Микроорганизм | Наименование | Локализация | Районы | Стабильность | Чужеродность происхождения, G-C: % островов/ % хромосомы | Функция | Размер, Кб |
Уропатогенная E. coli 536 | Остров патогенности I (Pai I) | Sel Ca, 82' | 16 кб, прямые повторы, производные от sel C общий мотив с Pai II повторами | Нет | Прямые повторы; отсутствуют в E. coli нормальных фекалий и в лабораторных штаммах | альфа-гемолизин | 70
Уропатогенная E. coli 536 | Pai II | Leu Xa, 97' | 18 кб, прямые повторы, производные от leu X, общий мотив с Pai I повторами | Нет | То же | альфа-гемолизин, prf (фибрии, адгезивные к клеткам хозяина), транскрипционные активаторы хромосомных генов | 190
Уропатогенные E. coli J96 | (Pai I) | Вблизи Phe Va, 64' | — | — | То же | альфа-гемолизин, pap (фибрии, адгезивные к клеткам хозяина), последовательности IS-элементов, последовательности R-плазмид, последовательности фага Р4 | >170
Уропатогенные E. coli J96 | Pai II | Phe Ra,94' | 135 кб, неполный прямой повторяющийся | Нет | То же | альфа — гемолизин, pis (фимбрии, адгезивные для клеток хозяина), цитотоксический некротизирующий фактор I типа, последовательности IS-элементов, последовательности фага Р4, Omp R гомолог | 106
Энтеропатогенные E. coli | Локус стирания энтероцитов, LEE | Sel Ca, 82' | Повторов или IS-элементов не найдено | Даb | G+C:39/51, не показано тесной связи | Вызывает АЕ-поражения, секреторная система III-типа | 35
S. typhymurium | SPI 1 | Между fhe и mut S, 63' | Повторов или IS-элементов не найдено; | в некоторых серологически различных штаммах граничат с IS3 | Да | G+C:42/52, отсутствуют в E. coli | Инвазия в культуры эпителиальных клеток, секреторная система III-типа | 40
S. typhymurium | SPI 2 | между ydh E и pyk F, 31' | Даb | G+C:42/52, отсутствуют в E.coli, законсервирован среди Salmonella | III тип секреции | 40
S. typhymurium | Сальмонелла-индуцируемый жгутиковый ген А, sif A | potB/potC | 140 кб, прямые повторы | Да | G+C:41/52, прямые повторы; отсутствие в E. coli; законсервирован среди Salmonella | Для формирования структур, связанных с сальмонелла-ассоциированными вакуолями в пределах эпителиальных клеток | 1,6
Y. pestis | Способность к адсорбции экзогенных пигментов, Pgm | Pho E | 2,2 кб, прямой повтор (=IS 100) | Нет | G+C: регион накопления гемина 47/46–50; | Рецептор иерсинеобактина / регион железорегулируемого белка 56–60/46–50; | Прямые повторы | Связывание гемина и конго красного. | Пестицинчувствительность. | Связывание железа. | Рост при 37 °C в обедненной среде | 102
V. cholerae 0137 | Otn A otn B | rfb | Фланкирован двумя различными IS — элементами | — | IS-элемент; не содержится в Vibrio cholerae 01 El Tor | Синтез капсулы и О антигена | 35
L. monocytogenes | — | Между prs и ldh | IS-элементов не обнаружено | Даb | В непатогенных видах отсутствует | Избавляет листерию от вакуоли; внутри и внеклеточное распространение | 9,6
Примечание: а — ген tRNA; b — сравнительно
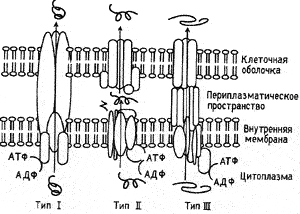
Рис. 13. Схема типов систем секреции бактерий. Для осуществления секреции все системы используют энергию АТФ-гидролиза. I и III типы секретируют белки через внутреннюю мембрану и клеточную оболочку бактерии за одну стадию; секретируемые белки не делают промежуточной остановки в периплазматическом пространстве, как это наблюдается при II типе секреции. Системы I и III типа сходны еще тем, что они не удаляют какой-то части секретируемого белка. В противоположность этому, N-концы белков, секретируемых по второму пути, утрачиваются ими при прохождении периплазматического пространства. Первый тип систем секреции представлен значительно меньшим количеством компонентов, чем третий (на рисунке это показано различающимися по форме и размеру белками). Третий тип секреции зависит от контакта с поверхностью эукариотической клетки [Galbn J., 1996]
У грамотрицательных бактерий такие белки проходят через две мембраны: внутреннюю, которая окружает цитоплазму, и внешнюю оболочку, которая является барьером между клеткой и окружающей средой. Между ними располагается периплазматическое пространство, через которое проходит общий путь секреции белков перед тем как была идентифицирована секреторная система рода Yersinia, уже были известны две другие системы (I и II типов) специализирующиеся на транспорте молекул к клеточной поверхности [Salmond G., Reeves PJ., 1993]. Протеины, секретируемые по механизму I типа, переходят непосредственно из цитоплазмы к клеточной поверхности, используя общий секреторный путь. Механизм секреции II типа использует этот путь до достижения белками периплазматического пространства. Затем белок переходит через наружную мембрану посредством других канальных белков. Обе системы (I и II типов) вовлечены в различные функции, включая ипатогенез. Например, альфа-гемолизин E. coli использует систему I типа Пилеформирующие энтеропатогеные E. coli (EPEC) и энтеропатогенные E. coli (BTBC), используют II тип секретирующей системы [Mecsas J., Strauss E.J., 1996].
3а последние годы у многих грамположительных бактерий, вызывающих болезни животных и растений, были обнаружены высококоннсервативные системы III типа [Gijsegem F., 1995]. Они ответственны за транспортировку эффекторных молекул непосредственно из цитоплазмы к клеточной поверхности, где эти молекулы взаимодействуют с клетками макроорганизма и изменяют функционирование их белков таким образом, чтобы способствовать выживанию и репликации бактерии [Michiels Т. et al., 1990]. Секреция эффекторных белков системой III типа происходит непосредственно после контакта возбудителя инфекционной болезни с клеткой хозяина (в отличие от систем I и II типов). Поэтому ее можно назвать контактзависимой секрецией [Galbn J., 1996]. Из возбудителей, использующих III тип секреторной системы, лучше всего изучена Yersinia pestis, вызывающая чуму, а также некоторые энтеропатогены (семейств Yersinia, Salmonella, Shigella и ЕРЕС), вызывающие как диарею, так и системные инфекции. Из-за того, что секретируемые бактериями эффекторные молекулы могут поражать различные цели, эти системные инфекции могут проявляться различными синдромами [Salyers A., Whitt D., 1994]. Представители семейства Yersinia используют для этих целей эффекторные молекулы, разрушающие ключевые функции иммунных клеток и делают их «безвредными» для себя [Cornelis G., 1992]. В опытах на культурах клеток было установлено, что Yersinia секретируется примерно 10 типов эффекторных молекул и, по крайней мере, 3 из них инъецируются в клетки-мишени [Rosqvist R., 1994]. Два типа — Yop E и Yop H, модифицируют белки макрофагов и нарушают способность макрофагиальных клеток нейтрализовать действие этих эффекторных молекул, что позволяет Yersinia «процветать» в ретикулоэндотелии.
В отличие от эффекторных молекул семейства Yersinia, блокирующих нормальные клеточные функции, эффекторные молекулы Shigella и одной из сальмонелл с III типом секреции, побуждают клетки к осуществлению ими функций, не являющихся для них обычными [Mecsas J., Strauss E.J., 1996].
«Острова патогенности» могут включать несколько различающихся фрагментов (табл. 2). Например, нестабильный участок ДНК, протяженностью 102 кб, отвечающий за вирулентность возбудителя чумы, включает несколько самостоятельных регионов. Один содержит ген накопления гемина — его G+C состав аналогичен G+C составу хромосомы чумного микроба. У другого региона, содержащего гены, кодирующие рецептор иерсиниобактина и железорегулирующего белка, процентное содержание G+C значительно выше характерного для хромосомы Y. pestis в целом. Весь 102 кб участок может быть делетирован у возбудителя чумы. Но делеция может захватить и только один из генов участка [Mecsas J., Strauss E.J., 1996]. В геноме другого представителя семейства — Y. enterocolitica, представлен только регион, кодирующий рецептор иерсиниобактина и железо-регулирующего белка [Fetherston J., 1992]. У других бактерий, например, Helicobacter pylori, строение «островов патогенности» еще сложнее [Mecsas J., Strauss E.J., 1996].
Нестабильность, как и наоборот, стабильность «островов», видимо не случайность в проявлении бактерией патогенности. Эти свойства способны создать бактерии дополнительные адаптивные преимущества. Высокая вирулентность может оказаться невыгодной бактерии на определенной стадии инфекционного процесса. Нестабильность же «островов патогенности» будет способствовать координированному снижению вирулентности всей популяцией возбудителя инфекции. Делеция «островов патогенности» может усилить экспрессию других, рядом расположенных, генов [Ritter A., 1995].
С другой стороны, отдельные факторы вирулентности являются адаптивными для бактерий. По этой причине они должны кодироваться на стабильных «островах патогенности». «Чужеродность» «островов» придает даже большую стабильность факторам вирулентности. Чужеродная ДНК, интегрировавшаяся с хромосомой, не вовлекается в рекомбинацию с ДНК близкородственных микроорганизмов, поэтому она может длительно поддерживаться в бактериальных популяциях [Falkow S., 1996]. Видимо применительно к каждому патогенному микроорганизму «стабильность» и «нестабильность» «островов» закрепляется естественным отбором.
В настоящее время нет полной ясности, каким образом «острова патогенности» приобретаются бактериями, но есть косвенные признаки того, что этот процесс идет непрерывно. Новый эпидемический штамм Vibrio cholerae О139 появился в результате приобретения «острова патогенности». Хотя известно, что он возник из штамма того же серотипа (О1), который является причиной происходящей в настоящее время пандемии холеры (О1 El Tor), V. cholerae О139 содержит дополнительный участок ДНК, который замещает часть О-антигенного кластера О1-штамма. Вставленная ДНК содержит открытую рамку считывания, гомологичную протеину, вовлеченному в синтез капсулы и О-антигена — двух факторов, по которым можно провести различия между О139 и О1 Е1 Тог. Тем самым повышается способность возбудителя холеры к инвазии [Mecsas J. Strauss E.J., 1996].
* * *
Бактерии являются мощными раздражителями иммунной системы, поэтому они могут использовать только первую стратегию паразитизма и в иммунокомпетентном хозяине лишь кратковременно имитировать вторую стратегию (L-формы). Бактерии обладают избыточными и альтернативными механизмами адгезии и колонизации, что позволяет им проявлять большую гибкость в выборе новых хозяев. Адгезия бактерий не является простым механическим взаимодействием их лиганд-структур с рецепторами на поверхности клеток-мишеней хозяина, имеющими другое предназначение. Взаимодействие патогена с клеткой хозяина приводит к активации сигнальных систем клеток-мишеней непосредственно каким-то бактериальным компонентом, либо через стимуляцию активационных факторов хозяина. Для противодействия иммунной системе бактерии используют как механизмы уклонения (экранирование клеточной стенки бактерий, антигенная мимикрия, образование L-форм бактерий и др.), так и разрушения (продуцирование секретируемых факторов, инактивирующих защиту хозяина; продуцирование «суперантигенов» и др.). Гены, кодирующие факторы патогенности бактерий и системы их секреции, вопреки распространенному еще недавно мнению об их преимущественно плазмидной локализации, в большинстве своем кластированны на участках хромосомной ДНК, называемых «островами патогенности». Эти участки активно участвуют в генетическом обмене между бактериями по пока еще неизвестному механизму. Патогенность «новых» для человека бактерий может объясняться тем, что они уже были преадаптированы к такой встрече в занимаемых ими ранее экологических нишах.






