ГОСУДАРСТВЕННОЕ БЮДЖЕТНОЕ ОБРАЗОВАТЕЛЬНОЕ УЧРЕЖДЕНИЕ ВЫСШЕГО ПРОФЕССИОНАЛЬНОГО ОБРАЗОВАНИЯ
«СТАВРОПОЛЬСКАЯ ГОСУДАРСТВЕННАЯ МЕДИЦИНСКАЯ АКАДЕМИЯ»
МИНИСТЕРСТВА ЗДРАВООХРАНЕНИЯ И СОЦИАЛЬНОГО РАЗВИТИЯ»
Российской федерации
КАФЕДРА БИОЛОГИИ
ХОДЖАЯН А.Б., ФЕДОРЕНКО Н.Н., ГЕВАНДОВА М.Г.
Структурно-функциональная организация клеток
Учебное пособие для студентов первого курса
Ставрополь, 2012
УДК:
ББК
С
Данное учебно-методическое пособие предназначено для студентов 1-го курса медицинского вуза.
Изд: СтГМА, Ставрополь, 2012., с..
Составители: д.м.н., проф.Ходжаян А.Б., к.м.н., доц.Федоренко НН., к.м.н., доц.Гевандова М.Г.
В учебно-методическом пособии «Структурно-функциональная организация клеток» изложены сведения о современных представлениях о строении и функциях прокариотических и эукариотических клеток, их эволюции и формах деления.
УДК:
ББК
С
Рецензент:
Рекомендовано к печати редакционно-издательским советом СтГМА
© Ставропольская государственная
медицинская академия, 2012
Все живые организмы состоят из клеток. Сама же история учения о клетке началась с создания микроскопа и связана с исследованиями английского ученого Роберта Гука, который впервые применил микроскоп для изучения биологических объектов. В 1665 году, рассматривая поперечные срезы стебля бузины и кору пробкового дерева (пробку), он пришел к заключению, что они состоят из мелких ячеек, напоминающих соты, и назвал их клетками (cells). Голландец, оптик-любитель, Антонио Левенгук (1685) обнаружил в болотной лозе простейшие одноклеточные организмы. Позже многие исследователи описывают микроскопическое строение в начале растений, а затем и животных. Если на первых порах в клетке видят только оболочку, то в 1825 году чешский ученый Ян Пуркинье, замечает, что клетка заполнена полужидким веществом, которое он назвал протоплазмой, и обнаруживает в нем клеточное ядро. В 1831 году клеточное ядро описывает и английский ботаник Б. Броун. К 1839 году немецкий ботаник М. Шлейден и немецкий зоолог Т. Шванн формулируют первую клеточную теорию, обобщившую наблюдения предшествовавших исследователей. Согласно этой теории, клетки являются простейшими элементарными структурами, из которых построены и развиваются ткани животных и растений. Создание клеточной теории стало важнейшим событием в биологии, определив универсальное значение клетки в построении живой природы. В то же время она поставила перед учеными ряд вопросов. Это вопрос о более детальном строении и развитии клеток, их происхождении, взаимоотношениях части и целого в организме, эволюции клеточных структур и др.
 В начале второй половины
В начале второй половины
XIX века
Рис.1. Портрет Р. Вирхова (1821-1902)
немецкий патолог Рудольф Вирхов в своем труде «Целлюлярная патология» выдвинул положение о том, что каждая клетка образуется от клетки, и связал патологические процессы организма с клеточными изменениями.
В последней четверти XIX века И.Д.Чистяковым (1843-1877), П.И. Перемежко (1833-1893), В.Флемингом (1843-1906) Э. Страсбургером (1844-1912) было открыто непрямое деление клеток (митоз) и были описаны многие органоиды клеток.
В 1934 году немецкий ученый Е. Руска, сконструировал электронный микроскоп и начинается период электронно-микроскопических исследований клетки, в ходе которых уточняется тонкая организация уже изученных клеточных структур и открываются ранее неизученные клеточные структуры.
Несмотря на новые открытия основные положения первой клеточной теории сохранили свое значение, однако были получены некоторые дополнения. Современная клеточная теория основывается на следующих основных постулатах:
1) Клетка является основной структурно-функциональной единицей всего живого;
2) Клетки различных организмов сходны по строению и основным процессам жизнедеятельности;
3) Клетки имеют мембранный принцип строения;
4) Новые клетки образуются только путем деления материнских;
5) Многоклеточный живой организм представляет собой сложноорганизованную, целостную интегрированную систему взаимодействующих клеток.
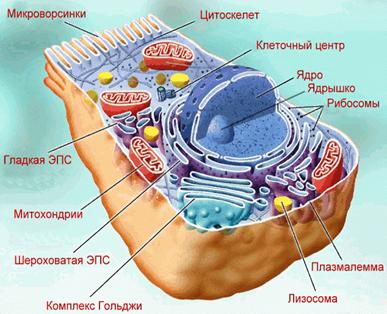
Рис.2. Схема электронно-микроскопической организации
эукариотической клетки.
Эволюция клетки
Все существующие в настоящее время клетки подразделяются на два типа: прокариоты и эукариоты. Эволюционно наиболее ранними являются прокариоты, которые появились около 3- 3,5млрд. лет назад, когда в ходе химической эволюции в мировом океане появились полипептиды и полинуклеотиды, из которых последние обладали матричными свойствами. Первыми нуклеотидами, видимо, были молекулы РНК, которые примитивным образом направляли первичный синтез белков.
Одним из решающих событий, приведших к формированию первой клетки, стало образование внешней мембраны за счет появившихся к тому времени амфипатрических молекул фосфолипидов и белков.
Первые клетки на Земле, по-видимому, имели некоторое сходство с микоплазмой. Современные микоплазмы – это похожие на бактерии мелкие по размерам организмы, обычно ведущие паразитический образ жизни у растений и животных. Некоторые из микоплазм, имея размеры 0,3 мкм, содержат РНК, достаточную для синтеза 750 различных белков, необходимых для жизни клетки.
По существующим в настоящее время представлениям, именно молекулы РНК первыми включились в эволюцию. ДНК стала необходимой только тогда, когда клетки сильно усложнились, и у них возникла необходимость в значительно большей генетической информации, чем та, которую могла стабильно поддерживать РНК. В роли хранителя наследственной информации вместо РНК стала ДНК.
Относительно механизмов образования эукариотических клеток существуют две гипотезы: инвагинационная и симбиотическая. Согласно инвагинационной гипотезе, ядро и мембранные органоиды сформировались за счет впячивания (инвагинаций) плазматической мембраны древней прокариотической клетки, содержащей несколько геномов, прикрепленных к клеточной мембране. В дальнейшем произошла специализация, и органеллы, содержащие элементы генома, трансформировались в ядро, митохондрии, пластиды, а не содержащие его – в другие органоиды мембранного происхождения.
Наиболее распространенной в настоящее время является симбиотическая гипотеза, согласно которой эукариотические клетки произошли вследствие объединения крупных анаэробных прокариотических клеток, живущих за счет брожения, с другими, более мелкими анаэробными бактериями и анаэробными фотосинтезирующими одноклеточными водорослями (предками современных сине-зеленых водорослей). Таким образом, видимо, сформировались митохондрии и пластиды, характерные для эукариотических клеток. Что касается ядра, то полагают, что оно является рудиментом какого-то древнего симбионта, утратившего свою цитоплазму.
Эукариотические клетки
Эукариотические клетки появились около 1,5 млрд. лет назад. Им свойственна более сложная организация. Они имеют различную форму и размеры и образуют как одноклеточные, так и многоклеточные организмы. Различают животные и растительные эукариотические клетки. Они имеют общий принцип строения, хотя им присущи и некоторые различия.
В типичной эукариотической клетке имеются: ядро, цитоплазма и оболочка.

Рис.11. Строение животной клетки.
Биологические мембраны
Характерной чертой строения растительных и животных клеток является мембранный принцип. Биологические мембраны входят в состав клеточной оболочки многих органоидов и оболочки ядра. Они имеют общую организацию и выполняют ряд важнейших функций.
Мембраны имеют толщину около 5-10 нм и по химической организации являются липопротеидными структурами. Их основу составляет липидный бислой, состоящий преимущественно из фосфолипидов.

Рис.12. Строение биологических мембран.
1 — молекула липида;
2 — липидный бислой;
Интегральные белки;
Периферические белки;
Полуинтегральные белки;
Гликокаликс;
Субмембранный слой;
Актиновые микрофиламенты;
Микротрубочки;
Промежуточные филаменты;
Кроме последних, в мембранах могут присутствовать гликолипиды и холестерол. Все они обладают свойством амфипатричности, т.е. у них есть гидрофильный («любящий воду») и гидрофобный («боящийся воды») концы. Первые обращены кнаружи от бислоя в виде двух противоположных рядов полярных «головок», а вторые – внутрь друг к другу и имеют вид разветвленных неполярных «хвостов». Большинство фосфолипидов и гликолипидов в водной среде самопроизвольно образуют бислой и смыкаются свободными концами, что лежит в основе восстановления структуры мембран при повреждении.
Липидный бислой может находиться в состоянии жидкого и твердого кристалла. Молекулы липидов высокоподвижны и способны перемещаться в пределах своего монослоя и очень редко из одного монослоя в другой.
Для липидного бислоя характерна асимметричность: его монослои отличаются друг от друга по составу липидов. Различный липидный состав отличают также разные мембраны одной и той же эукариотической клетки.
Вторым обязательным компонентом биологических мембран являются белки. Если липидный бислой определяет основные структурные особенности мембран, белки обеспечивают большинство мембранных функций. Они выступают в качестве специфических рецепторов, несут ферментативную функцию, а также осуществляют транспорт через мембрану различных веществ. Многие мембранные белки способны вращаться вокруг оси, перпендикулярной плоскости бислоя или перемещаться в плоскости самой мембраны, однако они не могут перескакивать с одной стороны бислоя на другую. Это контролируемый процесс, связанный различного рода взаимодействиями между белками мембраны и различными элементами цитоплазмы.
В 1935 году английские ученые Л. Даниэлли и Г. Даусон высказали идею о «бутербродной» модели биологических мембран, в соответствии с которой основу мембран составляет липидный бислой, покрытый сверху и снизу сплошным слоем белков.
В 1972 году М. Сингер и Дж. Николсон описывают жидкостно-мозаичную модель мембраны, получившую широкое признание. Согласно этой модели белки не образуют сплошных слоев поверх липидов бислоя, а вкраплены в него в виде мозаики: некоторые белки располагаются группами на поверхности бислоя (периферические белки), другие – пронизывают бислой полностью или частично (интегральные и полуинтегральные трансмембранные белки).
Молекулы белков, как и липидов, обладают амфипатрическими свойствами: у них есть гидрофобные участки, взаимодействующие с гидрофобными хвостами липидных молекул внутри бислоя, и гидрофильные участки, обращенные к водной среде и образующие с молекулами воды водородные связи.
Позже появились сведения о том, что часть белков, входящих в состав биологических мембран, образует в липидном бислое жесткий каркас, который сохраняется после вымывания липидов жирорастворяющими веществами. Этот вид модели получил название белково-кристаллической.
Описанные три вида моделей биологических мембран не исключают друг друга. Они могут встречаться в разных мембранах или же в разных участках одной и той же мембраны в зависимости от функциональных особенностей данного участка.
Интегральные белки;
Периферические белки;
Полуинтегральные белки;
Гликокаликс;
Субмембранный слой.
У растительных клеток поверх наружной клеточной мембраны располагается плотная целлюлозная стенка с порами, через которые осуществляется связь между соседними клетками посредством цитоплазматических мостиков.
Свойства биологических мембран:
1. Способность к самосборке после разрушающих воздействий. Это свойство определяется физико-химическими особенностями фосфолипидных молекул, которые в водном растворе собираются вместе так, что гидрофильные концы молекул разворачиваются наружу, а гидрофобные - внутрь. В уже готовые фосфолипидные слои могут встраиваться белки. Способность к самосборке имеет важное значение на клеточном уровне.
2. Полупроницаемость (избирательность в пропускании ионов и молекул). Обеспечивает поддержание постоянства ионного и молекулярного состава в клетке.
3. Текучесть мембран. Мембраны не являются жесткими структурами, они постоянно флюктуируют за счет вращательных и колебательных движений молекул липидов и белков. Это обеспечивает большую скорость протекания ферментативных и других химических процессов в мембранах.
4. Фрагменты мембран не имеют свободных концов, так как замыкаются в пузырьки.
Функции наружной клеточной мембраны (плазмолеммы):
Основными функциями плазмолеммы являются следующие: 1) барьерная, 2) рецепторная, 3) обменная, 4)транспортная.
1. Барьерная функция. Она выражается в том, что плазмолемма ограничивает содержимое клетки, отделяя его от внешней среды, а внутриклеточные мембраны разделяют цитоплазму на отдельные реакционные отсеки-компартменты.
2. Рецепторная функция. Одной из важнейших функций плазмолеммы является обеспечение коммуникации (связи) клетки с внешней средой посредством присутствующего в мембранах рецепторного аппарата, имеющего белковую или гликопротеиновую природу. Основная функция рецепторных образований -плазмалеммы – внешние сигналы, благодаря которым клетки правильно ориентируются и образуют ткани в процессе дифференцировки. С рецепторной функцией связана деятельность различных регуляторных систем, а также формирование иммунного ответа.
3. Обменная функция определяется содержанием в биологических мембранах ферментных белков, являющихся биологическими катализаторами. Их активность меняется в зависимости от рН среды, температуры, давления, концентрации как субстрата, так и самого фермента. Ферменты определяют интенсивность ключевых реакций метаболизма, а также их направленность.
4. Транспортная функция мембран. Мембрана обеспечивает избирательное проникновение в клетку и из клетки в окружающую среду различных химических веществ. Транспорт веществ необходим для поддержания в клетке соответствующего рН, надлежащей ионной концентрации, что обеспечивает эффективность работы клеточных ферментов. Транспорт поставляет питательные вещества, которые служат источником энергии, а также материалом для образования различных клеточных компонентов. От него зависят выведение из клетки токсических отходов, секреция различных полезных веществ и создание ионных градиентов, необходимых для нервной и мышечной активности. Изменение скорости переноса веществ может приводить к нарушениям биоэнергетических процессов, водно-солевого обмена, возбудимости и других процессов. Коррекция этих изменений лежит в основе действия многих лекарственных препаратов.
Существует два основных способа поступления веществ в клетку и вывода из клетки во внешнюю среду;
- пассивный транспорт,
- активный транспорт.
Пассивный транспорт идет по градиенту химической или электрохимической концентрации без затрат энергии АТФ. Если молекула транспортируемого вещества не имеет заряда, то направление пассивного транспорта определяется только разностью концентрации этого вещества по обеим сторонам мембраны (градиент химической концентрации). Если же молекула заряжена, то на ее транспорт влияет как градиент химической концентрации, так и электрический градиент (мембранный потенциал).
Оба градиента вместе составляют электрохимический градиент. Пассивный транспорт веществ может осуществляться двумя способами – простой диффузией и облегченной.
При простой диффузии ионы солей и вода могут проникать через селективные каналы. Эти каналы образуются за счет некоторых трансмембранных белков, формирующих сквозные транспортные пути, открытые постоянно или только на короткое время. Через селективные каналы проникают различные молекулы, имеющие соответствующие каналам размер и заряд.
Имеется и другой путь простой диффузии – это диффузия веществ через липидный бислой, через который легко проходят жирорастворимые вещества и вода. Липидный бислой непроницаем для заряженных молекул (ионов), и в то же время незаряженные малые молекулы могут свободно диффундировать. При этом, чем меньше молекула, тем быстрее она транспортируется. Довольно большая скорость диффузии воды через липидный бислой как раз и объясняется малой величиной ее молекул и отсутствием заряда.
При облегченной диффузии в транспорте веществ участвуют белки – переносчики, работающие по принципу «пинг-понг». Белок при этом существует в двух конформационных состояниях: в состоянии «понг» участки связывания транспортируемого вещества открыты с наружной стороны бислоя, а в состоянии «пинг» такие же участки открываются с другой стороны. Этот процесс обратимый. С какой же стороны в данный момент времени будет открыт участок связывания вещества, зависит от градиента концентрации. Таким способом через мембрану проходят сахара и аминокислоты.
При облегченной диффузии скорость транспортировки веществ значительно возрастает в сравнении с простой диффузией.
Кроме белков-переносчиков, в облегченной диффузии принимают участие некоторые антибиотики, например, грамицидин и валиномицин.
Поскольку они обеспечивают транспорт ионов, их называют ионофорами.
Активный транспорт веществ в клетке. Этот вид транспорта всегда идет с затратой энергии. Источником энергии, необходимой для активного транспорта, является АТФ. Характерной особенностью этого вида транспорта является то, что он осуществляется двумя способами:
1) с помощью ферментов, называемых АТФ-азами;
2) транспорт в мембранной упаковке (эндоцитоз).
В наружной клеточной мембране присутствуют такие белки-ферменты, как АТФ-азы, функция которых заключается в обеспечении активного транспорта ионов против градиента концентрации. Поскольку они обеспечивают транспорт ионов, то этот процесс называют ионным насосом.
Известны четыре основные системы транспорта ионов в животной клетке. Три из них обеспечивают перенос через биологические мембраны Na+ и К+, Са+, Н+, а четвертый – перенос протонов при работе дыхательной цепи митохондрий.
Примером механизма активного транспорта ионов может служить натрий-калиевый насос в животных клетках. Он поддерживает в клетке постоянную концентрацию ионов натрия и калия, которая отличается от концентрации этих веществ в окружающей среде: в норме в клетке ионов натрия бывает меньше, чем в окружающей среде, а калия – больше.
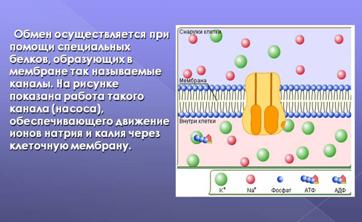
Рис.14. Схематическая модель калий -натриевого
насоса.
Вследствие этого по законам простой диффузии калий стремится уйти из клетки, а натрий диффундирует в клетку. В противовес простой диффузии натрий – калиевый насос постоянно выкачивает из клетки натрий и вводит калий: на три молекулы выбрасываемого наружу натрия приходится две молекулы вводимого в клетку калия.
Обеспечивает этот транспорт ионов натрий-калий-АТФ-аза – фермент, локализующийся в мембране таким образом, что пронизывает всю ее толщу. С внутренней стороны мембраны к этому ферменту поступают натрий и АТФ, а с наружной – калий.
Перенос натрия и калия через мембрану совершается в результате конформационных изменений, которые претерпевает натрий-калий-АТФ-аза, активизирующаяся при повышении концентрации натрия внутри клетки или калия в окружающей среде.
Для энергообеспечения этого насоса необходим гидролиз АТФ. Этот процесс обеспечивает все тот же фермент натрий-калий-АТФ-аза. При этом более одной трети АТФ, потребляемой животной клеткой в состоянии покоя, расходуется на работу натрий-калиевого насоса.
Нарушение правильной работы натрий-калиевого насоса приводит к различным серьезным заболеваниям.
КПД этого насоса превышает 50%, чего не достигают самые совершенные машины, созданные человеком.
Многие системы активного транспорта приводятся в действие за счет энергии, запасенной в ионных градиентах, а не путем прямого гидролиза АТФ. Все они работают как котранспортные системы (способствующие транспорту низкомолекулярных соединений). Например, активный транспорт некоторых сахаров и аминокислот внутрь животных клеток обусловливается градиентом иона натрия, причем чем выше градиент ионов натрия, тем больше скорость всасывания глюкозы. И, наоборот, если концентрация натрия в межклеточном пространстве заметно уменьшается, транспорт глюкозы останавливается. При этом натрий должен присоединиться к натрий-зависимому белку-переносчику глюкозы, который имеет два участка связывания: один для глюкозы, другой для натрия. Ионы натрия, проникающие в клетку, способствуют введению в клетку и белка-переносчика вместе с глюкозой. Ионы натрия, проникшие в клетку вместе с глюкозой, выкачиваются обратно натрий-калий-АТФ-азой, которая, поддерживая градиент концентрации натрия, косвенным путем контролирует транспорт глюкозы.
Цитоскелет
Цитоскелет – это трехмерный цитоплазматический комплекс волокнистых и трубчатых структур, придающий клетке определенную форму и выполняющий другие функции. Он представлен микротрубочками, микрофиламентами и промежуточными филаментами.
Микротрубочки – неветвящийся структурный компонент цитоплазмы, присутствующий практически во всех типах эукариотических клеток. Имеют диаметр 24 нм, а в длину –несколько мкм. Это динамические структуры, постоянно растущие с одного конца и деполяризующиеся с другого. Они построены из белка тубулина. Имеют вид полых цилиндров.
Их главная функция заключается в создании эластичного и устойчивого внутриклеточного каркаса, необходимого для поддержания формы клеток. Кроме того, они принимают участие в транспорте макромолекул и органоидов, обеспечивают подвижность жгутиков и ресничек. Входя в состав веретена деления, обеспечивают расхождение хромосом при делении клеток. Они могут быть факторами ориентированного движения клетки в целом.
Разрушение микротрубочек колхицином приводит к нарушениям транспорта веществ (например, к блокаде секреции) и механического переноса отдельных внутриклеточных компонентов. Кроме того, при делении клеток блокируется веретено деления.
Микрофиламенты – относятся к фибриллярным компонентам эукариотических клеток. Они располагаются в кортикальном слое цитоплазмы, прямо под плазмолеммой, в виде пучков или слоями. В среднем их длина составляет 6 нм.
В зависимости от химического состава микрофиламенты могут выполнять функции цитоскелета и участвовать в обеспечении движения. В их состав входят сократительные белки: актин, миозин, тропомиозин и др.
Основные функции микрофиламентов – это создание внутриклеточного сократительного аппарата, который обеспечивает амебоидное движение клеток, большинство движений органоидов и деление клеток.
Промежуточные филаменты (микрофибриллы) – это тонкие неветвящиеся, нередко лежащие пучками нити, которые присутствуют в клетках разных тканей и состоят из разных веществ, например, из кератина – в эпителии, десмина – в мышечных волокнах и др. Они выполняют опорно-каркасную функцию.
Включения
Включения – это непостоянные компоненты, которые могут появляться и исчезать в зависимости от функционального состояния клетки. Их классифицируют на: трофические, секреторные, экскреторные, пигментные, кристаллы. Трофические включения – это, как правило, резервные скопления гранул белка, гликогена, капельки жиров. Секреторные гранулы и пузырьки содержат биологически активные вещества и поступают по протокам желез в те или иные органы. Экскреторные включения не являются биологически активными веществами. Обычно это продукты метаболизма клеток, подлежащие удалению. Пигментные включения могут изменить цвет ткани временно или постоянно (примеры: гемоглобин, меланин, липофусцин – пигмент изнашивания, билирубин и др.). В растительных клетках встречаются включения кристаллов (чаще всего это оксалат кальция).
Строение и функции ядра
Ядро (лат. nucleus, греч. karion-ядро) – это обязательный компонент эукариотических клеток. Оно хорошо различимо в неделящихся клетках и выполняет ряд важнейших функций:
1) хранение и передача наследственной информации в клетке;
2) создание аппарата белкового синтеза – синтез всех видов РНК и образование рибосом.
Выпадение или нарушение любой из этих функций приводит клетку к гибели.

Рис.24. Схема ультрамикроскопического строения ядра.
Клетка содержит, как правило, одно ядро, но имеются двуядерные и многоядерные клетки.
Интерфазные ядра состоят из: ядерной оболочки, ядерного сока (кариоплазма, кариолимфа или нуклеоплазма), ядерного белкового остова, хроматина и ядрышек.
Ядерная оболочка (кариолемма) состоит из двух мембран, между которыми имеется перинуклеарное пространство шириной 10-40нм, заполненное электронно – микроскопически рыхлой субстанцией. Наружная мембрана ядерной оболочки со стороны цитоплазмы в ряде участков переходит в мембраны эндоплазматической сети, и на ее поверхности располагаются полирибосомы. Внутренняя мембрана ядерной оболочки участвует в обеспечении внутреннего порядка в ядре – в фиксации хромосом в трехмерном пространстве. Эта связь опосредуется с помощью слоя фибриллярных белков, сходных с промежуточными филаментами цитоплазмы.
В ядерной оболочке имеются поры диаметром около 90 нм. В этих участках по краям отверстия мембраны ядерной оболочки сливаются. Сами отверстия заполняются сложноорганизованными глобулярными и фибриллярными структурами. Совокупность мембранных перфораций и заполняющих их структур называется поровым комплексом.
По краю порового отверстия располагаются в три ряда гранулы (по 8 гранул в каждом ряду). При этом один ряд лежит со стороны цитоплазмы, другой – со стороны внутреннего содержимого ядра, а третий – между ними. От гранул этих слоев радиально отходят фибриллярные отростки, образуя в поре как бы перепонку – диафрагму. Фибриллярные отростки направляются к центрально расположенной грануле.
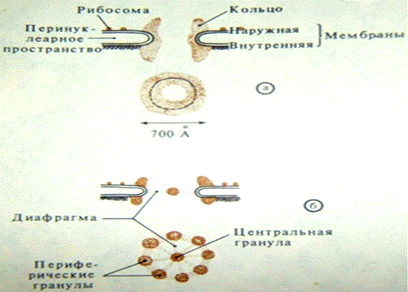
Рис.25. Строение ядерных пор (поровый комплекс).
Поровые комплексы участвуют в рецепции транспортируемых через поры макромолекул (белков и нуклеопротеидов), а также в активном переносе через ядерную оболочку этих веществ с использованием АТФ.
Число ядерных пор зависит от метаболической активности клеток. Чем интенсивнее протекают в клетке процессы синтеза, тем больше пор. В среднем на одно ядро приходится несколько тысяч поровых комплексов.
Основные функции ядерной оболочки следующие:
- барьерная (отделение содержимого ядра от цитоплазмы и ограничение свободного доступа в ядро крупных биополимеров);
- регуляция транспорта макромолекул между ядром и цитоплазмой;
- участие в создании внутриядерного порядка (фиксация хромосомного аппарата).
Кариоплазма (ядерный сок, или нуклеоплазма, или кариолимфа) – это содержимое ядра, которое имеет вид гелеобразного матрикса. Она содержит различные химические вещества: белки (в том числе и ферменты), аминокислоты и нуклеотиды в виде истинного или коллоидного раствора.
Ядерный или белковый остов (матрикс). В интерфазных ядрах негистоновые белки образуют сеть – «белковый матрикс». Он состоит из периферического фибриллярного слоя, выстилающего ядерную оболочку (ламина), и внутренней сети, к которой прикрепляются фибриллы хроматина. Матрикс участвует в поддержании формы ядра, организации пространственного положения хромосом. Кроме того, в нем содержатся ферменты, необходимые для синтеза РНК и ДНК, а также белки, участвующие в компактизации ДНК в интерфазных и митотических хромосомах.
Хроматин – комплекс ДНК и белков (гистоновых и негистоновых). Хроматин является интерфазной формой существования хромосом.

1.Эухроматин; 2. Гетерохроматин
Рис.26. Хроматин интерфазных хромосом.
В этот период разные участки хромосом имеют неодинаковую степень компактизации. Наибольшей степенью компактизации обладают генетически инертные участки хромосом. Они хорошо окрашиваются ядерными красителями и называются гетерохроматином. Различают конститутивный и факультативный гетерохроматин.
Конститутивный гетерохроматин образован нетранскрибируемой ДНК. Полагают, что он участвует в поддержании структуры ядра, прикреплении хромосом к ядерной оболочке, узнавании при мейозе гомологичных хромосом, разделении соседних структурных генов и в процессах регуляции их активности.
Факультативный гетерохроматин, в отличие от конститутивного, может становиться транскрибируемым на определенных стадиях клеточной дифференцировки или онтогенеза. Примером факультативного гетерохроматина может служить тельце Барра, образующееся у организмов гомогаметного пола за счет инактивации одной из Х-хромосом.
Декомпактизированные участки хромосом, которые плохо окрашиваются ядерными красителями, называются эухроматином. Это функционально активный, транскрибируемый хроматин.
Ядрышки – уплотненные тельца, обычно округлой формы, диметром менее 1 мкм. Присутствуют они только в интерфазных ядрах. Количество их колеблется в диплоидных клетках от 1 до 7, но в некоторых видах клеток, например, микронуклеусах инфузории, ядрышки отсутствуют.
Ядрышки содержат более 80% белка и около 15% р-РНК, а также ядрышковый хроматин. Ядрышки образуются в области вторичных перетяжек хромосом, представленных ядрышковыми организаторами. При транскрипции этих участков хромосом образуется рибосомальная РНК, которая, связываясь с белками, участвует в формировании малых и больших субъединиц рибосом.
Жизненный цикл клеток
В настоящее время считается бесспорным, что клетки возникают только в результате деления. При этом вновь возникшие клетки не сразу имеют все структурные компоненты, свойственные зрелым клеткам. Для созревания клеток должно пройти определенное время. Процесс созревания клеток называется дифференцировкой. В процессе дифференцировки клетки приобретают черты структурной и функциональной зрелости. Большинство клеток по истечении определенного времени приступает к делению (за исключением зрелых нейронов) или же погибает. Генетически запрограммированная гибель клеток называется апоптозом. В отличие от последней, гибель клеток в результате случайных внешних причин, например, травмы, ожога и т.д., называется некрозом.
Деление клеток лежит в основе роста организма и регенерации. Основным способом деления соматических клеток является митоз. Кроме него, встречаются и другие формы деления: амитоз и эндомитоз.
Промежуток времени между клеточными делениями называется интерфазой.
Некоторые цитологи выделяют два вида интерфаз: гетеросинтетическую и аутосинтетическую.
В период гетеросинтетеической интерфазы клетки работают на организм, выполняя свои функции составного компонента того или иного органа или такни. В период аутосинтетической интерфазы клетки готовятся к митозу или мейозу. В этой интерфазе выделяют три периода: пресинтетический – G1, синтетический – S, и постсинтетический – G2.
Период G1 – самая продолжительная фаза. В этот период клетка синтезирует РНК и белки.
В S-периоде продолжается синтез белка и происходит репликация ДНК. В большинстве клеток этот период длится 8-12 часов.
В G2 – периоде продолжается синтез РНК и белка (например, тубулина для построения микротрубочек веретена деления). Происходит накопление АТФ для энергетического обеспечения последующего митоза. Эта фаза длится 2-4- часа.
Кроме интерфазы, для характеристики временной организации клеток выделяют такие понятия, как жизненный цикл клеток, клеточный цикл и митотический цикл. Под жизненным циклом клетки понимают время жизни клетки с момента ее возникновения после деления материнской клетки и до конца ее собственного деления или же до гибели.
Клеточный цикл – это совокупность процессов, протекающих в аутосинтетическую интерфазу, и собственно митоз.
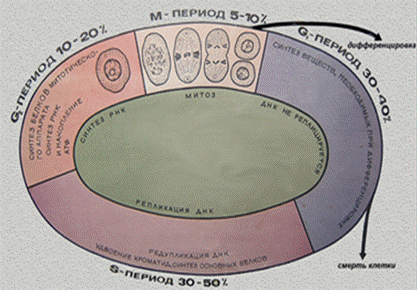
Рис.27. Клеточный цикл. Интерфаза.
Митотический цикл – это совокупность процессов, протекающих в клетке в течение собственно митоза.
МИТОЗ
Митоз (от греч. митос – нить), или кариокинез (греч. карион – ядро, кинезис – движение), или непрямое деление. Это процесс, в ходе которого происходит конденсация хромосом и равномерное распределение дочерних хромосом между дочерними клетками. Митоз включает в себя пять фаз: профаза, прометафаза, метафаза, анафаза и телофаза. В профазе хромосомы конденсируются (скручиваются), становятся заметными и располагаются в виде клубка. Центриоли делятся на две и начинают двигаться к клеточным полюсам. Между центриолями появляются нити, состоящие из белка тубулина. Происходит образование митотического веретена. В прометафазе ядерная оболочка распадается на мелкие фрагменты, а погруженные в цитоплазму хромосомы начинают двигаться к экватору клетки. В метафазе хромосомы устанавливаются на экваторе веретена и становятся максимально компактизированными. Каждая хромосома состоит из двух хроматид, связанных друг с другом центромерами, а концы хроматид расходятся, и хромосомы принимают Х-образную форму. В анафазе дочерние хромосомы (бывшие сестринские хроматиды) расходятся к противоположным полюсам. Предположение о том, что это обеспечивается сокращением нитей веретена, не подтвердилось.

Рис.28. Характеристика митоза и мейоза.
Многие исследователи поддерживают гипотезу скользящих нитей, согласно которой соседние микротрубочки веретена деления, взаимодействуя друг с другом и сократительными белками, тянут хромосомы к полюсам. В телофазе дочерние хромосомы достигают полюсов, деспирализуются, образуется ядерная оболочка, восстанавливается интерфазная структура ядер. Затем наступает разделение цитоплазмы – цитокинез. В животных клетках этот процесс проявляется в перетяжке цитоплазмы за счет втягивания плазмолеммы между двумя дочерними ядрами, а в растительных клетках мелкие пузырьки





