Предисловие
Ускоренное и устойчивое наращивание производства зерна, как это подчеркивается в Продовольственной программе СССР, является ключевой проблемой в сельском хозяйстве. Главный путь ее решения лежит через повышение урожайности зерновых культур при условии совершенствования агротехнической культуры возделывания. В этом плане важное значение приобретают мероприятия, направленные на резкое снижение потерь урожая, вызываемых возбудителями болезней зерновых культур.
Среди возбудителей болезней сельскохозяйственных растений головневые грибы занимают видное место. Их вредоносность подчас весьма значительна, в силу чего эта группа патогенов должна находиться под постоянным контролем со стороны практических работников сельского хозяйства.
В 40 - 50-е годы усилиями фитопатологов, в числе которых были А. И. Борггардт, Т. Д. Страхов, Н. А. Наумов, П. А. Пройда, П. Н. Давыдов, К. Я. Калашников, И. М. Поляков, была разработана и внедрена система мероприятий по борьбе с головней хлебных злаков. Эта система позволила обеспечить снижение недобора урожая от головневых болезней с 12 до 0.75 % и сохранила от гибели миллионы тонн зерна.
Результаты мер борьбы с головневыми заболеваниями оказались столь внушительными, что подчас создавалось впечатление о практически полной ликвидации этой группы заболеваний. Однако защита зерновых культур до сих пор остается общегосударственной задачей. В ряде мест страны отмечено усиление вредоносности головневых заболеваний. Причины этих явлений в основном биологического характера. Среди них - сдвиги в популяционном составе головневых грибов, возрастание однородности сортов, а также потеря устойчивости к головне у отдельных, широко культивируемых сортов. В борьбе с головневыми болезнями важное значение приобретает знание особенностей биологии их возбудителей. Головневые - одна из полно изученных во многих отношениях групп фитопатогенных грибов. Четкое знание разнообразных сторон биологии возбудителей головни, требующих к себе дифференцированного подхода, способствует разработке эффективных мероприятий по борьбе с ними.
В настоящее время в нашей стране известно около 20 видов возбудителей головни зерновых культур. Сведения о них часто разрозненны, что затрудняет их использование в практике сельского хозяйства. Цель настоящей книги - в некоторой степени систематизировать данные по биологии возбудителей. Часть этих сведений получена лишь в последнее время. Кроме возбудителей широко распространенных заболеваний, приведены виды, неизвестные пока в СССР, но которые представляют потенциальную опасность для зернового хозяйства. Всего представлены очерки о 25 видах. Внимание акцентировалось на вопросах, связанных с происхождением возбудителей, их специализацией, морфологией, циклами развития. Из мер борьбы рассмотрены лишь основные, наиболее результативные. Литература по головневым грибам обширна. Мы ограничивались лишь некоторыми, наиболее существенными, с нашей точки зрения, ссылками.
Возбудители головневых заболеваний подразделены нами на 5 неравных по числу видов групп в соответствии с 5 трибами, к которым относятся культивируемые хлебные злаки. Это возбудители головни пшеницы, ржи и ячменя (триба Triticeae), овса (триба Aveneae), риса (триба Oryzeae), кукурузы и сорго (триба Andropogoneae) и просяных культур (триба Paniceae). Основной части книги предшествует краткий очерк биологии головневых грибов.
Книга рассчитана на агрономов, специалистов по защите растений, научных работников - микологов, фитопатологов, селекционеров, а также на студентов и преподавателей сельскохозяйственных вузов и университетов.
Возбудители головни овса
Ustilago avenae (Pers.) Jens.- возбудитель пыльной головни овса (табл. III, а; V, а; VII, а).
Пыльная головня овса - одно из хорошо изученных головневых заболеваний, издавна известных земледельцам. Распространено оно на всех континентах, всюду, где возделывается овес.
Заболевание проявляется в период выметывания метелки При этом поражается обычно вся метелка. Все части цветка полностью разрушаются, замещаясь пылящими черными сорусами гриба. Сохраняются лишь осевые части метелки и колосков. При задержке развития возбудителя поражение сосредоточивается в нижней части метелки. В этом случае верхние цветки остаются непораженными, образуя здоровые зерновки. В редких случаях спороношения образуются на верхних листьях овса в виде темных полос.
Заболевание проявляется в двух формах: мягкой, наиболее часто встречающейся, с пылящим сорусом, и твердой - с компактным плотным сорусом, сходным с таковым Ustilago kolleri. При гибридологическом анализе признака строения соруса в пределах вида U. avenae показано, что признак "пыльный" сорус полностью доминирует над признаком "компактного" соруса (Holton, 1931).
Телиоспоры U. avenae 4.8 - 8, чаще 6.2 мкм, темные, шиповатые, иногда почти гладкие, шаровидные или короткоэллипсоидальные, сходные по морфологии и размерам со спорами U. nuda.
U. avenae по ряду признаков близок к U. kolleri (возбудителю покрытой головни овса). Оба вида различаются по строению спор (у U. kolleri они гладкие), а также по симптомам: у U. avenae преобладает пылящий тип соруса, у U. kolleri - компактный. Вместе с тем в природных условиях даже в пределах одной метелки можно обнаружить переходные формы между гладкими и шиповатыми спорами. В эксперименте и, вероятно, в природе U. avenae легко скрещивается с U. kolleri с образованием промежуточных форм как по образуемым симптомам, так и по морфологии спор. При этом во многих скрещиваниях признаки U. avenae доминируют над U. kolleri.
U. avenae весьма близок к возбудителю головни райграса - U. perennans, с которым легко скрещивается (Fischer, Holton, 1941). Согласно инокуляционным опытам, U. avenae не заражает райграс, a U. perennans не инфицирует овес (Rosch, 1926). Для обоих видов указывается общий хозяин - овсюг (Avena fatua L.). Ряд авторов рассматривает эти виды как синонимы (McAlpine, 1910; Fischer, 1953, и др.). Однако характер замещения сорусами элементов колосков овса и райграса у этих видов существенно различен, что наряду с неодинаковым кругом растений-хозяев позволяет считать их самостоятельными видами (Hille, 1958). Кроме того, у U. perennans споры несколько меньших размеров (5 - 7 мкм) и имеют менее четкие щетинки, замещающие лишь 1/4 или 1/3 часть поверхности споры; остальная часть споры остается практически гладкой.
U. аvеnае, так же как и U. kolleri, поражает многих представителей трибы Aveneae - виды родов Аvеnа, Helictotrichon, Trisetum. Еще Н. И. Вавилов (1919) отмечал, что U. avenae в сильной степени заражает, помимо культурных, также и многие дикорастущие виды овсов, морфологически весьма отличные от Avena sativa. В связи с этим ясно, что U. avenae мало реагирует на сортовые различия культурного овса. Для U. аvеnае и U. kolleri общим хозяином при искусственном заражении является Bromus unioloides (Thomas et al., 1981). Кроме того, для обоих патогенов выявлены и другие общие виды растений- хозяев из трибы Triticeae - Hordeum compressus и Agropyron tsukushiense var. transiense. * Эти растения инфицирует еще и возбудитель черной головни ячменя - U. nigra, с которым U. avenae сближает, помимо сходства в морфологии спор, также споридиальный способ прорастания и проростковый путь инфекции.
* (Как полагает Нильсен (Nielsen, 1978а), данная разновидность весьма близка, а может быть, идентична виду Roegneria pendulina (=Elymus pendulinus), произрастающему на советском Дальнем Востоке.)
По мнению Нильсена (Nielsen, 1978b), сходные по морфологическим и культуральным признакам пары U. avenae - U. nigra и U. hordei -U. kolleri родственны друг другу, а также виду U. bullata, возможному их предку. При скрещивании U. hordei и U. bullata (Fischer, 1951) во 2 - 4-м поколениях встречались споры, морфологически сходные с телиоспорами U. avenae - вида, который не использовался в скрещиваниях.
Для U. avenae характерна генеративно-проростковая инфекция, промежуточная между проростковой и эмбриональной. В этом случае летом инфицируются поверхностные ткани формирующейся зерновки. Мицелий зимует, распадаясь на толстостенные клетки разнообразной, чаще прямоугольной формы (геммы). Все части зародыша при поверхностной летней инфекции остаются непораженными (Gage, 1927). Весной следующего года перезимовавшие в зерновке гифы инфицируют в почве проростки овса. По другим наблюдениям, главную роль в инфекции играют все же непроросшие споры, зимующие на разных участках зерновки или метелки (например, на внутренней стороне чешуек, под пленками, на цветоносах, на остатках подсыхающего рыльца, пыльниках и т. д.), тогда как в тканях перикарпия зимующий мицелий практически отсутствует (Mills, 1967).
Из спор, приставших к поверхности зерновки, принимают участие в заражении только те споры, которые находятся в непосредственной близости от проростка.
При искусственном заражении легче всего инфицируются молодые проростки. Так, еще по сведениям Брефельда (Brefeld, 1895), при инокуляции спор в момент прорастания зерна заражается 17 - 20 % проростков, при длине проростка 1 см - 7 %, 2 См - 1 %. При длине проростков 8 см они совсем не заражаются, поскольку мицелий не достигает конуса нарастания побега.
В культуральных условиях у U. avenae отсутствует дикарио- тическая фаза и все клетки гриба оказываются одноядерными. Формирующиеся в заключительный период роста толстостенные пигментированные клетки являются вегетативными хламидоспорами, но не телиоспорами возбудителя (Флеров, 1923).
Телиоспоры U. avenae прорастают с образованием 4-клеточного промицелия, образующего споридии. При пониженных температурах 3 - 5°С споры не прорастают, как обычно, с образованием 3- или 4-клеточного промицелия и латеральных споридий, а формируют на концах промицелия ответвления гиф, анастомозирующие между собой, наподобие верхушечных споридий видов рода Urocystis (Goth, 1963). Не исключено, что при низких весенних температурах имеет место подобное прорастание телиоспор в почве при инфицировании ими зерновок. Имеются также свидетельства о прорастании телиоспор в естественных условиях с образованием инфекционных гиф без формирования споридий (Western, 1937).
Споридиальный тип роста U. avenae делает его удобным объектом как при гибридологическом анализе, так и при изучении возбудителя с использованием ауксотрофных мутантов. Генетика взаимоотношения этого вида с растением-хозяином укладывается в рамки гипотезы "ген-на-ген" (Halisky, 1965).
При скрещивании U. avenae и U. kolleri во многих случаях отмечено полное доминирование признака пыльного соруса, характерного для U. avenae, над компактным (непыльным) сорусом U. kolleri, а также доминирование признака шиповатости спор над гладкоспоровостью (Наnnа, Рорр, 1930; Holton, 1931). Тенденция к доминированию признака пыльного соруса и шиповатости спор свойственна практически всем исследованным видам головневых грибов. С использованием внутривидовых скрещиваний U. avenae, а также при его скрещивании с близким видом U. kolleri был проанализирован также характер наследования таких признаков, как окраска телиоспор, патогенность, симптоматология и т. д. В результате исследований показано независимое наследование признаков патогенности от признаков симптомов поражения. При этом различия по типам строения сорусов определяются одной парой аллельных генов, а различия по уровню патогенности - другой. Эти же данные говорят об отсутствии сцепления между этими генами, находящимися, по-видимому, в разных хромосомах (Левитин, Федорова, 1972).
Телиоспоры U. avenae прорастают без периода покоя. Минимальная температура для прорастания спор 0 - 5°С, оптимальная 22 - 25, максимальная 31 - 35 °С (Niemann, Schuhmann, 1962). Низкие почвенные температуры, удлиняя время нахождения в почве проростков, одновременно увеличивают возможность заражения всходов спорами патогена. Влажность почвы во время прорастания проростков, как и ее температура, во многом определяют степень инфекции. Обычно заражение происходит в интервале температур от 3 до 30°С и влажности - от 5 до 60 %. Наиболее благоприятная температура для инфекции 15 - 20 °С, а влажность - в пределах 35 - 40 %. При влажности почвы свыше 80 % прорастание спор в значительной степени заторможено, что связано с недостаточным доступом к ним кислорода. Повышенная влажность в зернохранилищах благоприятствует заражению мицелием поверхностных тканей зерновок овса.
Пыльная головня овса относится к числу весьма вредоносных заболеваний. Ее вредоносность главным образом проявляется в недоборе зерна. Кроме того, нередко снижаются всхожесть растений и качество зерна. На отдельных участках в Коми АССР потери достигали 70 % (Олимпиева, 1971), а в Кировской области - 94 % обследованной площади (Суслова, Веселкoва, 1980). В СССР к 1973 г. было идентифицировано 17 физиологических рас патогена (Тимошенко, 1973).
Против возбудителя пыльной головни овса широко используется протравливание семян. Эффективной мерой является их заблаговременное протравливание меркурбензолом (1.5 кг/т) и меркургексаном (1.5 - 2 кг/т). Весьма полезна также предпосевная обработка семян формалином (0.38 л/т), а также намачивание семян в малахитовой зелени в концентрации 0.2 % в течение 8 ч. Эффективны также обработки семян овса витаваксом и беномилом. При пониженной всхожести семян использование химических мер защиты недопустимо. К сожалению, в настоящее время проблема создания высокоустойчивых сортов к U. avenae остается нерешенной. Большинство районированных в СССР сортов овса сильно восприимчиво к U. avenae. Ценным источником для селекции овса на устойчивость к головневым и ржавчинным заболеваниям являются сорта овса из сортиментов CША и Канады, где имеются иммунные сорта к U. avenae. Устойчивые овсы из этих стран в основном представлены видом Avena byzantina и его межвидовыми гибридами с A. sativa (Тимошенко, Родионова, 1977).

Табл. I. Внешний вид заболеваний.а - здоровый колос пшеницы; б - колос пшеницы, пораженный Tilletia caries (DC.) Tul.; в - колос пшеницы, пораженный Ustilago tritici (Pers.) Jens.; г - колос пшеницы, пораженный U. tritici (Pers.) Jens, на заключительной стадии развития головни; д - колос пшеницы, пораженный Tilletia indica Mitra. Уменьш. в 4 - 5 раз
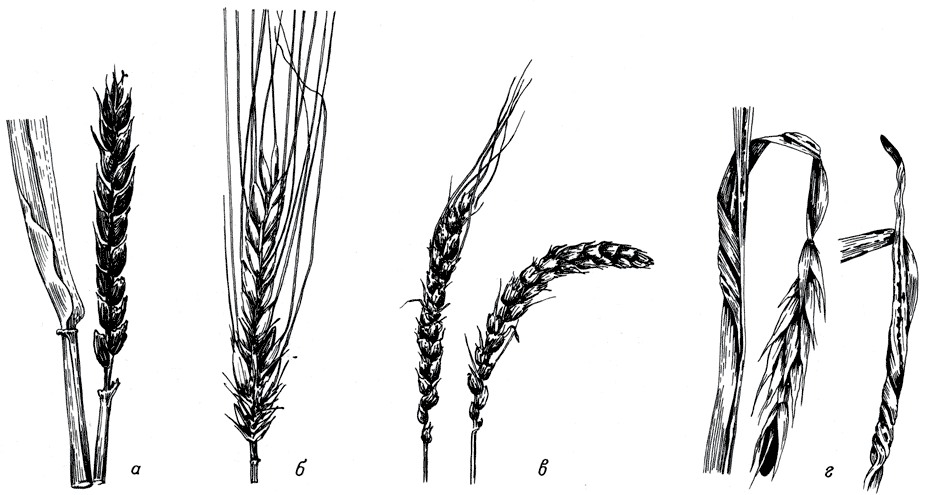
Табл. II. Внешний вид заболеваний.а - колос ячменя, пораженный Ustilago nuda (Jens.) Rostr.; б - колос ржи, пораженный U. vauilovii Jacz.; в - колос ячменя, пораженный U. hordei (Pers.) Lagerh.; г - стебель и листья ржи, пораженные Urocystis occulta (Wallr.) Rab. Уменьш. в 4 - 5 раз

Табл. III. Внешний вид заболеваний.а - метелка овса, пораженная Ustilago avenae (Pers.) Jens.; б - початок кукурузы, пораженный U. zeae (Beckm.) Ung.; в - початок кукурузы, пораженный Sphacelotheca reilianum Clint.; г - соцветие проса, пораженное S. destruens (Schlecht.) Stev. et John.; д - зерновки сорго, пораженные S. sorghi Ehrenb. (по:Zambettakis, 1971); е - зерновки сорго, пораженные Tolyposporium ehrenbergii (Kuehn) Pat. а, г-е-уменьш. в 3 раза; б, в - в 4 раза
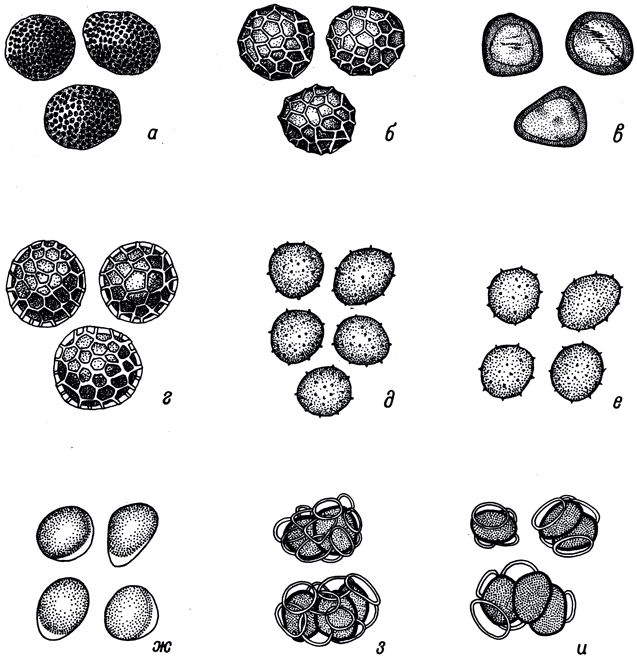
Табл. IV. Морфология телиоспор. а - Tilletia intermedia (Gassner) Savul.; б - Т. triticoides Savul.; в - Т. laevis Kuehn; г - Т. controversa Kuehn; д - Ustilago tritici (Pers.) Jens.; e - U. nuda (Jens.) Rostr.; ж - U. hordei (Pers.) Lagerh.; з - Urocystis tritici Koern.; и - Urocystis occulta (Wallr.) Rab. X1300
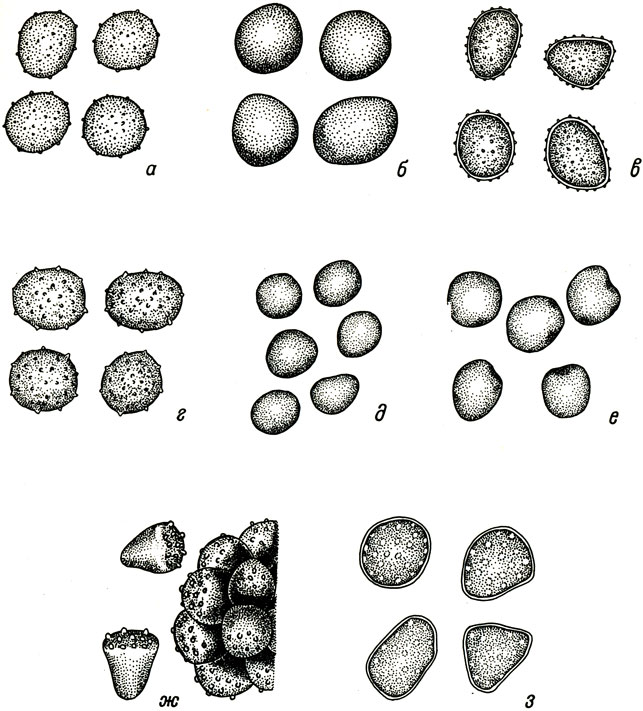
Табл. V. Морфология телиоспор. а - Ustilago avenae (Pers.) Jens.; б - U. kolleri Wille; в - U. bullata Berk.; г - U. zeae (Beckm.) Ung.; д - Sphacelotheca sorghi Ehrenb.; e - S. cruenta (Kuehn) A. A. Potter; ж - Tolyposporium ehrenbergii (Kiehn) Pat.; з - Ustilago crameri Koern. (no: Zambettakis, 1971). X 1300
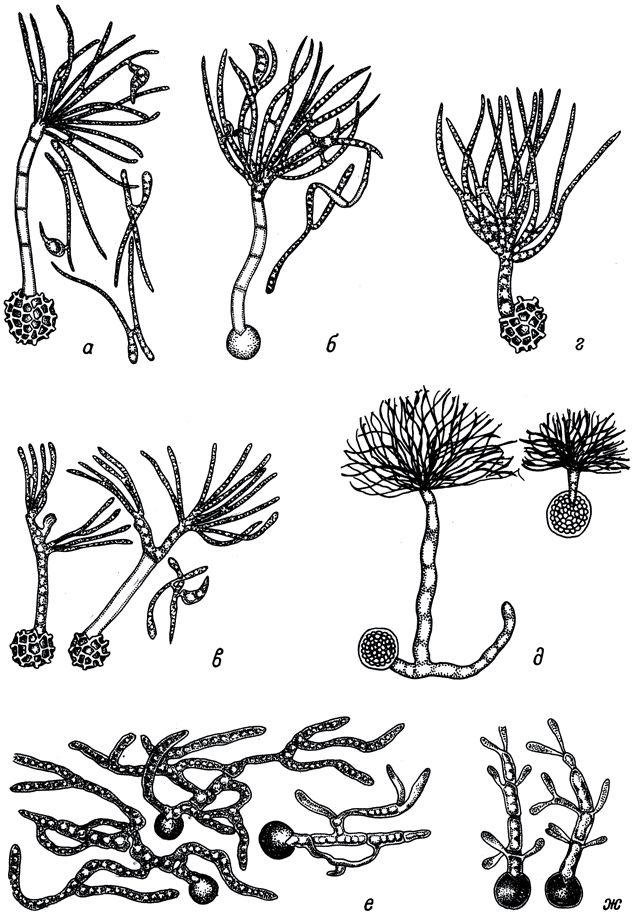
Табл. VI. Прорастаниетелиоспор. (По: Zambettakis, 1973). а - Tilletia caries (DC.) Till.; б - Т. laevis Kuehn; в - Т. controversa Kuehn; г - Т. secalis (DC.) Koern.; д - Т. indica Mitra; e - Ustilago tritici (Pers.) Jens.; ж - U. nuda (Jens.) Rostr. X1200
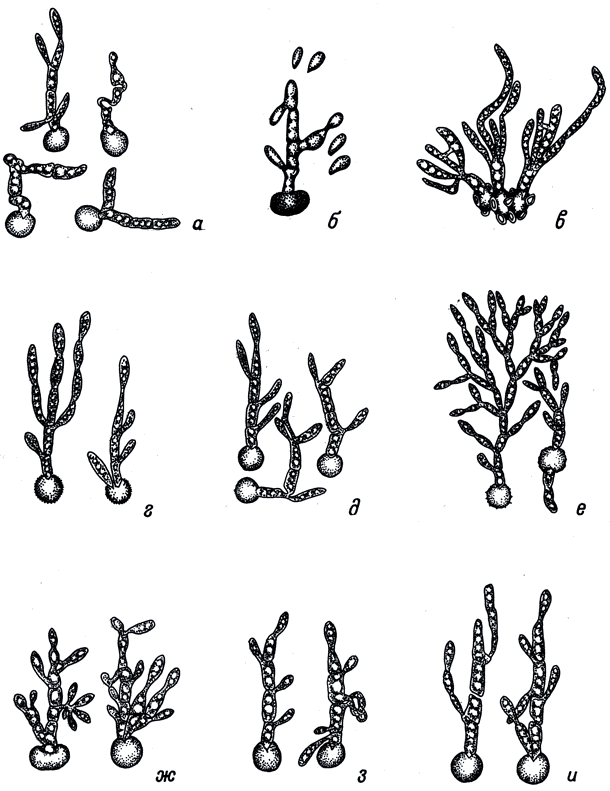
Табл. VII. Прорастаниетелиоспор. (По: Zambettakis, 1973). а - Ustilago hordei (Pers.) Lagerh.; б - U. nigra Тар k е; в - Urocystis tritici Koern.; г - Ustilago avenae (Pers.) Jens.; д - U. kolleri Wille; e - U. zeae (Beckm.) Ung.; ж - Sphacelotheca reiliana Clint.; з - S. cruenta (Kuehn) A. A. Potter; и - S. sorghi (Lk.) Clint. X1200
Ustilago kolleri Wille - возбудительтвердой, илипокрытой, головниовса (табл.V, б; VII, д).
Синоним: U. levis (Kell. et Swing.) Magn.
U. kolleri - возбудитель твердой головни овса - был впервые выделен в качестве разновидности U. avenae в 1890 г. в Северной Америке. Позднее, в 1893 г., он был признан в Норвегии в качестве самостоятельного вида.
Покрытая головня овса широко распространена по земному шару, встречаясь, как и U. avenae, практически всюду, где культивируется овес. Обычно U. kolleri регистрируется реже, чем U. avenae, и часто смешивается с ним, тем более что в пределах вида U. avenae имеются формы, по симптомам сходные с U. kolleri. В СССР возбудитель встречается в отдельных областях в европейской части СССР, Сибири и в северных районах Дальнего Востока. 3. В. Тимошенко (1973) отмечает, что ареал U. kolleri за последние годы существенно сократился и в некоторых областях, где возбудитель был хорошо известен, повторно выявить его не удается. По-видимому, немаловажную роль в этом сыграли профилактические мероприятия по обеззараживанию семян. Не обнаружено заболевание в Приморском крае. В целом U. kolleri встречается гораздо реже, чем U. avenae; при этом вредоносность твердой головни овса меньше, чем пыльной. К 1972 г. в СССР на основе анализа образцов инфицированного овса из различных регионов была выделена единственная физиологическая раса - U. Kolleri.
Гриб полностью разрушает зерновки обычно по всей метелке овса. Вследствие укорочения и недоразвития цветоносов метелки приобретают как бы компактный облик. Созревающая споровая масса замещает собой не только зерновки, но также и цветковые чешуи и часто основания колосковых чешуй. Сами колосковые чешуи в отличие от поражения при пыльной головне овса сохраняются целыми и прикрывают созревающие сорусы гриба. В некоторых случаях при слабом заражении верхняя часть метелок может остаться непораженной и на ней формируются нормальные зерновки. Споры U. kolleri не распыливаются наподобие спор U. avenae, поскольку склеены между собой слизистым веществом соруса, а распадаются на отдельные комочки при созревании соруса. Иногда поражение образуется на верхних листьях овса. Споры U. kolleri темно-коричневые, 4.6 - 8.1, чаще 6.3 мкм и всегда гладкие, чем они отличаются от шиповатых спор U. Avenae.
U. kolleri, помимо обычного овса, способен инфицировать также дикорастущие виды рода Avena. Кроме того, при искусственном заражении этот вид патогенен также для некоторых видов родов Agropyron и Hordeum (Nielsen, 1978b). Гриб поражает также Bromus unioloides (Thomas et al., 1981). U. kolleri близок к U. avenae и хорошо скрещивается с ним, при этом гибридное потомство имеет более широкий круг растений-хозяев, чем родительские формы. Ряд микологов объединяет виды U. kolleri и U. hordei, имеющие гладкие споры, в один вид на основе их морфологического сходства (Fischer, 1943; Ainsworth, Sampson, 1950, и др.) - Такому объединению противоречит разная специализация этих видов: U. kolleri не поражает ячмень, a U. hordei не инфицирует овес. В то же время при искусственном заражении выявлены общие растения-хозяева для этих видов - Agropyron tsukushiense var. transiense и Hordeum compressum, что в известной мере подтверждает их близость. Общим предком для U. kolleri, так же как и для U. hordei, U. avenae, по- видимому, явился U. bullata - вид с шиповато-бородавчатыми спорами, поражающий широкий спектр видов дикорастущих злаков (Nielsen, 1978b). При анализе качественного состава полипептидов U. kolleri и U. avenae было выявлено значительное сходство между этими видами, а также их родство с такими видами, как U. nigra и U. hordei. В этой связи был сделан вывод о значительной филогенетической близости всех указанных видов и даже о целесообразности их объединения в один таксон (Kim et al., 1984).
Способ инфекции U. kolleri во многом аналогичен тому, что известно для U. avenae. Заражение растений происходит во время прорастания семян, причем гифы проникают большей частью у основания молодого проростка. Подобно U. avenae, инфекция этим видом также может осуществляться геммами или покоящимся мицелием, находящимся под пленками зерновки. Споры U. kolleri способны энергично прорастать уже при 6 - 10°С, в связи с чем цветковые пленки, а иногда и перикарпий зерновок могут инфицироваться при хранении семян, обмолоте и уборке. В целом U. kolleri менее теплолюбив, чем U. Avenae.
При прорастании спор образуется 3 - 4-клеточный промицелий, дающий споридий, обильно почкующийся. В естественной обстановке инфекция может осуществляться после скрещивания споридий, а также путем прямого прорастания телиоспор с образованием дикариотического мицелия (Brandwein, 1944).
U. kolleri принадлежит к видам, недостаточно изученным в генетическом отношении. Известно, что он относится к стабильным, слабо мутирующим видам. Холтон (Holton, 1936) установил, что на протяжении многих половых и бесполовых поколений этого вида в течение 18 лет сохраняется мутант-альбинос. Скрещивание U. kolleri с U. avenae позволило в результате гибридологического анализа сделать ряд ценных заключений относительно доминирования признаков головневых грибов, контролирующих патогенность, тип соруса, а также строение и окраску телиоспор.
Меры борьбы с покрытой головней овса такие же, как и с пыльной головней овса. Они состоят в обеззараживании семян химическими препаратами и выведении устойчивых сортов. Структура соруса возбудителя, в частности склеивание телиоспор в плотные комочки (как и у U. hordei), затрудняет эффективное применение фунгицидов.
Возбудители головни риса
Tilletia barclayana (Bref.) Sacc. et Syd. - возбудитель черной головни колосков риса.
Синонимы: Т. horrida Таk., Neouossia horrida (Таk.) Padw. et A. Khan.
Черная головня КОЛОСКОВ риса была впервые обнаружена в Японии в окрестностях Токио в 1896 г., при этом возбудитель был описан как Tilletia horrida Таk. Позже заболевание выявлено во многих других регионах Азии, Америки, Африки, где временами оно вызывает существенный ущерб. Например, в штате Южная Каролина (США) посевы были поражены головней на 25 %. Впоследствии было обнаружено, что патоген способен также инфицировать виды рода Pennisetum, образуя в колосках телиоспоры, идентичные по морфологии с телиоспорами ранее описанного О. Брефельдом вида Tilletia (Neovossia) barclayana на Pennisetum orientale (Tullis, Johnson, 1952). На основе сходства в морфологии и инокуляционных опытов Т. horrida стал рассматриваться в качестве синонима Т. barclayana.
В СССР патоген может быть карантинным объектом. Он был обнаружен в единичных экземплярах в 1927 г. на рисовых плантациях вблизи оз. Ханка и в долине р. Раздольной в Приморском крае (Абрамов, 1939). Позднее в этих местах возбудитель не выявлялся (Проценко, Аблакатова, 1941; Азбукина, 1980). Однако, по сообщению 3. М. Азбукиной, в сентябре 1984 г. на зерновках риса в двух районах Приморского края - Анучинском и Кировском - возбудитель был выявлен снова.
Заболевание обнаруживается чаще всего на 1 - 2 завязях метелки. Гораздо реже поражается большее число колосков. Пораженные завязи прикрыты чешуями, так что часто оказывается трудно отличить здоровые колоски от больных. В целом характер поражения напоминает таковой при индийской головне пшеницы. Телиоспоры гриба 22 - 35 мкм в диаметре, почти черные, с темно-коричневым экзоспорием, несущим длинные, заостренные на вершине шипы 2.5 - 4.2 мкм длины и бесцветные продолговатые придатки (апикулюсы), характерные для рода Neovossia. Таксономическое значение таковых придатков часто оспаривается, в связи с чем возбудителя в последнее время относят к роду Tilletia (Duran, Fischer, 1961; Азбукина, 1981). В то же время индийские микологи предпочитают рассматривать его в составе рода Neouossia (Singh et al., 1979). При прорастании телиоспор образуются многочисленные (до 50) нитевидные споридии 38 - 53 мкм длины, не копулирующие между собой.
Т. barclayana весьма близок к виду Т. pulcherrima, выявленному в образцах зерновок риса на Дальнем Востоке. Последний вид обычно поражает представителей трибы Paniceae (Panicum, Digitaria, Echinochloa) и нередко рассматривается в качестве синонима Т. barclayana.
Различия в морфологии телиоспор этих видов можно определить лишь с помощью сканирующего микроскопа (Азбукина, 1981). Виды несколько отличаются и характером поражения завязей. Так, завязи, инфицированные Т. pulcherrima, имеют яйцевидную форму, 3 - 4 мм длины, иногда согнутые, выступающие из слегка расходящихся чешуй. Завязи, пораженные Т. barclayana, плотно прикрыты чешуями.
Споры гриба зимуют на почве и прорастают, плавая по поверхности воды. Споридии ветром переносятся на завязи растений в стадии цветения или восковой спелости, при этом возбудитель локально инфицирует отдельные завязи (Templeton, 1961). Заболевание обнаруживается вскоре после заражения. Следовательно, почвенной инфекции проростков и системного развития мицелия в тканях не происходит. Возбудитель обычно передается семенами. Заболевание не относится к числу вредоносных, однако контроль за ним необходим. Защитные мероприятия сводятся к обработке поверхности зерновок риса. При исследовании кариологии прорастающих телиоспор этого вида получены свидетельства его гомоталлизма, как и в случае Т. indica (Singh, Pavgi, 1972).
Entyloma oryzae Syd. et P. Syd. - возбудитель листовой головни риса.
Головня на листьях риса впервые была обнаружена в 1913 г. на Филиппинских островах. Позднее болезнь выявлена на плантациях риса в Японии, Китае, Афганистане, Индии, южных штатах США, Аргентине, Венесуэле, а также в других странах Азии и Америки. В СССР головня листьев риса обнаружена в Краснодарском крае, но значительного распространения болезнь не имела, так что рисосеящие районы страны практически от нее свободны. Однако в связи с тем, что листовая головня способна причинить значительный ущерб посевам риса, это заболевание находится под контролем карантинной службы. Болезнь обычно рассматривается как невредоносная.
Для болезни характерно появление на обеих сторонах листьев пятен свинцово-серого цвета, округлых, угловатых или коротко- линейных, 0.5 - 2X0.5 - 1.5 мм, часто сливающихся между собой. При разрыве эпидермиса высвобождается темно-бурая масса телиоспор, неправильной формы, гладких, 8 - 11X7 - 10 мкм в диаметре.
Телиоспоры при наличии влаги прорастают без периода покоя, образуя одноклеточный промицелий с продолговатыми споридиями. Последние копулируют между собой на поверхности листьев, и инфекционная гифа внедряется в ткани. Телиоспоры сохраняют жизнеспособность на остатках растений и семенах не более года.
Возбудитель головни Е. oryzae весьма сходен по морфологии (а может быть, и идентичен) с видом Е. lineatum, поражающим листья цицании (Zizania latifolia), изредка встречающимся на Дальнем Востоке.
Литература
Абрамов И. И. Болезни сельскохозяйственных растений Дальнего Востока. Хабаровск, 1939. 294 с.
Агарков В. А. Морфология проявления и ранняя диагностика головни проса. - Селекция и семеноводство, 1951, вып. 10, с. 21 - 28.
Азбукина 3. М. Возбудители головневых болезней зерновых. - В кн.: Возбудители болезней сельскохозяйственных растений Дальнего Востока. М., 1980, с. 111-124.
Азбукина 3. М. О таксономии Tilletia barclayana (Bref.) Sacc. et Syd. - В кн.: Систематика, экология и география споровых растений Дальнего Востока. Владивосток, 1981, с. 39 - 40.
Альмуратов Н. О значении комочков хламидоспор возбудителя каменной головни ячменя. - Вестн. с.-х. науки, Алма-Ата, 1966, т. 9, № 12, с. 98 - 101.
Артьемьева Н. Н. Поражаемость образцов проса различными эколого-географическими популяциями головни. - В кн.: Сб. тр. аспирантов и молодых науч. сотр. ВИР. Л., 1961, № 1, с. 135-139.
Бейлин И. Г. Эпифитотия Tilletia secalis на южном Алтае. - Вестн. защиты раст., 1940, № 1 - 2, с. 230-234.
Бондарцев А. С. Болезни культурных растений и меры борьбы с ними. М.; Л., 1931. 600 с.
Вавилов Н. И. Иммунитет растений к инфекционным заболеваниям. М., 1919. 240 с.
Вареница Е. Т., Мозговой А. Ф., Смирницкая П. П. Вредоносность твердой головни озимой пшеницы в Нечерноземной зоне. - Вестн. с.-х. науки, 1977, № 9, с. 30 - 32.
Гаркавый П. Ф., Кирдогло Е. К. Некоторые вопросы селекции ярового ячменя на устойчивость к пыльной головне. - Тр. по прикл. ботан., генет. и селекции, 1977, т. 58, № 3, с. 18-23.
Гешеле Э. Э. Головневые окрестностей Синельникова. - Матер, по микол. и фитопатол., 1927, вып. 6, с. 92 - 95.
Горленко М. В. Сельскохозяйственная фитопатология. М., 1968. 436 с.
Горленко М. В. Миграции фитопатогенных микроорганизмов. М., 1975. 108 с.
Грисенко Г. В., Дудка Е. Л. Особенности развития пыльной головни кукурузы в зависимости от гидротермических факторов. - Бюл. Всесоюз. науч.-иссл. ин-та кукурузы, 1971, т. 4 (21), с. 51 - 54.
Грисенко Г. В., Дудка Е. Л. Выживаемость хламидоспор Sorosporium reilianum (Kuehn) McAlp. в полевых условиях. - Микол. и фитопатол., 1979, т. 13, вып. 1, с. 37 - 42.
Грисенко Г. В., Сиденко И. Е. Агротехника в борьбе с болезнями кукурузы. - Защита раст., 1967, № 3, с. 17 - 19.
Гутнер Л. С. Головневые грибы. М.; Л., 1941. 383 с.
Додов Д. Н., Митов Н. Праховита главня - Ustilago tritici Pers. по ръжта. - Изв. Центр, науч.-иссл. ин-та защиты раст., 1961, т. 1, с. 111 - 117.
Дорофеев В. Ф., Кривченко В. И. Иммунологическая характеристика редких видов пшеницы. Л., 1975. 48 с.
Дошимов У. Специализация штаммов возбудителя пыльной головни сорго. - В кн.: Матер. VI конф. по споровым раст. Средней Азии и Казахстана, Душанбе, 1978. 162 с.
Дошимов У. Головневые грибы сорго. Ташкент, 1982. 67 с.
Жукова И. В. Об условиях заражения озимой ржи грибом Urocystis occulta (Wallr.) Rab. - Микробиология, 1951, т. 20, № 2, с. 152-154.
Жуковский П. М. Культурные растения и их сородичи. Изд. 3-е. Л., 1971. 751 с.
Запрометов Н. Г. Болезни культурных растений в Средней Азии. Ташкент, 1925. 165 с.
Запрометов Н. Г. Болезни джугары (сорго). - Тр. Ташк. с.-х. ин-та, 1964, т. 16, с. 283-298.
Захарова Т. И. Биологические особенности и таксономическое положение возбудителя карликовой головни озимой пшеницы: Автореф. дис.... канд. биол. наук. М., 1963. 17 с.
Ивахненко А. Н. Природа гипертрофических разрастаний соцветий кукурузы, зараженных пыльной головней. - Тр. Харьк. с.-х. ин-та, 1962, т. 38 (75), с. 86-99.
Игнатавичюте М. К. Головневые грибы Прибалтики. Вильнюс, 1975. 278 с.
Каратыгин И. В. Диморфизм прорастания спор Ustilago maydis (DC.) Cda. на листьях Zea mays. - Докл. АН СССР, 1968, т. 183, № 6, с. 1458-1460.
Каратыгин И. В. Генетическая характеристика цикла развития Ustilago maydis (DC.) Cda. - Микол. и фитопатол., 1969, т. 3, вып. 4, с. 368 - 376.
Каратыгин И. В. Географическое распространение Ustilago maydis (DC.) Cda. - Новости сист. низших раст., 1970, т. 6, с. 161 - 170.
Каратыгин И. В. Мицелий Ustilago maydis (DC.) Cda в тканях патологических новообразований при пузырчатой головне. - Докл. АН СССР, 1971, т. 201, № 6, с. 1500 - 1503.
Каратыгин И. В. Половая несовместимость у видов порядка Ustilaginales в связи с изменчивостью физиологических рас. - Журн. общ.биол., 1973, т. 34, вып. 6, с. 916 - 922.
Каратыгин И. В. Структура соруса и таксономическое положение Sorosporium realianum (Kuehn) McAlp. - Новости сист. низших раст., 1977, т. 14, с. 80 - 84.
Каратыгин И. В. Головневые грибы: Онтогенез и филогенез. Л., 1981. 202 с.
Каратыгин И. В. Место головневых грибов в системе грибных организмов. - В кн.: Эволюция и систематика грибов. Л., 1984, с. 110 - 118.
Кардаева Н. С. Источники сохранения возбудителя головни проса (Sphacelotheca panicimiliacei (Pers.)Bub.) и обоснование мер борьбы с ней в Волгоградской области: Автореф. дис.... канд. биол. наук. Л., 1974. 24 с.
Кечек Н. А. К вопросу о сохраняемости жизнеспособности хламидоспор твердой головни в почве. - Изв. АН АрмССР. Биол. и с.-х. науки, 1950, т. 3, № 1, с. 29-37.
Кривченко В. И. Физиологическая специализация и распространение рас Ustilago tritici (Pers.) Jens, в СССР. - Микол. и фитопатол., 1970, т. 4, № 2, с. 130 - 138.
Кривченко В. И. Изучение расового состава головневых грибов зерновых культур. - В кн.: Генетика и селекция болезнеустойчивых сортов культурных растений. М., 1974, с. 136 - 155.
Кривченко В. И. Проблема размещения источников устойчивости к инфекционным болезням зерновых культур для селекции на иммунитет. - Тр. по прикл. ботан., генет. и селекции, 1977, т. 58, вып. 3, с. 7 - 13.
Кривченко В. И. Использование генофонда в селекции сельскохозяйственных культур на устойчивость к вредным организмам. - В кн.: Проблемы защиты растений от вредителей, болезней и сорняков. М., 1979, с. 114 - 118.
Кривченко В. И., Мягкова Д. В. Взаимоотношения растения-хозяина и возбудителя пыльной головни. - Тр. V Всесоюз. совещ. по иммунитету растений. 2. Зерновые культуры, 1969, вып. 2, с. 55 - 59.
Кривченко В. И., Ямалеев А. М., Мигушова Э. Ф. Устойчивость к пыльной головне и геномный состав пшеницы. - Генетика, 1976, т. 12, № 4, с. 5 - 11.
Кузнецов Л. В. О жизнеспособности гриба Ustilago zeae (Beckm.) Ung. - возбудителя пузырчатой головни кукурузы. - Вестн. Моск. гос. ун-та. Сер. VI, Биол. и почв., 1963, вып. 2, с. 30 - 39.
Кузнецов Л. В., Скрипка О. В. О взаимоотношениях диплоидных аргинин-зависимых мутантов гриба Ustilago zeae (Beckm.) Ung. с растением-хозяином Zea mays. - Микол. и фитопатол., 1974, т. 8, вып. 3, с. 189 - 193.
Кульпинова М. П. Крупнопузырчатая головня сорго. - Тр. Азерб. с.-х. ин-та Сер.агробиол., 1957, т. IV, с. 26 - 34.
Курсанов Л. И. Микология. М., 1940. 480 с.
Лавров Н. Н. Определитель растительных паразитов культурных и дикорастущих полезных растений Сибири. Вып. 1. Томск, 1932. 148 с.
Лавров Н. Н. Флора грибов и слизевиков Сибири. Вып. 3. Микофлора хлебных злаков. - Тр. Томск, гос. ун-та, 1948, т. 104, с. 1 - 80.
Ландсберг Г. С. Начальные гистопатологические изменения кукурузы при галлогенезе, вызываемом Ustilago maydis (DC.) Cda. - Микол. и фитопатол., 1974, т. 8, вып. 1, с. 61 - 63.
Левитин М. М., Федорова И. В. Генетика фитопатогенных грибов. Л, 1972. 215 с.
Лобик А. И. О нахождении мокрой головни Tilletia foetens (Berk, et Curt.) Trel. на ржи Secale cereale. - Изв. Сев.-Кавк. ст. защиты раст., 1930, № 6 - 7, с. 165-166.
Мурашкинский К. Е. Tilletia controversa в предгорьях Алтая. - В кн.: Проблема пшенично-пырейных гибридов. М., 1937, с. 207 - 210.
Мурашкинский К. Е. Микофлора семян из археологических образцов. - Природа, 1940, № 4, с. 81-83.
Мягкова Д. В. Генофонд видов и групп пшениц, устойчивых к пыльной и твердой головне. - Тр. по прикл. ботан., генет. и селекции, 1977, т. 58, вып. 3, с. 72-76.
Мягкова Д. В. Эффективность генов устойчивости "Ut" к пыльной головне пшеницы. - Тр. конф.: Проблемы и пути повышения устойчивости растений, Л., 1981, ч. 3, с. 30-31.
Немлиенко Ф. Е. Пузырчатая головня кукурузы в степи УССР. - Тр. Укр. науч.-иссл. ин-та зерн. хоз-ва, 1947, т. 14, с. 3-59.
Новотельнова Н. С. Индийская головня пшеницы. - Тр. Всесоюз. науч.-иссл. ин-та защиты раст., 1970, т. 29, с. 134 - 142.
Олимпиева М. Ф. Головня зерновых культур. - В кн.: Прогноз распространения новейших вредителей и болезней сельскохозяйственных культур. М., 1971 - 1979.
Пальчевский Н. А. Болезни культурных злаков Южно-Уссурийского края. Спб., 1891. 80 с.
Пидопличко Н. А. Грибы-паразиты культурных растений: Определитель. Т. 1. Грибы совершенные. Киев, 1977. 296 с.
Понировский П. Н. Пыльная головня ячменя в Харьковской области. - Защита раст., 1962, № 2, с. 21.
Попов В. И., Силаев А. И. Эффективность протравливания семян сорго против двух видов головни. - Науч. тр. Ленингр. с.-х. ин-та, 1978, т. 351, с. 85 - 87.
Проценко А. Е., Проценко Е. П. О центрах формирования твердой головни Tilletia. - Бюл. Всесоюз. ин-та растениеводства, 1960, вып. 8, с. 19 - 21.
Проценко Е. П., Аблакатова А. А. Обследование риса на черную головню (Tilletia horrida Таk.) в Приморском крае. - В кн.: Справочник по вопросам карантина растений. 1941, № 2, с. 8-9.
Салунская Н. И. Мшливость популяцiй гриба Ustilago zeae (Beckm.) Ung. - збудника пухирчастоi сажки кукурудзи. - 3axicт рослин, 1969, т. 7, с. 41 - 51.
Силаев А. И., Чумаков А. Е. Головня сорго в Поволжье: Распространенность, вредоносность и болезнеустойчивость сортообразцов. - Микол. и фитопатол., 1979, т. 13, вып. 5, с. 414-418.
Спагенберг Г. Е., Гутнер Л. С. Изучение расового состава твердой головни пшеницы (Tilletia levis и Т. tritici) в полевых условиях. - В кн.: Итоги работ ВИЗР за 1935 г. Л., 1936, с. 489-491.
Степанов К. М., Чумаков А. Е. Прогноз болезней сельскохозяйственных растений. Л., 1967. 208 с.
Степановских А. С. Гистология паразитизма Ustilago nigra Tapke, возбудителя черной пыльной головни ячменя. - Микол. и фитопатол., 1978, т. 12, вып. 4, с. 324-328.
Степановских А. С. Рекомендации по оздоровлению ячменя от головни. Курган, 1982. 23 с.
Строчевой А. Л. Карликовая головня - опасная болезнь озимой пшеницы. - Защита раст. от вредителей и болезней, 1958, вып. 3, с. 30






