при частоте сокращений v .
Рассмотрим структуру механической ССЦ. Средние за систолу и диастолу величины давления в аорте (см. рис. 2.2):
PS( n iЗС, Wi) = 1/ tS( n iЗС, Wi)  ,
,
PD( n iЗС, Wi) = 1/ tD( n iЗС, Wi)  ,
,
где PS(niЗС,Wi), PD(niЗС,Wi) – соответственно среднее за систолу и среднее за диастолу давление, p(niЗС,Wi,t) – мгновенное значение давления в аорте i-животного при частоте niЗС.
Среднее за кардиоцикл давление в аорте i-животного в золотом режиме кровоснабжения
 = 1/ T( n iЗС, Wi)
= 1/ T( n iЗС, Wi) 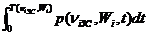 .
.
Анализ кривой давления (см. рис. 2.2), произведенный нами [190], показал, что PS(niЗС,Wi) @ PD(niЗС,Wi) = . Добавим к этому, что пульсации давления в коронарных сосудах по мере удаления от аорты снижаются. В мышечных артериях пульсации давления уже незначительны, а в микрососудах практически отсутствуют [341]. Это означает, что средние за систолу и диастолу давления в каждом из этих сосудов так же, как и в аорте, равны друг другу. Этот феномен имеет важное значение, поскольку позволяет обеспечить оптимальное крово- и кислородное обеспечение сердечной мышцы и в систолу, и в диастолу (см. раздел 3.4).
Величины среднего систолического и среднего диастолического давлений, отнесенные к длительности сердечного цикла,
 = [tS( n iЗС ,Wi)/T( n iЗС ,(Wi)]
= [tS( n iЗС ,Wi)/T( n iЗС ,(Wi)] 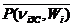 ,
,
 = [tD( n iЗС ,Wi)/T( n iЗС ,(Wi)] ,
= [tD( n iЗС ,Wi)/T( n iЗС ,(Wi)] ,
где  , - соответственно среднее систолическое и среднее диастолическое давления, отнесенные к длительности сердечного цикла при золотой частоте niЗС.
, - соответственно среднее систолическое и среднее диастолическое давления, отнесенные к длительности сердечного цикла при золотой частоте niЗС.
Вследствие того, что длительности систолы tS(niЗС,Wi) = 0,382T(niЗС,Wi) и диастолы tD(niЗС,Wi) = 0,618T(niЗС,Wi),
= 0,382 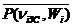 , (2.9)
, (2.9)
 = 0,618 . (2.10)
= 0,618 . (2.10)
На основе анализа экспериментальных данных, полученных у различных видов млекопитающих в условиях физической нагрузки, в аортальном давлении i-животного происходят следующие изменения [190]:
 = ni/niЗС , (2.11)
= ni/niЗС , (2.11)
 = , (2.12)
= , (2.12)
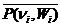 =
= 
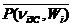 , (2.13)
, (2.13)
где , - соответственно среднее по отношению к длительности кардиоцикла систолическое и диастолическое давления, - среднее давление в аорте i-животного при частоте ni. Выражения (2.11) - (2.13) установлены при анализе экспериментальных данных, полученных у нескольких видов млекопитающих. Произведенный анализ выражений (2.11) - (2.13) показал, что среднестатистическая ошибка этих выражений не превышает предельной погрешности измерений. Отметим, что относительному изменению среднего давления в аорте соответствует одинаковое относительное изменение среднего давления на каждом сосуде [247].
Исходя из (2.9)-(2.13) и заменив ni/niЗС на d, получим математическую структуру механической ССЦ в условиях покоя и нагрузки:
0,382 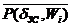 + 0,618 @ . (2.14)
+ 0,618 @ . (2.14)
В (2.14) представлена гармония «противоположностей» механической ССЦ в золотом режиме и при физической нагрузке. При d=dЗС=1 закон (2.14) представляет «золотую» структуру «противоположностей»:
0,382 + 0,618 = .
Рассмотрим математические особенности сопряжения «противоположностей» объемной ССЦ в условиях покоя и физической нагрузки. Предварительно отметим, что кривые сердечного выброса у различных видов млекопитающих совершенно идентичны [266, 267]. Эта аналогия позволяет нам не рассматривать особенности объемной ССЦ для отдельных видов млекопитающих.
Для объемной ССЦ среднее систолическое значение за сердечный цикл  соответствует объему крови, изгнанной из левого желудочка за одно сокращение, среднее диастолическое значение за сердечный цикл
соответствует объему крови, изгнанной из левого желудочка за одно сокращение, среднее диастолическое значение за сердечный цикл  равно объему оставшейся крови и суммарное значение
равно объему оставшейся крови и суммарное значение  - конечнодиастолическому объему. Экспериментально показано [207, 266, 267], что в покое (ni=niЗС) у человека и различных видов млекопитающих с разницей в весе в 1500 раз соотношение величин ,
- конечнодиастолическому объему. Экспериментально показано [207, 266, 267], что в покое (ni=niЗС) у человека и различных видов млекопитающих с разницей в весе в 1500 раз соотношение величин ,  и практически совпадает с золотой пропорцией:
и практически совпадает с золотой пропорцией:
0,382  + 0,618 = .
+ 0,618 = .
Это выражение представляет золотую гармонию «противоположностей» объемной ССЦ.
Установлено [271, 280], что при различной мышечной нагрузке объем крови, изгнанной за одно сокращение, постоянен. С другой стороны, показано, что чем больше нагрузка, тем в большей степени уменьшаются размеры желудочков [237, 315, 361]. В условиях физической нагрузки [190]:
=  , (2.15)
, (2.15)
=  /(
/(  ), (2.16)
), (2.16)
=  / , (2.17)
/ , (2.17)
где 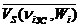 , , - соответственно среднее систолическое, среднее диастолическое и суммарное (конечнодиастолическое) значения объема левого желудочка i-животного, отнесенные к длительности кардиоцикла, в золотом режиме и при физической нагрузке.
, , - соответственно среднее систолическое, среднее диастолическое и суммарное (конечнодиастолическое) значения объема левого желудочка i-животного, отнесенные к длительности кардиоцикла, в золотом режиме и при физической нагрузке.
При учете (2.3) – (2.5) и (2.15) – (2.17) и замене ni/niЗС на d получим обобщенный закон объемной ССЦ:
0,382 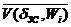 + 0,618 / d2 @ / d. (2.18)
+ 0,618 / d2 @ / d. (2.18)
В (2.18) представлена гармония систолической и диастолической «противоположностей» объемной ССЦ в условиях покоя и нагрузки.
Рассмотрим особенности кровотоковой ССЦ в условиях покоя и нагрузки. Форма пульсаций кровотока в крупных коронарных артериях аналогична [311; 347]. Установлено [320], что соотношения между значениями коронарного кровотока в характерных точках кардиоцикла не меняются при любом уровне нагрузки. Отмеченные особенности позволяют нам рассматривать общий коронарный кровоток, не рассматривая кровотоки в отдельных артериях.
Средние величины коронарного кровотока за систолу и диастолу в золотом режиме кровоснабжения:
QS(niЗС,Wi) = 1/tS(niЗС,Wi)  ,
,
QD(niЗС,Wi) = 1/tD(niЗС,Wi)  ,
,
 = 1/T(niЗС,Wi)
= 1/T(niЗС,Wi) 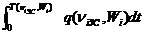
где QS(niЗС,Wi), QD(niЗС,Wi) – соответственно средний за систолу и средний за диастолу кровотоки, - средний за кардиоцикл кровоток, q(niЗС,Wi,t) – мгновенное значение кровотока i-животного при золотой частоте niЗС. Отметим, что пульсации кровотока в коронарных сосудах по мере удаления от аорты заметно снижаются; в мышечных артериях и микрососудах средние кровотоки в систолу и диастолу отличаются незначительно [341]. Следовательно, во всех мышечных и терминальных сосудах QS(niЗС,Wi)=QD(niЗС,Wi)= .
Средние величины систолического и диастолического кровотоков, отнесенные к длительности сердечного цикла:
 = [tS(niЗС,Wi)/T(niЗС,Wi)]
= [tS(niЗС,Wi)/T(niЗС,Wi)]  ,
,
 = [tD(niЗС,Wi)/T(niЗС,Wi)] ,
= [tD(niЗС,Wi)/T(niЗС,Wi)] ,
где , - соответственно средний систолический и средний диастолический кровотоки, отнесенные к длительности кардиоцикла i-животного при золотой частоте niЗС.
Исходя из (2.3) – (2.5),
 = 0,382
= 0,382  , (2.19)
, (2.19)
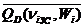 = 0,618[ . (2.20)
= 0,618[ . (2.20)
Качественная сторона гемодинамических изменений в коронарном русле во время физической нагрузки достаточно хорошо изучена. Рассмотрим некоторые данные, полученные в условиях нагрузки. Установлено, что при любой фиксированной мышечной нагрузке относительные изменения кровотоков к их величинам в покое одинаковы во всех ветвях левой коронарной артерии [280, 347] и эндо- и эпикардиальных слоях сердечной мышцы [229]. Не меняется отношение среднего кровотока в коронарном синусе к среднему коронарному кровотоку в левой коронарной артерии при изменении давления в аорте [282] и сжатии легочной артерии [309]. Отметим, что средний коронарный кровоток 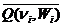 и минутный выброс сердца в покое и при нагрузке связаны между собою постоянным соотношением 0,04-0,05, одинаковым для человека и млекопитающих [189, 317]. Как было представлено нами выше, объем крови, изгнанной за одно сокращение сердца, постоянен при любой нагрузке. Следовательно, средний коронарный кровоток при нагрузке возрастает при частоте ni в ni/niЗС раз. Очевидно, что во всех крупных, мелких и средних коронарных артериях средний кровоток при нагрузке возрастает в ni/niЗС раз. Как следствие этого феномена, анализ относительных изменений в одной из коронарных артерий, полученных при нагрузке, позволяет получить представление об аналогичных изменениях общего среднего кровотока. В условиях физической нагрузки по отношению к золототому режиму кровоснабжения сердца составляющие кровотоковой ССЦ изменяются следующим образом [190]:
и минутный выброс сердца в покое и при нагрузке связаны между собою постоянным соотношением 0,04-0,05, одинаковым для человека и млекопитающих [189, 317]. Как было представлено нами выше, объем крови, изгнанной за одно сокращение сердца, постоянен при любой нагрузке. Следовательно, средний коронарный кровоток при нагрузке возрастает при частоте ni в ni/niЗС раз. Очевидно, что во всех крупных, мелких и средних коронарных артериях средний кровоток при нагрузке возрастает в ni/niЗС раз. Как следствие этого феномена, анализ относительных изменений в одной из коронарных артерий, полученных при нагрузке, позволяет получить представление об аналогичных изменениях общего среднего кровотока. В условиях физической нагрузки по отношению к золототому режиму кровоснабжения сердца составляющие кровотоковой ССЦ изменяются следующим образом [190]:
 = (ni/niЗС)3/2 , (2.21)
= (ni/niЗС)3/2 , (2.21)
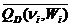 =
=  , (2.22)
, (2.22)
= ni/niЗС  . (2.23)
. (2.23)
Исходя из (2.3) – (2.5), (2.21) – (2.23) и заменив ni/niЗС на d, получим обобщенный математический закон композиции кровотоковой ССЦ млекопитающих в условиях покоя и физической нагрузки:
0,382 d3  + 0,618 d @ d2 . (2.24)
+ 0,618 d @ d2 . (2.24)
В (2.24) представлен математический закон сопряжения «противоположностей» кровотоковой ССЦ. В условиях покоя, когда d=dЗС, (2.24) приобретает классическую «золотую» форму «противоположностей»:
0,382 + 0,618 @ .
Обобщим законы (2.8), (2.14), (2.18) и (2.24). В золотом режиме общий закон композиции всех рассматриваемых ССЦ можно представить обобщенным выражением:
0,382 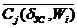 + 0,618 = , (2.25)
+ 0,618 = , (2.25)
где - среднее за кардиоцикл (суммарное) значение j-параметра i-животного в золотом режиме кровоснабжения организма.
Для средних (суммарных) значений j-параметров различных видов млекопитающих экспериментальным путем получены среднестатистические зависимости от веса i-животного [190, 264, 265, 284, 299; 327-329]:
= ajWibj, (2.26)
где aj, bj - величины, соответствующие j-параметру сердца (эти величины имеют постоянное значение независимо от веса животных). С учетом (2.25) – (2.26), выражение (2.25) приобретает вид:
0,382 ajWibj + 0,618 ajWibj = ajWibj. (2.27)
В выражении (2.27) «присутствует» «золотая» пропорция:
ajWibj: 0,618 ajWibj = 0,618 ajWibj : 0,382 ajWibj = 1,618…
Естественно, изменением веса животных в течение кратковременной мышечной нагрузки можно пренебречь. С учетом (2.26) и представленных выше зависимостей (2.8), (2.14), (2.18) и (2.24), влияние физической нагрузки на величину систолических, диастолических и суммарных элементов ССЦ представлено нами в обобщенном законе [193]:
0,382 
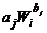 +0,618
+0,618  @
@  , (2.28)
, (2.28)
где kj - величина, соответствующая j-параметру (эта величина не зависит от уровня физической нагрузки и веса животных). Естественно, что при d=dЗС=1 выражение (2.28) предстает в виде (2.27). Отметим, что относительное расхождение между левой и правой частями приблизительного тождества (2.28) в заданных пределах (δ = 1¸4) составляет не более (-3 - +7)%, причем расхождение в 7% соответствуют верхнему уровню нагрузки. Отметим, что расхождение (-3 - +7)% находится в пределах ±(5-6)%-отклонения параметра от оптимального значения, при которых оптимальность системы еще сохраняется [23]. Закон (2.28) с достаточной точностью отображает реальные соотношения между «противоположностями» во всех ССЦ. В (2.28) представлен обобщенный математический закон композиции «пространства» (системы) временных, механических, объемных и кровотоковых ССЦ различных видов животных в условиях покоя и физической нагрузки.
Рассмотрим последовательность «сопряжения» принципа оптимального вхождения и гармонии «противоположностей» в законе (2.28). Первым этапом «сопряжения» является «подбор» оптимальной величины среднего (суммарного) значения j-параметра по отношению к весу i-животного. «Противоположности» на этом «этапе» представлены участками «положительных» и «отрицательных» приращений относительно оптимальной «рабочей» величины параметра. На втором этапе происходит «подбор» оптимального соотношения «противоположностей» (систола - диастола) для среднего (суммарного) значения j-параметра. Первый этап сопряжения гармонии «противоположностей» и принципа оптимального вхождения представлен постоянными величинами aj и bj; второй связан с числами 0,382 и 0,618. Отметим, вторая стадия сопряжения соответствуют золотому режиму кровоснабжения сердца. При «возмущении» организма физической нагрузкой «появляются» новые числа. Происходит «подбор» оптимальной средней величины каждого j-параметра в соответствии с уровнем нагрузки d. Среднее значение j-параметра разделено на «противоположности» (систола - диастола) в соответствии с величиной d. При нагрузке оптимальное вхождение j-«противоположностей» представлено числами kj, kj+1 и kj-1 и величиной d.
Табл. 1. Постоянные j-величины «пространства» ССЦ
|
j-ССЦ
| j-коэффициенты
|
| aj
| bj
| kj
|
| Временная
| 0,249
| 0,250
| -2
|
| Механическая
| 1,47×105
| 0,033
| 1
|
| Объемная, мл
| 1,76
| 1,02
| -1
|
| Кровотоковая
| 7,48
| 0,81
| 2
|
Примечание. Во всех расчетах по (2.28) временная, механическая, объемная и кровотоковая ССЦ соответственно имеют размерность сек, дн/см2, мл, мл/мин, [Wi] - кг.
Таким образом, сопряжение принципа оптимального вхождения и гармонии оптимальных «противоположностей» на всех этапах построения «пространства» (2.28) имеет математическую базу. Величины 0,382, 0,618, aj, bj, kj, kj+1, kj-1 и диапазон d=1÷4 образуют числовой каркас «пространства» всякого i-животного (2.28). Эти «особые» числа постоянны для различных видов млекопитающих независимо от их вида и размеров [193]. Таким образом, в «особых» числах представлена математическая основа оптимальной композиции «пространства» ССЦ. Отметим, что универсальные золотые числа 0,382 и 0,618 «присутствуют» во всех j-параметрах. Величины aj, bj, и kj представлены в Табл. 1.
Следует обратить внимание на еще одну особенность деления j-ССЦ при нагрузке. В покое «вхождение» «противоположностей в j-ССЦ представлено «чистыми» золотыми числами, а при нагрузке – золотыми числами + определенными отклонениями для каждого d. Экспериментально показано (см. раздел 2.1.5), что и при отклонениях «противоположностей» от пропорции золотого сечения имеет место экономия энергии, максимальная для данного уровня нагрузки. Поэтому все «противоположности» в диапазоне d=1÷4 можно условно считать «золотыми», поскольку они имеют в своей основе «чистое» золотое сечение + оптимальные отклонения от последнего.






