ИНТЕРАКТИВНЫЕ ЛЕКЦИИ
Дробление зиготы человека
Процесс оплодотворения резко активизирует метаболизм яйцеклетки человека, выводя её из анабиотического состояния. Прежде всего, значительно возрастает потребление кислорода, усиливается углеводный и фосфатный обмен, начинается интенсивный синтез белков. Одним из важнейших результатов оплодотворения является инициация дробления яйца путем последовательных митотических делений на все более мелкие клетки - бластомеры.
Дробление зародыша происходит в течение 4 суток после образования зиготы. Оно происходит в маточных трубах по мере продвижения зиготы в полость матки. Этот период называют трубным периодом эмбриогенеза. Яйцеклетка человека – вторично изолецитального типа.
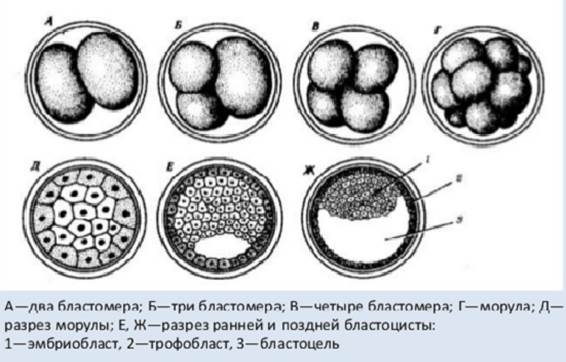
Рис. 2. Дробление зиготы человека
Дробление зародыша человека полное, неравномерное, асинхронное. С самого начала появляется полоса дробления по меридиану с образованием 2 бластомеров неодинаковой величины. Затем образуется 3 бластомера. Увеличение числа бластомеров происходит в начале медленно. Через 30 часов образуются 2 бластомера, через 40 часов – 4 бластомера. В ходе дробления появляются крупные темные бластомеры и мелкие светлые бластомеры. В процессе дробления мелкие бластомеры располагаются снаружи от крупных. Мелкие называются трофобластами, а крупные – эмбриобластами. Трофобласты обеспечивают трофику эмбриобласта.
К 3 суткам дробление ускоряется и к середине 3 суток образуется зародыш, который называется морулой, в ней отсутствует полость (стадия 12-ти бластомеров).
В начале 4 суток формируется бластоцист (бластула), имеющий полость. Стенкой полости является трофобласт. Эмбриобласт оттеснен к одному из полюсов бластоцисты и называется зародышевым узелком. Этот зародыш находится в маточной трубе и к 5 суткам состоит из 58 бластомеров. Бластодерма — слой ядер, а позднее клеток, из которых состоит зародыш с неполным дроблением на стадии бластулы.
К концу 5- началу 6 суток бластоцист попадает в полость матки. К этому времени он состоит из 108 бластомеров.
К концу 7 суток зародыш находится в матке в свободном виде и подготавливается к имплантации – внедрению в эндометрий (рис. 3).
ИМПЛАНТАЦИЯ - это погружение бластоцисты в функциональный слой эндометрия матки.
Она включает 2 фазы: 1) адгезию и 2) инвазию.
АДГЕЗИЯ - это приклеивание. Во время адгезии бластоциста приклеивается к эпителию эндометрия вблизи маточной железы. В это время в трофобласте появляется второй бесклеточный слой, который называется симпластотрофобластом, или синцитиотрофобластом. 1-й слой - клеточный, называется цитотрофобластом. В трофобласте в это время синтезируются и накапливаются протеолитические ферменты.
ИНВАЗИЯ характеризуется тем, что протеолитические ферменты растворяют сосуды. Бластоциста погружается в образовавшуюся ямку (имплантационную ямку), заполненную продуктами распада функционального слоя эндометрия. Это все происходит на микроскопическом уровне, так как величина зародыша в это время незначительно превышает размеры яйцеклетки (180 мкм в диаметре). После погружения зародыша в имплантационную ямку на 9-е сутки вход в эту ямку закрывается нарастающим эпителием эндометрия матки.
Одновременно с инвазией начинается 1-я фаза гаструляции (деламинация). Таким образом, в это время зародыш оказывается изолирован от полости матки. Продукты распада эндометрия, заполняющие имплантационную ямку, всасываются трофобластом зародыша и являются для него питательным веществом. Такой тип питания называется гистотрофным, который продолжается до конца 2-й недели эмбриогенеза, затем сменяется гематотрофным типом.
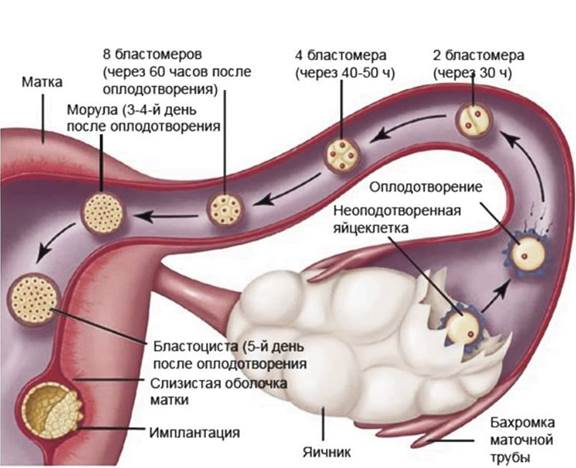
Рис. 3. Концептус от оплодотворения до имплантации
Считается, что этап дробления имеет следующие характерные черты:
1) дробящийся зародыш не растет;
2) не меняется его внешняя форма, но внутри образуется первичная полость тела - бластоцель;
3) после каждого деления количество ДНК в ядрах удваивается, поэтому суммарное содержание ДНК в зародыше непрерывно возрастает;
4) характер структурной гетерогенности ооплазмы в процессе дробления не меняется;
5) в бластомерах восстанавливается нормальное ядерно-плазменное отношение.
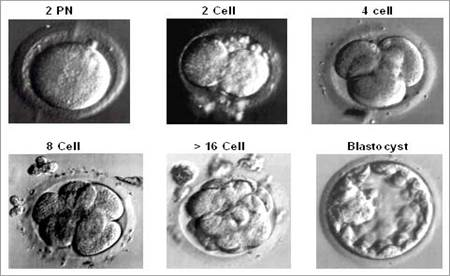
Рис.4. Микрофотография дробления зиготы человека
3.3 ГАСТРУЛЯЦИЯ
После дробления, которое завершается образованием бластулы или морулы начинаются направленные перемещения клеточных масс, которые ведут к образованию двухслойного зародыша – гаструлы. Процесс формирования из бластулы гаструлы называется гаструляцией, а возникающие при этом клеточные слои – зародышевыми листками.
Способы гаструляции довольно разнообразны. Отчасти они связаны со строением бластулы. При гаструляции наблюдается сочетание нескольких основных типов движений (способов гаструляции).
ИНВАГИНАЦИЯ
Представляет собой вворачивание участка клеточной стенки зародыша подобно тому, как вдавливается внутрь мягкая стенка резинового мяча, когда на него нажимают. При инвагинации механическая целостность стенки бластулы не нарушается. Впячивание одного из участков бластодермы внутрь осуществляется целым пластом (рис. 5).
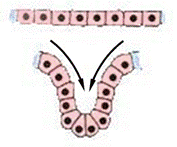
Рис. 5. Схема процесса инвагинации: А – инвагинационная гаструла; Б – движение пласта клеток
Процесс инвагинации возможен только в яйцах с небольшим или средним количеством желтка. В результате образуется двуслойный мешок, наружной стенкой которого является первичная эктодерма, а внутренней – первичная энтодерма. Бластоцель (первичная полость тела или полость Бэра) постепенно вытесняется, в ходе впячивания образуется гастроцель - первичный кишечник или архентерон, а отверстие, при помощи которого он сообщается с внешней средой, называют бластопором или первичным ртом. Последний, со всех сторон окружен губами бластопора.
Судьба бластопора у разных животных неодинакова. У многих животных первичный рот, развиваясь и дифференцируясь, превращается в дефинитивный рот взрослого организма - первичноротые (черви, моллюски и членистоногие). Не менее обширна группа вторичноротых, у которых бластопор превращается в анальное отверстие (щетинкочелюстные, плеченогие, иглокожие, кишечнодышащие) или в нервно-кишечный канал, находящийся в заднем конце эмбриона (хордовые).
ИНВОЛЮЦИЯ
При этом происходит вворачивание внутрь зародыша увеличивающегося в размерах наружного пласта клеток, который распространяется по внутренней поверхности остающихся снаружи клеток (рис. 6).
а  б
б 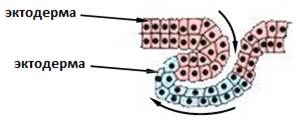
Рис. 6. Схема инволюции:
А - исходный пласт клеток, Б – движения клеточного пласта в ходе инволюции
ИММИГРАЦИЯ
– эволюционно наиболее древний способ гаструляции, открытый М.И.Мечниковым в 1884 г. Иммиграционная гаструла характерна для многих кишечнополостных. Она развивается вследствие активного выселения (ингрессии) части клеток стенки бластулы внутрь бластоцеля. Это выселение клеток может происходить как с одного (вегетативного) полюса – униполярная иммиграция, так и с двух противоположных (анимального и вегетативного) полюсов – биполярная иммиграция (рис. 7).
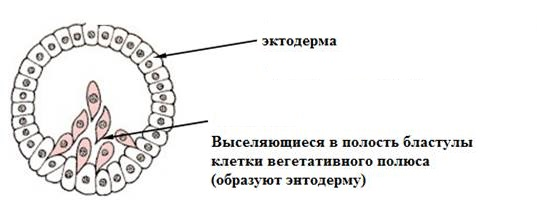
Рис. 7. Униполярная иммиграция (схема)
Иногда процессы иммиграции идут без особого порядка по всей поверхности бластулы - мультиполярная иммиграция. Выселившиеся клетки позднее образуют внутренний слой гаструлы – энтодерму. Таким образом, как и в ходе инвагинации зародыш становится двухслойным.
Важной особенностью иммиграционной гаструлы является отсутствие бластопора, а значит нет и характерного для инвагинационной гаструлы сообщения гастроцеля с внешней средой.
При иммиграции происходит перемещение отдельных клеток или групп клеток, не объединенных в единый пласт. Иммиграция, в том или ином виде, встречается у всех зародышей, но в наибольшей степени характерна для второй фазы гаструляции высших позвононых (птиц и млекопитающих).
ДЕЛАМИНАЦИЯ (расслаивание)
В тех случаях, когда дробление заканчивается образованием бластулы с невыраженной или почти отсутствующей полостью (бластоцелем), например морулы у некоторых кишечнополостных, наблюдается расщепление единого клеточного пласта на два более или менее параллельных. Каждая клетка, образующая бластодерму, путем митотического деления расщепляется и отшнуровывает второй слой (рис. 8).
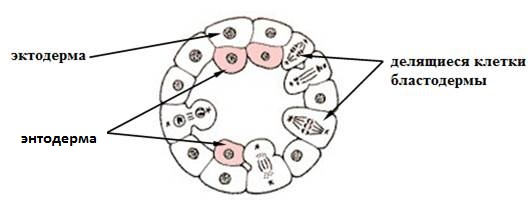
Рис. 8. Деляминация (схема)
При деляминации клеточные перемещения практически отсутствуют. Этот тип гаструляции ограничивается выравниванием внутренних стенок клеток наружного слоя, причем такое выравнивание нередко идет волной от одной соседней клетки к другой. Вдоль выровненных поверхностей формируется базальная мембрана, отделяющая внешний клеточный слой (эктодерму) от внутренней массы клеток, которая вся становится энтодермой. Деляминацию можно наблюдать у зародышей с неполным типом дробления, таких как пресмыкающиеся, птицы, низшие млекопитающие.
ЭПИБОЛИЯ
Представляет движение эпителиальных пластов клеток (обычно эктодермальных), которые распространяются как одно целое, а не индивидуально и окружают глубокие слои зародыша. Другими словами, происходит обрастание мелкими клетками анимального полюса более крупных, отстающих в скорости деления и менее подвижных, клеток вегетативного полюса (рис. 9).
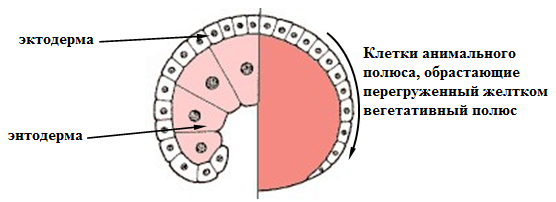
Рис. 9. Эпиболия (схема)
Формирование эпиболической гаструлы ярко выражено у земноводных и некоторых организмов с резко телолецитальным строением яиц. В результате у зародышей таких животных бластопор отсутствует и архентерон не формируется. Только впоследствии, когда макромеры делясь становятся меньшего размера, образуется полость - формируется зачаток первичного кишечника.
Все описанные типы гаструляции редко встречаются отдельно, обычно комбинируются: например: инвагинация с эпиболией, деламинации с иммиграцией. В результате принято выделять последний, смешанный, способ гаструляции.
Закладка мезодермы и формирование целома
После завершения гаструляции материал, оставшийся на поверхности зародыша, формирует наружный зародышевый листок – эктодерму, а погрузившийся любым способом внутренний зародышевый листок (энтодерма) образует впоследствии стенку пищеварительного тракта с его производными. Кроме того, погрузившийся внутрь материал помимо энтодермы содержит и закладку среднего зародышевого листка мезодермы, который потом отделяется от энтодермы.
Таким образом, в связи с гаструляцией, параллельно с ней или на следующем, обусловленном ей же, этапе у всех животных, за исключением кишечнополостных, возникает мезодерма. Это совокупность клеточных элементов залегающих между эктодермой и энтодермой, т.е. в бластоцеле.
В настоящее время вопросы образования мезодермы у животных, в общем, могут считаться решенными в плоскости формально-морфологического описания этих процессов. Если отвлечься от разнообразных деталей формирования мезодермы у разных животных, то можно выделить два принципиально разных пути ее образования.
· ТЕЛОБЛАСТИЧЕСКИЙ
У первичноротых во время гаструляции на границе между эктодермой и энтодермой, по бокам бластопора, уже имеются две большие клетки (или несколько таких клеток) – телобласты, получившие в ходе делений дробления всю полярную ооплазму. Впоследствие они отделяют от себя мелкие клетки и формируют, таким образом, средний пласт – мезодерму. Постепенно телобласты, давая новые и новые поколения клеток мезодермы, отодвигаются к заднему концу зародыша. По этой причине такой способ закладки называют телобластическим.
· ЭНТЕРОЦЕЛЬНЫЙ
Принципиально другой способ закладки мезодермы наблюдается у вторичноротых животных: иглокожих, бесчерепных (ланцетника), хордовых. Материал будущей мезодермы вворачивается вместе с энтодермой в составе единого гастрального впячивания, и в процессе инвагинации граница между обеими закладками, как правило, не различима. Такое впячивание, включающее в себя материал и энтодермы, и мезодермы, а у хордовых еще и хорды, называется первичным кишечником, тогда как гастроцель представляет собой полость первичной кишки (рис. 10).
ПЛАЦЕНТА.
Плаценты млекопитающих разделяются на 4 вида: 1) эпителиохориальные; 2.) синдесмохориальные, или десмохориальные; 8) эндотелиохориальные и 4) гемохориальные.
В зависимости от типа питания (трофики) различают 2 типа плацент.
Этапы дифференцировки
1. Химическая разнородность цитоплазмы яйцеклетки (усиливается после оплодотворения).
2. Химическая разнородность цитоплазмы бластомеров
3. В разных бластомерах разные индукторы включают разные транскриптоны
4. Синтезируются разные белки-ферменты, которые катализируют разные типы биохимических реакций.
5. В разных бластомерах идет синтез разных типо- и тканеспецифических белков.
6. Образуются разные типы клеток, создается морфологическая разнородность.
7. Разные типы клеток образуют разные ткани.
8. Разные ткани формируют разные органы.
Механизмы морфогенеза:
1. Эмбриональная индукция – влияние группы клеток эмбриона на соседние клетки (Г.Шпеман, Г.Мангольд). Первичный индуктор (верхняя губа бластопора) детерминирует образование нервной трубки, затем индуцируется развитие хорды, а после этого пищеварительной трубки.
2. Морфогенетические поля (А.Г.Гурвич) – дистантные взаимодействия клеток электрической или гравитационной природы.
3. Градиент физиологической активности (Ч.Чайлд) – интенсивность обменных процессов выше в головном отделе зародыша по сравнению с хвостовым отделом, что оказывает пространственное регулирующее действие на морфогенез.
4. Позиционная информация клетки – при помощи межклеточных взаимодействий каждая клетка оценивает свое местоположение в зачатке органа, а затем дифференцируется в соответствии с этим положением.
6.ТЕРАТОГЕНЕЗ. КРИТИЧЕСКИЕ ПЕРИОДЫ ЭМБРИОГЕНЕЗА.
Процесс нарушения естественного хода эмбриогенеза под влиянием факторов окружающей среды называется тератогенезом (греч. teras – урод, чудовище). Тератоген – средовой (внешний) фактор, действующий на эмбрион или плод и вызывающий нарушение его строения или функционирования, не вызывая при этом изменения наследственных структур.
Эффект тератогенного воздействия зависит от ряда условий:
- Время действия тератогена. Действие тератогена до наступления стадии клеточной дифференциации приводит, как правило, к гибели зародыша. При действии тератогена в период дифференцировки органов возникают различные пороки развития. Тип порока зависит от чувствительности конкретного органа в момент воздействия вредного фактора;
- Доза и взаимодействие с другими факторами. Для большинства тератогенов существует дозо-зависимый эффект, т.е. чем выше доза, тем тяжелее поражение. В некоторых случаях тератогенность фактора повышается при наличии других факторов (например, тератогенное влияние антиконвульсантов усиливается в комбинации с другими препаратами).
- Материнские факторы. Риск возникновения тератогенно обусловленной аномалии зависит от индивидуального порога чувствительности материнского организма к тератогенному агенту.
Тератогенные факторы разнообразны по природе и среди них выделяют:
· физические факторы (механические или радиационные воздействия),
· химические факторы (лекарственные вещества, химические вещества, используемые в быту и промышленности и др.)
· биологические факторы (вирусы, микоплазмы, протозойные инфекции и другие внутриутробные инфекции).
Действие тератогенов приводит к появлению пороков развития. Врожденные пороки бывают первичные, в результате прямого действия тератогенов (пример: атрезия водопровода мозга) и вторичные, в результате осложнения первичных пороков (пример: водянка головного мозга).
Причины, механизмы развития, профилактику появления пороков развития изучает наука тератология.
Частота встречаемости пороков развития в популяциях человека 1-2%.
Разновидности врожденных пороков развития:
· агенезия – отсутствие органа (например, конечности);
· гипогенезия – недоразвитие органа (например, гонад);
· гипергенезия – усиленное развитие (например, полидактилия);
· атрезия – заращение естественных отверстий и каналов (например,
пищевода, ануса);
· эктопия – изменение места расположения органа (например,сердце с правой стороны).
Таблица 6.1. Часто встречаемые врожденные пороки развития
| Формы аномалий
| Определение
| Сроки развития (неделя)
|
|
| Анэнцефалия -
| отсутствие головного мозга, свода костей черепа выбухание оболочек и тканей
|
26
|
| Менингомиелоцеле -
| выбухание оболочек и тканей спинного мозга сквозь дефект в позвоночнике
|
28
|
| Экстрофия мочевого пузыря -
| отсутствие нижней части передней брюшной стенки и передней стенки мочевого пузыря
|
30
|
| Губа расщепленная (заячья) -
| дефект развития верхней губы
| 9-10
|
| Грыжа пуповинная (омфалоцеле) -
| из-за дефекта брюшной стенки наружу выходят внутренние органы, покрытые истонченными тканями пуповины отсутствие у детей мужского пола
|
10
|
| Гипоспадия -
| дистальной части мочеиспускательного канала, а его наружное отверстие находится в мошонке или промежности; у детей женского пола - отверстие мочеиспускательного канала открывается во влагалище
|
12
|
| Крипторхизм -
| задержка опущения яичек в мошонку
| 12
|
Причины развития врожденных пороков:
1) генетические (различные мутации);
2) экзогенные (действие факторов среды);
3) мультифакториальные (совместное действие факторов 1-й и 2-й групп);
4) взаимодействие частей зародыша (эмбриональная индукция).
С конца Х1Х в. существует представление о наличии в онтогенезе человека периодов наибольшей чувствительности к повреждающему действию разнообразных факторов. Эти периоды получили название критических. Например, эпидемия краснухи (немецкая корь) в 1963-1965 гг. вызвала гибель 20 тыс. плодов и рождение 30 тыс. детей с врожденными пороками развития. Тератогенными являются вирус простого герпеса и цитомегаловирус, простейший одноклеточный паразит Toxoplasma gondii. Все это говорит о том, что мать не полностью защищает организм от влияния внешней среды.
Предэмбриональный период. Тератогены оказывают действие на половые клетки, вызывая развитие гаметопатий (рис. 6.1.).
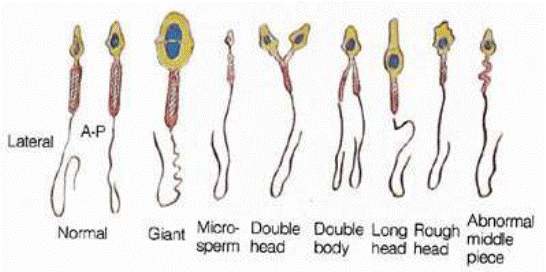
Рис. 6.1. Варианты патологически измененных морфотипов сперматозоидов человека
Пренатальны период. Тератогены оказывают действие на различные стадии эмбриогенеза, вызывая развитие эмбриопатий.
Для любого органа наиболее критическим периодом является время его роста и образования специфических для этого органа структур. Различные органы имеют различные критические периоды, однако период между 15 и 60 сутками – критический для многих органов. Сердце формируется между 3 и 4 неделями, наружные половые органы наиболее чувствительны к воздействиям между 8 и 9 неделями. Мозг и скелет чувствительны к вредным влияниям постоянно, начиная с 3 недели после зачатия до конца беременности и позже.
К критическим периодам относят:
· оплодотворение;
· имплантацию (7-8-е сутки эмбриогенеза);
· развитие осевого комплекса зачатков органов и плацентацию (3 - 8-я недели);
· развитие головного мозга (15 - 20-я недели);
· формирование основных систем организма, в том числе половой (20 - 24-я недели).
Действие тератогенных факторов во время эмбрионального (с 3 по 8 недели) периода может привести к врожденным порокам развития.
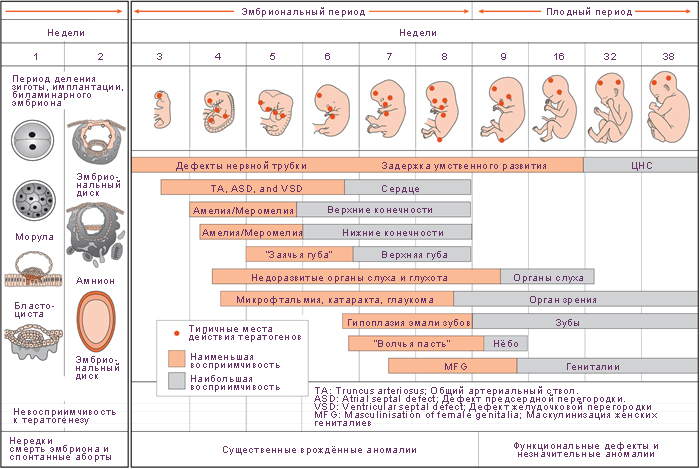
Рис. 6.2. Критические периоды развития человека (внутриутробный период)
Врожденные (анцестральные) пороки развития – это такие структурные нарушения, которые возникают до рождения и выявляются сразу или через определенный период после рождения.
Различают:
Аномалии – отклонения в развитии, которые не сопровождаются нарушением здоровья (родимые пятна, родинки, заячья губа). Функция органов не нарушается.
Пороки – это отклонения в развитии органов и тканей, которые сопровождаются нарушением состояния здоровья.
Уродства – это грубые нарушения органов и систем:
а) отсутствие органов
б) недоразвитие органов и систем – гипоплазия;
в) нарушение формы и расположения органов: в последнем случае это может быть пороком, если нарушено состояние здоровья, а может быть аномалией, если здоровье не нарушено;
г) изменение количества органов.
50% из всех пороков развития приходятся на опорно-двигательную систему. На 2-м месте пороки сердечно-сосудистой системы, на 3-м мочеполовой. Кривошея, диафрагмальные грыжи, врожденный вывих бедра, недоразвитие конечностей, увеличение количества позвонков, кисты желез. Отсутствие вторичного твердого неба у рыб и земноводных является филогенетической предпосылкой «волчьей пасти» у людей.
В постнатальном периоде, выделяют несколько критических периодов:
• новорождения - первые дни после рождения в связи с перестройкой всех процессов жизнедеятельности (питания, дыхания, выделения, кровообращения и др.);
• полового созревания - 12-16 лет, когда происходит гормональная перестройка;
• полового увядания - около 50-ти лет, когда происходит угасание функций эндокринных желез (особенно половых).
Причины критических периодов постнатального онтогенеза принципиально те же, что и пренатального: изменения гормонального фона, появление «новых» и исчезновение «старых» индукторов, включение и выключение разных блоков генов.
План
1. Связь онтогенеза и филогенеза. Биогенетический закон.
2. Филэмбриогенезы.
3. Филогенез нервной системы Хордовых.
4. Филогенез кровеносной системы Хордовых.
5. Онтофилогенетические предпосылки ВПР.
Связь онтогенеза и филогенеза. Биогенетический закон
Онтогенез (индивидуальное развитие) – это совокупность процессов развития организма с момента образования зиготы и до смерти, в ходе которого происходит преобразование его морфофизиологических, физиолого-биохимических и цитогенетических признаков, на основе реализации генетической информации в определенных условиях среды.
Филогенез – историческое развитие живых организмов: как всего органического мира Земли, так и отдельных таксонов (от царств до видов). Термин «филогенез» ввел Э. Геккель в 1866 г.
Понятия онтогенеза и филогенеза неразрывно связаны между собой: с точки зрения эволюционной теории, историческое развитие живой природы представляет собой чреду онтогенезов. Онтогенез - повторение филогенеза. Онтогенез - основа филогенеза.
Впервые взаимосвязь онтогенеза и филогенеза раскрыл К. Бэр в ряде положений. К. Бэр в 1828 г. сформулировал 3 закона связи онто- и филогенеза:
1) закон зародышевого сходства: на ранних этапах эмбрионального развития зародыши различных животных в пределах типа сходны между собой (например, разных классов подтипа позвоночных).
2) закон последовательности появления признаков различного систематического ранга: первоначально в эмбриональном развитии появляются признаки типа, затем подтипа, класса, подкласса, отряда, семейства, рода, вида и, наконец – индивидуальные.
3) закон эмбриональной дивергенции (расхождения признаков у зародышей): в процессе эмбриогенеза, по мере приобретения признаков различного систематического ранга, у зародышей различных животных в пределах типа возникают различия.
Объяснить эти законы можно так. На ранних этапах эмбриогенеза зародыши животных разных классов позвоночных (например, рыбы, птицы, млекопитающие) похожи между собой. Со временем между ними появляются различия в пределах классов, а дальше – в пределах отрядов (пример: зародыш свиньи и зародыш человека).
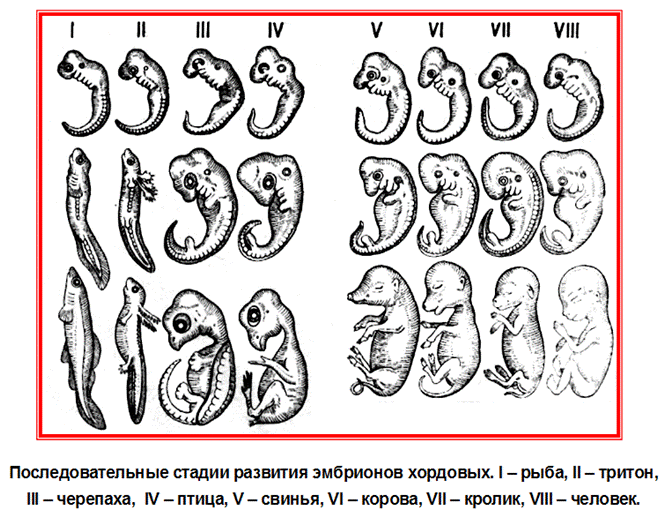
Закон параллелизма Меккеля-Серре говорит о том, что каждое существо в своем эмбриональном развитии повторяет взрослые формы животных более низких ступеней развития. Ч. Дарвин, подтверждая связь онтогенеза и филогенеза, создал учение о рекапитуляциях. Рекапитуляция – это повторение у зародышей признаков их предков. Действительно, например, зародыш человека на ранних стадиях развития похож на зародыш рыб и амфибий, на более поздних стадиях – на зародышей других млекопитающих, на самых поздних стадиях – на плод человекообразных обезьян.
В зародыше потомков, писал Ч. Дарвин, мы видим «смутный портрет предков». Другими словами, уже на ранних стадиях эмбриогенеза разных видов в пределах типа выявляется большое сходство. Следовательно, по индивидуальному развитию можно проследить историю данного вида. Именно эта мысль и была положена в основу биогенетического закона, сформулированного Ф. Мюллером и Э. Геккелем в 1866 году (также известен под названиями «закон Геккеля», «основной биогенетический закон»): каждое живое существо в своем индивидуальном развитии (онтогенез) повторяет, в известной степени, формы, пройденные его предками или его видом (филогенез), т.е. онтогенез есть краткое и быстрое повторение филогенез. Генетической основой биогенетического закона служит общность генов регуляции онтогенеза, которые передаются по наследству от предковых форм.
Однако оказалось, что рекапитуляции в онтогенезе могут быть не всегда. Не всегда наблюдается строгая последовательность повторения этапов филогенеза. Так, зародыш человека никогда не повторяет взрослых стадий рыб, амфибий и рептилий, а сходен только с их зародышами. Ранние стадии эмбриогенеза характеризуются большей консервативностью, поэтому рекапитулируют более полно, чем поздние.
Основываясь на своих знаниях и наблюдениях, Геккель выделил две группы признаков – палингенезы и ценогенезы. Палингенезы – это повторение у зародышей признаков предков (закладка хорды, жаберных щелей, двухкамерного сердца у наземных позвоночных. Ценогенезы – это эмбриональные адаптации, которые имеются только у зародышей и отсутствует у взрослых организмов (зародышевые оболочки у высших позвоночных – амнион, хорион, аллантоис).
Филэмбриогенезы
Трактовка биогенетического закона позднее была развита А.Н. Северцовым (1910-1939) в теории филэмбриогенезов. Основные положения теории:
1. Если бы не изменялся ход онтогенеза, то потомки не отличались бы от предков.
2. Посредством филэмбриогенеза может изменяться ход онтогенеза как целостного организма, так и отдельных органов, тканей и клеток.
3. Путём филэмбриогенеза происходят филогенетические изменения как взрослого организма, так и промежуточных стадий его развития.
Северцов разделял взгляды Геккеля и Мюллера о первичности онтогенетических изменений по отношению к изменениям взрослых организмов и рассматривал онтогенез не только как результат филогенеза, но и как его основу. Онтогенез не только удлиняется путем прибавления стадий: он весь перестраивается в процессе эволюции; он имеет свою историю, закономерно связанную с историей взрослого организма и частично ее определяющую.
Филэмбриогенезы – изменения в онтогенезе, которые переходят во взрослое состояние и обеспечивают приспособление взрослых форм к условиям внешней среды. Как правило, возникают в ходе морфогенеза (органогенеза).
В зависимости от того, на каких стадиях эмбриогенеза происходят эти изменения, выделяют три типа филэмбриогенезов: архаллаксисы, девиации и анаболии.
Архаллаксисы – изменения на ранних стадиях эмбриогенеза. Они выражаются в изменении начальных процессов дифференцировки зачатков, в изменении начальной массы зачатков, в сдвигах места и времени закладки органов. Рекапитуляция в этом случае отсутствует. Примеры: у двудольных растений – закладка сразу двудольного зародыша; развитие волоса у млекопитающих, гомолога эмбриональной чешуи у рыб.
Изменения на ранних стадиях эмбриогенеза приводят к крупным преобразованиям онтогенеза и являются основным источником прогрессивной эволюции взрослых форм.
Девиации – изменения в развитии органа на средних стадиях его формирования. Рекапитуляция частичная. Примером может служить развитие сердца в онтогенезе млекопитающих. Вначале идет повторение стадий трубки, двухкамерного и трехкамерного образования, но стадия неполной перегородки между желудочками, характерной для пресмыкающихся, вытесняется развитием сразу полной перегородки, характерной для млекопитающих.
Анаболии – изменения на поздних стадиях развития органа. Их называют еще надставками. Рекапитуляция наблюдается, изменения возникают на поздних стадиях. Примерами анаболий являются изменения в строении скелета позвоночных (изгибы позвоночника, сращение швов в мозговом черепе), в дифференцировке мышц и в распределении кровеносных сосудов в организме человека и млекопитающих.
Посредством способов филэмбриогенеза может происходить как прогрессивное развитие (путём усложнения строения и функций организмов), так и регрессивное (путём упрощения строения и функций организмов вследствие приспособления их к новым, менее разнообразным условиям существования) (например, при паразитизме).
Эволюция чаще идет путем подбора анаболий, поэтому наблюдаются рекапитуляции, сходство эмбрионов на ранних стадиях развития и усложнение организации по мере появления новых систематических групп (типа хордовых, подтипа позвоночных, высших позвоночных и, наконец, человека). В процессе онтогенеза организм развивается как единое целое по определенному плану строения. Это обусловливает согласованность развития органов и систем, которая сложилась в процессе филогенеза.
Филогенез нервной системы Хордовых
Хордовым животным присущ трубчатый тип нервной системы. У них в эмбриональном периоде из эктодермы над хордой закладывается нервная трубка, которая у ланцетника сохраняется всю жизнь и выполняет функцию центрального отдела нервной системы, а у позвоночных она преобразуется в спинной и головной мозг. При этом головной мозг развивается из переднего отдела нервной трубки, а из остальной ее части - спинной мозг.
Головной мозг у позвоночных состоит из пяти отделов: переднего, промежуточного среднего, продолговатого мозга и мозжечка.
Функции нервной системы:
· регулирует работу всех систем органов;
· обеспечивает связь организма с окружающей средой;
· объединяет организм в единое целое;
· определяет психическую деятельность человека как социального существа.
Нервная система ланцетника представлена трубкой, которая расположена над хордой. Полость нервной трубки называется невроцель. В
передней расширенной части трубки находится обонятельная ямка. По всей нервной трубке расположены глазки Гессе (светочувствительные клетки).
Формирование головного мозга у зародышей всех позвоночных начинается с появления на переднем конце нервной трубки вздутий - мозговых пузырей. Первоначально закладываются 3 мозговых пузыря. Дальше делятся пополам первый и третий пузыри – образуется 5 мозговых пузырей, а из них 5 отделов головного мозга: 1 – telencephalon – собственно передний мозг –(полушария и боковые желудочки); 2 – diencephalon – промежуточный мозг (его полость – третий желудочек); 3 – mesencephalon – средний мозг; 4 – metencephalon (мозжечок); 5 – myelencephalon (продолговатый мозг).
Последний без резкой границы переходит в спинной мозг.
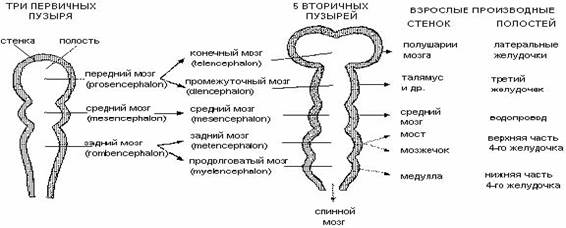
В нервной трубке есть полость - невроцель, которая в ходе образования пяти мозговых пузырей формирует расширения - мозговые желудочки (у человека их 4). В этих участках мозга различают дно (основание) и крышу (мантия). Крыша располагается над - а дно под желудочками.
Вещество мозга неоднородно - представлено серым и белым веществом. Серое - это скопление нейронов, а белое образовано отростками нейронов, покрытыми жироподобным веществом (миелиновой оболочкой), которое придает веществу мозга белый цвет. Слой серого вещества на поверхности крыши любого отдела мозга называется корой.
Большую роль в эволюции нервной системы играют органы чувств. Именно концентрация органов чувств на переднем конце тела обусловила
прогрессивное развитие головного отдела нервной трубки. Полагают, что передний мозговой пузырь сформировался под влиянием обонятельного, средний - зрительного, а задний - слухового рецепторов.
Эволюция нервной системы идет по пути дифференцировки нервной
трубки на головной и спинной мозг (центральная нервная система) и
дифференцировки периферической нервной системы (ганглии и нервные
волокна).
Более сложными являются направления эволюции головного мозга:
· увеличение его объема;
·дифференцировка отделов и образование изгибов (передний изгиб –в области среднего мозга, средний – изгиб моста, задний изгиб – в области
продолговатого мозга);
· развитие коры переднего мозга, борозд и извилин;
· развитие и совершенствование органов чувств.
РЫБЫ
Передний мозг небольшой, не разделен на полушарии, имеет только
один желудочек. Его крыша не содержит нервных элементов, а образована
эпителием. Нейроны сосредоточены на дне желудочка в полосатых
телах и в отходящих спереди от переднего мозга обонятельных
долях. По существу, передний мозг выполняет функцию обонятельного
центра.
Средний мозг является высшим регуляторным и интегративным центром. Он состоит из двух зрительных долей и является наиболее крупным отделом мозга. Такой тип мозга, где высшим регуляторным центром является средний мозг, называется ихтиопсидпым.
Промежуточный мозг состоит из крыши (таламуса) и дна (гипоталамуса) С гипоталамусом связан гипофиз, а с таламусом - эпифиз.
Мозжечок у рыб хорошо развит, поскольку их движения отличаются большим разнообразием.
Продолговатый мозг без резкой границы переходит в спинной мозг и в нем сосредоточен пищевой, сосудодвигательный и дыхательный центры.
От мозга отходит 10 пар черепно-мозговых нервов, что характерно для низших позвоночных.
|
|
|






