Дополнительная моторная область имеет, однако, иную топографическую организацию для контроля двигательной функции. Эта область лежит главным образом в продольной щели большого мозга, но занимает также несколько сантиметров верхней поверхности лобной коры. Сокращения, вызываемые стимуляцией этой области, — часто не односторонние, а двусторонние. Например, стимуляция ведет к одновременному сжиманию кистей обеих рук; такие движения, вероятно, являются рудиментарной функцией кистей, необходимой для лазания.
В целом эта область функционирует совместно с премоторной корой для обеспечения общей двигательной активности, необходимой для поддержания равновесия, фиксации определенных сегментов тела, позных движений головы и глаз и других, лежащих в основе более тонкого двигательного контроля рук и кистей премоторной областью и первичной моторной корой.
Нейрохирурги обнаружили у человека несколько высокоспециализированных моторных областей коры большого мозга, которые контролируют специфические двигательные функции. Локализация этих регионов была установлена либо путем электрической стимуляции, либо на основании данных о выпадении моторных функций при деструктивных поражениях определенных корковых областей. Некоторые из наиболее важных регионов следующие.
Область Брока и речь. На рисунке показана премоторная область, обозначенная «формирование
Первичная моторная кора (ПМК) занимает переднюю центральную извилину (4 поле по Brodmann). Расположенные в V слое ПМК гигантские пирамидные нейроны Беца являются главным источником кортико-спинальных волокон, которые непосредственно контактируют с альфа-мотонейронами передних рогов и являются основой пирамидного (двигательного) тракта.
Благодаря прямым связям со спинальными мотонейронами первичной моторной коры обладает наиболее низким порогом индукции движений при электрической или магнитной стимуляции.
Карты первичных двигательных областей коры больших полушарий вначале основывались на данных экспериментов, в которых в ответ на ее электрическое раздражение регистрировались дискретные движения на противоположной (контралатеральной) стороне тела ПЕРвичная кора - раздражение - низкий порог вызывают движения мышц. Однако оказалось, что если повысить интенсивность стимулов, движения можно вызвать при раздражении и других областей коры. На основании этих и многих других результатов (наблюдения за нарушениями при экспериментальных повреждениях мозга у животных; регистрация электрической активности мозга; современные визуализирующие исследования мозга человека) в коре больших полушарий выявлено несколько двигательных ("моторных") областей (рис. 40.1). Это первичная двигательная (моторная) кора прецентральной извилины, прилегающая к ней рострально премоторная область, вторичная соматосенсорная кора на крыше латеральной щели (область SII) и дополнительная двигательная область на медиальной стороне полушария. Часть премоторной области, примыкающая рострально к двигательному представительству лица, принадлежит лобным глазодвигательным полям.
Стимулы, нанесенные на поверхность первичной двигательной коры, приводят к дискретным движениям групп мышц на противоположной (контралатеральной) стороне. При микростимуляции (через микроэлектрод) можно наблюдать и сокращения индивидуальных мышц. Картирование с помощью микростимуляции показало, что двигательная кора - это мозаика двигательных точек, относящихся к определенным мышцам или группам мышц. Эти точки называются корковыми эфферентными зонами и соответствуют двигательным колонкам. Они организованы соматотопически и в совокупности составляют двигательный гомункулус (рис. 39.5).
Стимуляция дополнительной двигательной области может вызывать вокализацию либо сложные позные движения, например, медленное перемещение контралатеральной руки вперед, назад, вверх. Оно сопровождается движением головы и глаз в сторону руки. Позные движения могут быть двусторонними. При стимуляции этой области двигательной коры возможны и ритмические движения. Однако результат может иметь и обратный характер: временная остановка движения или речи. После одностороннего удаления дополнительной двигательной области наблюдаются медленные движения контралатеральных конечностей и склонность к неестественным движениям контралатеральной кисти (форсированным хватательным движениям контралатеральной кисти). При раздражении премоторной коры движения возникают только в случае высокой интенсивности стимулов.
Стимуляция лобных глазодвигательных полей одного из полушарий сопровождается содружественными саккадическими движениями глаз в противоположном направлении. Вертикальные саккады появляются только при двусторонней стимуляции лобных глазодвигательных полей. Одностороннее удаление лобного глазодвигательного поля ослабляет фиксацию взгляда на противоположной стороне; у человека глаза часто отклоняются в сторону повреждения. Исчезают саккады, связанные с воспоминаниями, но сохраняются саккады, возникшие в ответ на зрительные стимулы. При двустороннем повреждении лобных глазодвигательных полей и верхних бугорков четверохолмия прекращаются любые саккадические движения глаз.
Ссылки:
1. Агранулярн ая кора больших полушарий головного мозга
2. Активность индивидуальных кортико-спинальных нейронов
3. Ассоциативная кора
4. Двигательные петли головного мозга
5. Двигательный гомункулус
6. Доминирование полушария и речь
7. КОРА БОЛЬШИХ ПОЛУШАРИЙ ГОЛОВНОГО МОЗГА И УПРАВЛЕНИЕ ДВИЖЕНИЯМИ
8. Кора больших полушарий и нервная деятельность: основные положения
9. Кортикоцеребеллум (полушария мозжечка)
10. Общие сведения о роли мозжечка в двигательном контроле
11. Пирамидные клетки Беца
12. Подразделение стриатума на стриосомы и матрикс
13. Проекции глубинных ядер мозжечка: кортикоцеребеллум
14. Различия между двигательными петлями базальных ганглиев и мозжечка
15. Региональные различия структуры неокортекса
16. Рис. 39.4(fis). Области окончаний кортико-спинальных трактов
17. Рис. 40.2(fis). Двигательная кора и подкорковые структуры
18. Роль базальных ганглиев в двигательной регуляции
19. Роль дополнительной двигательной области в формировании команд
20. Сенсорная обратная связь кортико-спинальных нейронов
21. Стриосома стриатума
22. Структуры головного мозга в управлении движениями: основные положения
23. Центробежная регуляция соматовисцеральной чувствительности
24. ЭЭГ при эпилепсии
Каждая мышечная группа тела человека имеет пространственное представительство в строго определенной области прецентральной извилины коры головного мозга. Такой принцип получил название соматотопическая организация коры больших полушарий -моторный гомункулус (рис. 32).
 В связи с вышеизложенным выскажите свое мнение по поводу наличия/отсутствия (зачеркните неправильное) сенсорного гомункулуса. Рассмотрите рисунке 30 и определите местонахождение сенсорного гомункулуса: позиция 1 или 2 на рисунке 30 (неправильное зачеркните).
В связи с вышеизложенным выскажите свое мнение по поводу наличия/отсутствия (зачеркните неправильное) сенсорного гомункулуса. Рассмотрите рисунке 30 и определите местонахождение сенсорного гомункулуса: позиция 1 или 2 на рисунке 30 (неправильное зачеркните).
 Моторные зоны коры больших полушарий располагаются в разных отделах коры больших полушарий.
Моторные зоны коры больших полушарий располагаются в разных отделах коры больших полушарий.
Различают:
- основную (первичную) моторную зону, расположенную у человека в прецентральной извилине;
- дополнительную моторную зону, расположенную на медиальной поверхности коры;
- премоторную зоны коры, расположенную перед моторной зоной, ответственная за тонус мышц и осуществляющую координированные движения головы и туловища.
Отдельно рассказать о том, что замыслы движений зарождаются не в мотроной коре, подробно рассмотреть замысел, в чем произвольность движения заключается
Двигательные зоны управления произвольными движениями у человека находятся в передней/задней (зачеркните неправильное) центральных извилинах коры. Первичная моторная кора обозначена на рисунке 30 цифрой 1 или 2 (зачеркните неправильное).
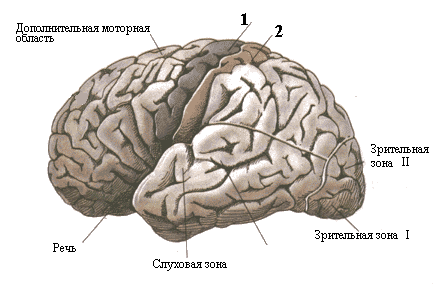
Рис. 30. Проекционные зоны коры.
 Стопа и колено
Стопа и колено
 Тело и локоть
Тело и локоть
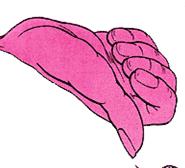 Кисть и пальцы
Кисть и пальцы

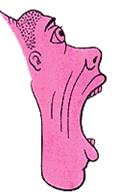 Шея, лицо, губы и челюсть
Шея, лицо, губы и челюсть
 Язык
Язык
Рис. 31. Части тела человека.
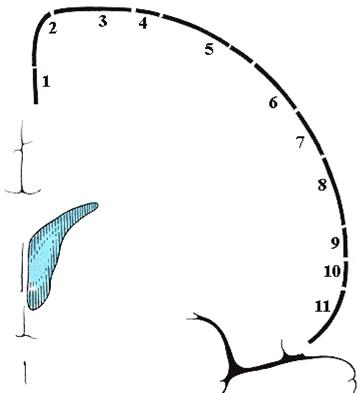
Рис. 32. Соматотопическая организация двигательной коры человека.
На рисунке 31 изображены части тела человека, а на рисунке 32 оцифровано их топографическое распределение в двигательной коре. Проставьте на рисунке 31 цифры, соответствующие распределению их на коре (рис. 32).
Нейроны в двигательной коре, так же как и в сенсорной зоне организованы в вертикальные диаметром , которые являются функциональными единицами коры. Все нейроны вертикально ориентированной ____________________ отвечают на одно и тоже афферентное раздражение одинаковой реакцией и совместно формируют эфферентный ответ, стимулируя группу мышц.
 В 1870 г. немецкие исследователи Фрич и Гитциг методом электрической стимуляции мозга собак впервые определили границы моторной коры. По современным данным, первичная моторная кора, управляющая движениями туловища и конечностей, соответствует полям 4 и 6 (по Бродману) (см. приложение 1). Ростральнее находятся корковые поля, управляющие движениями глаз (поля 8, 10) и речи (поля 44, 46). Анатомическим признаком моторных областей коры является агранулярность (невыраженность IV слоя коры). Кроме первичной моторной коры, в двигательном контроле у высших млекопитающих участвуют также премоторная, дополнительная моторная области коры, а также поле 5 (ростральная теменная кора).
В 1870 г. немецкие исследователи Фрич и Гитциг методом электрической стимуляции мозга собак впервые определили границы моторной коры. По современным данным, первичная моторная кора, управляющая движениями туловища и конечностей, соответствует полям 4 и 6 (по Бродману) (см. приложение 1). Ростральнее находятся корковые поля, управляющие движениями глаз (поля 8, 10) и речи (поля 44, 46). Анатомическим признаком моторных областей коры является агранулярность (невыраженность IV слоя коры). Кроме первичной моторной коры, в двигательном контроле у высших млекопитающих участвуют также премоторная, дополнительная моторная области коры, а также поле 5 (ростральная теменная кора).
Нисходящие пути двигательного контроля млекопитающих в соответствии с организацией их спинальных механизмов принято делить на латеральные (кортико- и руброспинальные) и медиальные (вестибуло- и ретикулоспинальные) проводящие системы. Латеральные проводящие системы в филогенезе млекопитающих возникли позже медиальных и обслуживают преимущественно движения конечностей, в том числе связанные с локомоцией. В отличие от этого медиальные системы преимущественно имеют отношение к рефлексам позы и установки головы и тела в пространстве.
Кортикоспинальная система двигательного контроля. Пирамидный (кортикоспинальный) тракт у млекопитающих – филогенетически самый молодой двигательный тракт. Он образован аксонами крупных нейронов, тела которых находятся в пятом слое 4-го и 6-го полей коры. У обезьян большая часть волокон берет начало от 4-го (31%) и 6-го (29%) полей коры, но 40% начинаются от теменной коры. Точные данные для человека отсутствуют, но известно, что не менее 60% волокон пирамидного тракта берут начало от поля 4. У человека пирамидный тракт одной стороны содержит более 1 млн. волокон.
На спинальном уровне у млекопитающих описано два кортикоспинальных тракта: мощный латеральный пирамидный тракт, волокна которого полностью перекрещиваются на уровне первого шейного сегмента, и неперекрещивающийся вентральный пирамидный тракт. Морфологические исследования показали, что у кошек и обезьян вентральный тракт заканчивается преимущественно в грудных сегментах спинного мозга. Этот тракт изучен недостаточно. Волокна латерального пирамидного тракта прослеживаются до поясничных сегментов спинного мозга. Согласно данным электрофизиологии латеральный пирамидный тракт активирует нейроны серого вещества основания заднего рога спинного мозга, которые, в свою очередь, активируют мотонейроны вентрального рога. Только у животных с подвижными фалангами пальцев (приматы, крыса, енот и некоторые другие) волокна пирамидного тракта активируют эти мотонейроны непосредственно.
Участие премоторных и теменных полей коры в двигательном контроле. Корковый уровень двигательного контроля представлен также премоторными полями коры и дополнительным моторным полем, а также полем 5 теменной коры, о функции которого пока известно очень мало.
Дополнительная моторная кора (ДМК). У приматов ДМК расположена на медиальной поверхности полушария (см. рис. 5.21). Подобно первичной моторной коре ДМК организована соматотопически. Движения, вызванные электрической стимуляцией ДМК, по сравнению со стимуляцией моторной коры выглядят более сложными и растянутыми во времени. Такие движения часто похожи на целенаправленные действия, в ряде случаев они развиваются билатерально на обеих конечностях и часто длятся больше времени электрической стимуляции ДМК.
При электрической стимуляции ДМК мозга человека во время нейрохирургических операций часто наблюдали вокализацию, которая могла сопровождаться гримасами лица, координированными движениями конечностей и торможением произвольных движений. Вентральный кортико-спинальный тракт и значительная часть кортико-бульбарного тракта могут рассматриваться как медиальная система путей. Эти тракты оканчиваются на интернейронах медиальной группы в спинном мозге, а также на нейронах соответствующей медиальной группы в стволе мозга и опосредуют управление осевыми мышцами. Эти мышцы часто сокращаются билатерально, обеспечивая позу и другие двусторонние функции, например, жевание или сдвигание бровей.
Другие пути медиальной системы начинаются в стволе мозга. Это латеральный вестибулоспинальный тракт и медиальный вестибулоспинальный тракт, ретикулоспинальный тракт моста и ретикулоспинальный тракт продолговатого мозга, тектоспинальный тракт.
Экстирпация ДМК у человека вызывает временную потерю речи (афазию), которая обычно проходит через несколько недель. Отмечают также замедление выполнения ритмических движений. Удаление ДМК у низших обезьян вызывает признаки нарушения целенаправленных движений (апраксию), нарушается также координация движений обеих рук. Вследствие этого у обезьян резко нарушается выполнение бимануальных задач, в которых необходима координация обеих рук. Нейронная активность ДМК коррелирует с характером движения, но установить связь с отдельными параметрами самого движения не удается. Предполагают, что нейронные сети ДМК принимают непосредственное участие в обработке сенсорной информации и в подготовке к движению (препрограммирование). В пользу этого говорит также усиление локального кровотока в ДМК у человека при намерении выполнить движение.
Участие в программировании движений премоторных полей коры.
Поражение премоторных зон у человека не сопровождается параличом контралатеральных конечностей. Вместе с тем их основным симптомом является отчетливое нарушение двигательных навыков. Клинически это проявляется в том, что у больного изменяется почерк, машинистка теряет быстроту и плавность своей работы, квалифицированный рабочий не способен автоматизированно выполнять серию операций, входящих в привычный двигательный акт. В клинике такие симптомы описывают под названием «инертность двигательных стереотипов». Характерными являются невозможность больного переключаться, например, с сильных ударов на слабые при отстукивании ритма ладонью, а также своеобразное «заклинивание» движения, двигательная персеверация (рис. 5.23). Наиболее отчетливо нарушения проявляются в контралатеральной очагу руке, но при поражении левого (доминантного) полушария они часто проявляются в работе обеих рук. Клиницисты отмечают, что у таких больных сохранены как намерения выполнить движение, так и общий план его выполнения, однако в своей исполнительной части движение высвобождается из-под влияния двигательной программы.

В тех случаях, когда поражение располагается в нижних отделах премоторной зоны левого (доминантного по речи у правшей) полушария, происходит нарушение речи. Нарушения проявляются при переходе больного от одной артикуляции к другой (при произнесении любого слова или словосочетания). Нарушение кинетической организации артикуляции является основой той формы моторной афазии, которую описал П. Брока (1861). Следует отметить, что нарушения у больных с поражениями нижних отделов левой премоторной зоны проявляются не только в устной речи, но и в письме. Нарушения письменной речи считают следствием нарушения плавности перехода от одного компонента слова к другому и патологической персеверации однажды написанного слова. Некоторые дополнительные сведения Вы найдете в главе 7 «Высшие функции нервной системы».
Регистрация активности отдельных нейронов показала, что нейроны премоторной коры низших обезьян активируются до выполнения движения, что может указывать на их связь с программированием этого движения. При этом в структуре нейронной реакции находит отражение направление, в котором будет производиться данное движение (в ответ, например, на условный сигнал). Изучение корреляции нейронных характеристик с параметрами движений рук у обезьяны приводит исследователей к мысли, что в премоторных нейронных сетях содержится своеобразная «библиотека» отдельных целенаправленных движений. Например, если обезьяна подносит рукой ко рту кусочек пищи, то рисунок нейронного разряда не зависит от траектории этого движения (например, из разных точек экстраперсонального пространства). Предполагают, что премоторная кора у приматов участвует в программировании целенаправленных движений (рук) в экстраперсональном пространстве (в пределах вытянутой руки).
Участие в двигательном контроле теменной коры. По современным данным, поле 5 теменной коры также принимает участие как в сенсорных, так и в моторных процессах. Поле 5 часто рассматривают как верхний этаж обработки кинестетической информации. Показано, что при локальном обратимом выключении (например, охлаждением) поля 5 у обезьян нарушались условные двигательные рефлексы, выполнение которых существенно зависело от сохранности проприоцептивного контроля, а при выключении поля 7 страдала также и зрительная обратная связь. У кошек глубокий распад условных двигательных рефлексов на зрительные сигналы происходил только при совместном разрушении полей 5 и 7, тогда как при изолированном разрушении каждого из теменных полей условные двигательные рефлексы заметно не страдали.
Данные, полученные при исследовании нейронных сетей, указывают, что нейроны моторной коры кодируют движение в координатах динамики движения и имеют дирекциональную), активность определяется направлением движения конечности) настройку. В отличие от этого нейронные сети поля 5 кодируют пространственную траекторию движения, и эти нейроны лишь незначительно модулируют свою активность при воздействии нагрузок на движущуюся руку. Отсюда был сделан вывод, что нейронные сети моторной коры кодируют движение в координатах его динамики, тогда как нейроны теменной коры кодируют пространственную траекторию движения.
В управлении сложными движениями мышц конечностей человека наиболее важна роль латерального кортико-спинального тракта (рис. 39.3).
Вентральный кортико-спинальный тракт относится к медиальной двигательной системе.
Латеральный кортико-спинальный тракт и кортико-бульбарный тракт начинаются в обширной части коры мозга, охватывающей двигательную область коры мозга, премоторную область коры мозга и дополнительную двигательную область коры мозга, а также соматосенсорную кору. В этих трактах участвуют в том числе аксоны крупных и мелких пирамидных клеток V слоя коры, а также гигантских пирамидных клеток Беца. Клетки Беца определяют гистологическую картину двигательной коры.
В самой каудальной области продолговатого мозга примерно 80% аксонов переходят на противоположную сторону. Затем в составе заднего бокового (латерального) канатика они спускаются в виде латерального кортико-спинального тракта. Неперекрещенная группа аксонов продолжает идти каудально в составе переднего (вентрального) канатика в виде переднего (вентрального) кортико-спинального тракта.
Кортико-бульбарный тракт оканчивается в стволе мозга у соответствующих ядер черепных нервов. Часть кортико-бульбарного пути оканчивается контралатерально (на другой стороне мозга) в ядре лицевого нерва на мотонейронах, снабжающих нижнюю часть лица, а также в ядре подъязычного нерва. Этот компонент организован так же, как латеральный кортико-спинальный тракт. Аксоны остальной части кортико- бульбарного тракта образуют окончания на обеих сторонах мозга, подобно аксонам вентрального кортико-спинального тракта.
Пирамидный, или кортико-спинальный путь начинается от больших пирамидных клеток (Беца) коры верхних 2/3 предцентральной извилины и околоцентральной дольки, проходит через внутреннюю капсулу основание ножек мозга, основание моста, пирамиды продолговатого мозга. На границе со спинным мозгом он разделяется на боковой и передний пирамидные пучки. Боковой (большой) образует перекрест и спускается в боковом канатике спинного мозга, заканчиваясь на клетках переднего рога. Передний не перекрещивается и идет в переднем канатике. Образуя посегментный перекрест, его волокна также заканчиваются на клетках переднего рога. Отростки клеток переднею рога образуют передний корешок, двигательную порцию спинномозгового нерва и заканчиваются в мышце двигательным окончанием.
Кортико-ядерный путь начинается в нижней трети предцентральной извилины, идет через колено (изгиб) внутренней капсулы и заканчивается на клетках двигательных ядер черепных нервов противоположной стороны. Отростки клеток двигательных ядер образуют двигательную порцию соответствующего нерва.
К рефлекторным двигательным путям (экстрапирамидным) относятся красноядерно-спинномозговой (руброспинальный) путь - от клеток красного ядра среднего мозга, тектоспиналъный путь - от ядер холмиков пластинки крыши среднего мозга (четверохолмия), связанный со слуховыми и зрительными восприятиями, и вестибуло-спинальный - от вестибулярных ядер из ромбовидной ямки, связанный с поддержанием равновесия тела.
Двигательные команды в виде потенциалов действия передаются непосредственно от коры больших полушарий к сегментам спинного мозга. Путь с помощью наиболее важного моторного пути, который называется кортикоспинальный тракт или пирамидный (см. рис. 33), поскольку он начинается от гигантских пирамидных клеток коры. Примерно 30% волокон кортикоспинального тракта исходят из первичной моторной коры, 30% - от премоторной и дополнительной моторной коры, а 40% - от соматосенсорных областей, находящихся позади центральной борозды. Аксоны пирамидных нейронов спускаются вниз и проходят через заднюю ножку внутренней капсулы (между хвостатым ядром и скорлупой базальных ганглиев) и следуют далее через ствол мозга, образуя пирамиды продолговатого мозга. Пирамидный путь состоит из двух отделов: первый - корково-ядерный – начинается от указанных выше нейронов моторных областей и заканчивается на ядрах черепных нервов в стволе мозга (рис. 33). Функция этого отдела пирамидного пути – регуляция произвольных движений мышц лица, глотки, языка, гортани, причем иннервация всех мышц лица, глотки и гортани происходит из обоих полушарий, кроме мышц языка и нижней части лицевой мускулатуры. Второй отдел – корковоспинальный – начинается от пирамидных клеток и формирует синапсы на вставочных (основная часть волокон) и α – мотонейронах передних рогов сегментов спинного мозга (рис. 33). 85% этого пути переходит на противоположную сторону. Аксоны альфа мотонейронов регулирует произвольные движения мышц рук на шейно-верхнегрудном уровне сегментов и мышцы ног – на пояснично-крестцовом уровне сегментов спинного мозга. 15% волокон этого отдела пирамидного пути не переходят на противоположную сторону и обеспечивают двухстороннюю иннервацию мышц туловища.
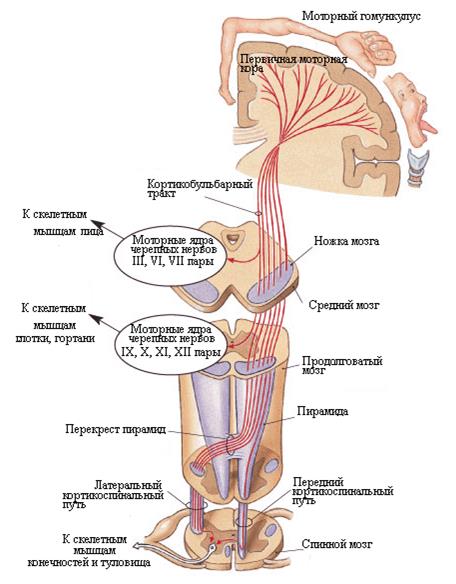
Рис.33. Кортикоспинальный (пирамидный) путь.





