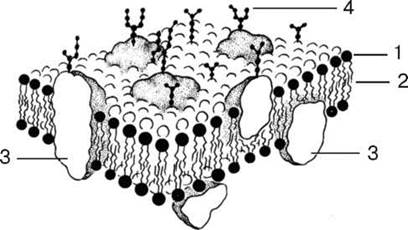
1 - липиды; 2 - гидрофобная зона би-слоя липидных молекул; 3 - интегральные белки мембраны; 4 - полисахариды гликокаликса
Общей чертой всех мембран клетки является то, что они представляют собой тонкие (6-10 нм) пласты липопротеидной природы (липиды в комплексе с белками).
Основными химическими компонентами клеточных мембран являются липиды (40 %) и белки (60 %); кроме того, во многих мембранах обнаружены углеводы (5-10 %).
К липидам относится большая группа органических веществ, обладающих плохой растворимостью в воде (гидрофобность) и хорошей растворимостью в органических растворителях и жирах (липофиль-ность). Состав липидов в разных мембранах неодинаков. Например, плазматическая мембрана в отличие от мембран эндоплазматической сети и митохондрий обогащена холестерином. Характерными представителями липидов, встречающихся в клеточных мембранах, являются фосфо-липиды (глицерофосфатиды), сфингомиелины и из стероидных липидов - холестерин.
Особенностью липидов является разделение их молекул на две функционально различные части: гидрофобные неполярные, не несущие зарядов («хвосты»), состоящие из жирных кислот, и гидрофильные, заряженные полярные «головки». Это определяет способность липидов самопроизвольно образовывать двухслойные (билипидные) мембранные структуры толщиной 5-7 нм.
Мембраны различаются и набором белковых молекул. Многие мембранные белки состоят из двух частей - участков, богатых полярными (несущими заряд) аминокислотами, и участков, обогащенных неполярными аминокислотами: глицином, аланином, валином, лейцином. Такие белки в липидных слоях мембран располагаются так, что их неполярные участки погружены в «жирную» часть мембраны, где находятся гидрофобные участки липидов. Полярная (гидрофильная) же часть этих белков взаимодействует с головками липидов и обращена в сторону водной фазы. Эти белки пронизывают мембрану, их называют интегральными белками мембран. Кроме интегральных белков, существуют белки, частично встроенные в мембрану, - полуинтегральные и примембранные, не встроенные в билипидный слой. По биологической роли белки мембран можно разделить на белки-ферменты, белки-переносчики, рецепторные и структурные белки.
Углеводы мембран входят в их состав не в свободном состоянии, они связаны с молекулами липидов или белков. Такие вещества называются соответственно гликолипидами и гликопротеидами. Как бы ни было велико различие между мембранами по количеству и составу их липидов, белков и углеводов, мембраны обладают рядом общих свойств, определяемых их основной структурой. Все мембраны являются барьерными структурами, резко ограничивающими свободную диффузию веществ между цитоплазмой и средой, с одной стороны, и между матриксом и содержимым мембранных органелл - с другой.
Плазматическая мембрана, или плазмолемма (membrana cellularis), среди различных клеточных мембран занимает особое место. Это поверхностная периферическая структура, не только ограничивающая клетку снаружи, но и обеспечивающая ее непосредственную связь с внеклеточной средой, а следовательно, и со всеми веществами и стимулами, воздействующими на клетку.
Химический состав плазматической мембраны. Основу плазмолеммы составляет липопротеиновый комплекс. Она имеет толщину около 10 нм и, таким образом, является самой толстой из клеточных мембран.
Снаружи от плазмолеммы располагается надмембранный слой - гликокаликс (glycocalyx). Толщина этого слоя около 3-4 нм, он обнаружен практически у всех животных клеток, но степень его выраженности различна. Гликокаликс представляет собой ассоциированный с плазмолеммой гли-копротеиновый комплекс, в состав которого входят различные углеводы. Углеводы образуют длинные, ветвящиеся цепочки полисахаридов, связанные с белками и липидами, входящими в состав плазмолеммы. При использовании специальных методов выявления полисахаридов (краситель рутениевый красный) показано, что они образуют структуру, подобную «чехлу» поверх плазматической мембраны.
В гликокаликсе могут располагаться белки, не связанные с билипидным слоем. Как правило, это белки-ферменты, участвующие во внеклеточном расщеплении различных веществ, таких как углеводы, белки, жиры и др.
Функции плазматической мембраны - это разграничение цитоплазмы с внешней средой, рецепции и транспорта различных веществ как внутрь клетки, так и из нее.
С плазмолеммой связана локализация специфических рецепторов, отвечающих за такие важные процессы, как взаимное распознавание клеток и развитие иммунитета. Так, в составе плазмолеммы всех клеток находится молекула гистосовместимости I класса (гликопротеин), которая состоит из:
a) интегрального трансмембранного белка, часть которого находится в цитоплазме, другая - пронизывает плазмолемму, и последняя, наиболее длинная часть молекулы, находится в гликокаликсе;
b) периферического мембранного белка с малой молекулярной массой;
c) короткой молекулы белка, которая нековалентно связывается с петлями внеклеточной части интегрального трансмембранного белка. Именно последняя часть молекулы (пептид из 9 аминокислот) является фрагментом нормального белка клетки данной особи. Он и распознается как «свой» клетками иммунной системы человека. В случае мутации на месте белка гистосовместимости появляется белок с иной структурой молекулы (например, кодируемый вирусом), и в ответ на это возникает иммунная реакция со стороны организма, направленная на уничтожение данной клетки. Этот механизм сохраняет генетическую индивидуальность клеток, следовательно, и организма.
В плазмолемме светочувствительных клеток животных расположена специальная система фоторецепторных белков (родопсин), с помощью которых световой сигнал превращается в химический, что, в свою очередь, приводит к генерации электрического импульса.
Эндоплазматическая сеть
Эндоплазматическая сеть (эндо-плазматический ретикулум) была открыта К. Р. Портером в 1945 г. Этот компонент вакуолярной системы клетки представляет собой совокупность вакуолей, плоских мембранных мешков или трубчатых образований, создающих трехмерную мембранную сеть. В состав сети входят гранулярные и агранулярные участки, которые могут чередоваться.
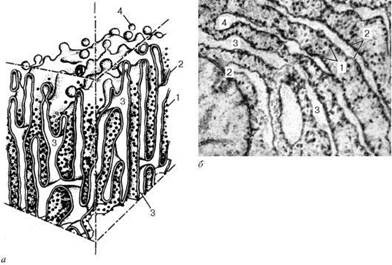
Строение гранулярной эндоплазматической сети: а - схема; б - электронная микрофотография участка среза эпителиальной клетки печени. 1 - рибосомы; 2 - пластинки; 3 - внутренние полости цистерн; 4 - отщепляющиеся мембранные пузырьки, лишенные рибосом.
Гранулярная эндоплазматическая сеть (reticulum endoplasmicum granulosum) на ультратонких срезах представлена замкнутыми мембранами, которые на сечениях образуют уплощенные мешки, цистерны, трубочки.
Диаметр цистерн значительно варьирует и в зависимости от функциональной активности клетки колеблется от 20 нм до несколько микрометров. Отличительной чертой мембран гранулярной эндоплазматической сети является то, что они со стороны гиалоплазмы покрыты многочисленными рибосомами.
Гранулярная эндоплазматическая сеть имеет разное строение. Для малоспециализированных клеток или для клеток с низкой метаболической активностью характерно наличие редких и разрозненных цистерн. Если возникают локальные скопления гранулярной эндоплазматической сети, то это свидетельствует об активном синтезе секреторных белков. Так, в клетках печени и некоторых нервных клетках гранулярная эндоплазматическая сеть собрана в отдельные зоны. В клетках поджелудочной железы гранулярная эндоплазмати-ческая сеть в виде плотно упакованных друг около друга мембранных цистерн занимает базальную и околоядерную зоны клетки. Рибосомы, связанные с мембранами эндоплазматической сети, участвуют в синтезе белков, выводимых из данной клетки («экспортируемые» белки). Кроме того, гранулярная эндоплазматическая сеть принимает участие в синтезе белков - ферментов, необходимых для организации внутриклеточного метаболизма, а также используемых для внутриклеточного пищеварения.
Белки, накапливающиеся в полостях эндоплазматической сети, могут, минуя гиалоплазму, транспортироваться в вакуоли комплекса Гольджи, где они модифицируются и входят в состав либо лизосом, либо секреторных гранул, содержимое которых остается изолированным от гиалоплазмы мембраной. Внутри канальцев или вакуолей гранулярной эндоплазматической сети происходит модификация белков, например, связывание их с сахарами (первичное глюкозилирование).
В гранулярной эндоплазматической сети на ее рибосомах происходит синтез мембранных интегральных белков, которые встраиваются в толщу мембраны. Здесь же со стороны гиалоплазмы идет синтез липидов и их встраивание в мембрану. В результате этих двух процессов наращиваются сами мембраны эндоплазматической сети и другие компоненты вакуоляр-ной системы клетки.
Агранулярная (гладкая) эндоплазматическая сеть (reticulum endoplasmaticum nongranulosum) также представлена мембранами, образующими мелкие вакуоли, трубки, канальцы, которые могут ветвиться, сливаться друг с другом. В отличие от гранулярной эндоплазматической сети на мембранах гладкой эндоплазматической сети нет рибосом. Диаметр вакуолей и канальцев гладкой эндоплазматической сети обычно около 50-100 нм. Гладкая эндоплаз-матическая сеть возникает и развивается на основе гранулярной эндоплаз-матической сети. В отдельных участках гранулярной эндоплазматической сети образуются новые липопротеидные мембранные участки, лишенные рибосом. Эти участки могут разрастаться, отщепляться от гранулярных мембран и функционировать как самостоятельная вакуолярная система.
Деятельность гладкой эндоплазматической сети связана с метаболизмом липидов и некоторых внутриклеточных полисахаридов. Гладкая эндоплазматическая сеть участвует в заключительных этапах синтеза липидов. Она сильно развита в клетках, секретирующих стероиды, например, в эндокринных клетках коркового вещества надпочечников, в эпителиальных клетках извитых семенных канальцев.
Тесная топографическая связь гладкой эндоплазматической сети с отложениями гликогена (запасной внутриклеточный полисахарид животных) в гиалоплазме различных клеток (клетки печени, мышечные волокна) указывает на ее возможное участие в метаболизме углеводов.
В поперечнополосатых мышечных волокнах гладкая эндоплазматиче-ская сеть способна депонировать ионы кальция, необходимые для функции мышечной ткани.
Очень важна роль гладкой эндоплазматической сети в дезактивации различных вредных для организма веществ за счет их окисления с помощью ряда специальных ферментов. Особенно четко она проявляется в клетках печени. Так, при некоторых отравлениях в клетках печени появляются ацидофильные зоны (не содержащие РНК), сплошь заполненные гладким эндо-плазматическим ретикулумом.
Комплекс Гольджи
Комплекс Гольджи (пластинчатый комплекс) был открыт в 1898 г. К. Гольджи. Автор, используя свойства связывания тяжелых металлов (осмия или серебра) с клеточными структурами, выявил в нервных клетках сетчатые образования, которые он назвал внутренним сетчатым аппаратом (apparatus reticularis internus). В дальнейшем его стали называть аппаратом, или комплексом Гольджи (complexus Golgiensis). Подобные структуры затем были описаны во всех клетках эукариот.
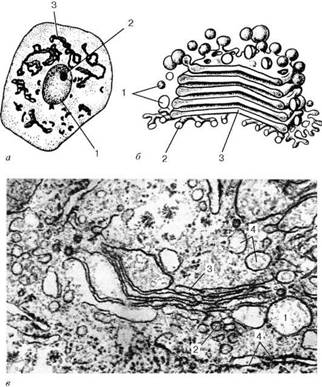
Комплекс Гольджи: а - нервная клетка спинного мозга, импрегнация серебром по методу Гольджи: 1 - ядро; 2 - ядрышко; 3 - комплекс Гольджи; б - схема ультрамикроскопического строения (трехмерная реконструкция); в - комплекс Гольджи на ультратонком срезе (печеночная клетка): 1 - пузырьки; 2 - трубочки; 3 - уплощенные мешочки (цистерны); 4 - фрагменты гранулярной эндоплазматической сети
Отдельная зона скопления этих мембран называется диктиосомой (стопкой Гольджи). Таких зон в клетке может быть несколько. В диктиосоме плотно друг к другу (на расстоянии 20-25 нм) расположены 5-10 плоских цистерн, между которыми находятся тонкие прослойки гиалоплазмы. Каждая цистерна имеет переменную толщину: в центре ее мембраны могут быть сближены (до 25 нм), а на периферии иметь расширения - ампулы, ширина которых непостоянна. Кроме плотно расположенных плоских цистерн, в зоне комплекса Гольджи наблюдается множество мелких пузырьков (везикул), которые встречаются главным образом в его периферических участках. Иногда они отшнуровываются от ампулярных расширений на краях плоских цистерн. В зоне диктиосомы различают проксимальную (cis) и дистальную (trans) поверхности. В секретирующих клетках обычно комплекс Гольджи поляризован: его проксимальная поверхность обращена к ядру, в то время как дистальная - к поверхности клетки.
В клетках отдельные диктиосомы могут быть связаны друг с другом системой везикул и цистерн, примыкающих к дистальной поверхности, так что образуется рыхлая трехмерная сеть, выявляемая в световом и электронном микроскопах («транс-сеть» комплекса Гольджи).
Комплекс Гольджи участвует в сегрегации и накоплении продуктов, синтезированных в эндоплазматической сети, в их химической перестройке, созревании; в его цистернах происходят синтез полисахаридов, их комплексирование с белками, что приводит к образованию пептидогликанов. С помощью комплекса Гольджи осуществляется процесс выведения готовых секретов за пределы секреторной клетки. Кроме того, комплекс Гольджи обеспечивает формирование лизосом. Внутри полостей комплекса Гольджи с помощью различных ферментов (транс-глюкозидаз) по разному модифицируются лизосомные белки и белки секретов: происходят последовательная замена и наращивание олигосахаридных цепочек.
В цистернах дистальной (trans) поверхности происходит сортировка белков: на внутренних поверхностях мембран цистерн располагаются рецепторы, узнающие или секреторные белки, или белки, входящие в состав лизосом (гидролазы). В результате от цистерн дистальной поверхности диктиосом отщепляются два типа мелких вакуолей: а) содержащие гидролазы - лизосомы (первичные); б) секреторные белки.
Секреторная функция комплекса Гольджи заключается в том, что синтезированный на рибосомах белок, накапливающийся внутри цистерн эндо-плазматической сети, транспортируется далее в вакуоли комплекса Гольджи.
Лизосомы
Лизосомы (lysosomae) - это разнообразный класс вакуолей размером 0,2- 0,4 мкм, ограниченных одиночной мембраной. Характерным признаком лизосом является наличие в них гидролитических ферментов - гидролаз (протеиназы, нуклеазы, фосфатазы, липазы и др.), расщепляющих различные биополимеры при кислом значении рН. Лизосомы были открыты в 1949 г. де Дювом.
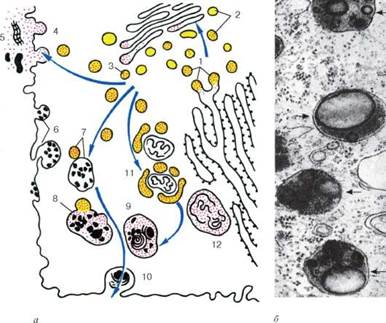
Строение лизосом: а - схема участия структур клетки в образовании лизосом и во внутриклеточном пищеварении: 1 - образование из гранулярной эндоплазматической сети мелких пузырьков, содержащих гидролитические ферменты; 2 - перенос ферментов в комплекс Гольджи; 3 - образование первичных лизосом; 4 - выделение и использование (5) гидролаз при внеклеточном расщеплении; 6 - эндоцитозные пузырьки; 7 - слияние первичных лизосом и эндоцитозных пузырьков; 8 - образование вторичных лизосом; 9 - телолизосомы; 10 - экскреция остаточных телец; 11 - слияние первичных лизосом с разрушающимися структурами клетки; 12 - аутофаголизосома; б - электронная микрофотография среза гетерофаголизо-сом (обозначены стрелками)
Кроме собственно лизосом (первичных) различают аутофаголизосомы, или гетеролизосомы (вторичные лизосомы), и телолизосомы (остаточные тельца).
Разнообразие морфологии лизосом объясняется тем, что эти частицы участвуют в процессах внутриклеточного переваривания, образуя сложные пищеварительные вакуоли как экзогенного (внеклеточного), так и эндогенного (внутриклеточного) происхождения.
Лизосомы (первичные) представляют собой мелкие мембранные пузырьки размером около 0,2-0,5 мкм, заполненные бесструктурным веществом, содержащим гидролазы, в том числе активную кислую фосфатазу, которая является маркерным ферментом для лизосом. Эти мелкие пузырьки практически очень трудно отличить от мелких везикул на периферии зоны комплекса Гольджи, которые также содержат кислую фосфатазу. Местом ее синтеза является гранулярная эндоплазматическая сеть. Затем этот фермент появляется в цистернах проксимальной поверхности диктиосомы, а затем в мелких везикулах по периферии диктиосомы и, наконец, в лизосомах. Таким образом, весь путь образования лизосом очень сходен с образованием секреторных (зимогенных) гранул в клетках поджелудочной железы, за исключением последнего этапа.
Гетерофаголизосомы (вторичные лизосомы), или внутриклеточные пищеварительные вакуоли, формируются при слиянии лизосом с фагоцитарными или пиноцитозными вакуолями. Если происходит слияние лизосомы с измененными органеллами самой клетки, то такая структура называется аутофаголизосома. При этом ферменты лизосомы получают доступ к субстратам, которые они и начинают расщеплять. Вещества, попавшие в состав гетероили аутофаголизосом (вторичных лизосом), расщепляются гидролазами до мономеров, которые транспортируются через мембрану лизосомы в гиалоплазму, где они реутилизируются, т. е. включаются в различные обменные процессы.
Однако расщепление, переваривание макромолекул лизосомой может идти в ряде клеток не до конца. В этом случае в вакуолях лизосом накапливаются непереваренные продукты. Такая органелла носит название тело-лизосома, или остаточное тельце (corpusculum residuale). Остаточные тельца содержат меньше гидролитических ферментов, в них происходит уплотнение содержимого, его перестройка. Часто в остаточных тельцах наблюдается вторичная структуризация неперевариваемых липидов, которые образуют слоистые структуры. Там же откладываются пигментные вещества. Например, у человека при старении организма в клетках мозга, печени и в мышечных волокнах в телолизосомах происходит отложение «пигмента старения» - липофусцина.
При участии лизосом (аутофаголизосом) может происходить модификация продуктов, которые синтезированы самой клеткой. Так, с помощью лизосомальных ферментов в клетках щитовидной железы гидролизуется тиреоглобулин, что приводит к образованию тиреоидных гормонов, которые затем выводятся в кровеносное русло путем экзоцитоза.
Пероксисомы
Пероксисомы (peroxysomae) в клетках тканей человека - это небольшие (размером 0,3-1,5 мкм) овальной формы тельца, ограниченные мембраной, содержащие гранулярный матрикс, в центре которого часто видны кри-сталлоподобные структуры, состоящие из фибрилл и трубок (сердцевина). Пероксисомы особенно характерны для клеток печени, почек. Во фракции пероксисом обнаруживаются ферменты окисления аминокислот, при работе которых образуется перекись водорода, а также выявляется фермент катала-за, разрушающий ее. Каталаза пероксисом играет важную защитную роль, так как Н2О2 является токсичным веществом для клетки.
Митохондрии
Митохондрии (mitochondrial) - энергетическая система клетки, органеллы синтеза АТФ. Их основная функция связана с окислением органических соединений и использованием освобождающейся при распаде этих соединений энергии для синтеза молекул АТФ. Исходя из этого, митохондрии часто называют энергетическими станциями клетки, или органеллами клеточного дыхания.
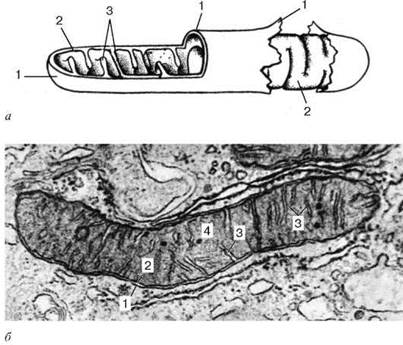
Ультрамикроскопическое строение митохондрии: а - схема; б - электронная микрофотография среза митохондрии печеночной клетки. 1 - наружная митохондриальная мембрана; 2 - внутренняя митохондриальная мембрана; 3 - кристы; 4 - митохондриальный матрикс.
Термин «митохондрия» был введен Бенда в 1897 г. для обозначения зернистых и нитчатых структур в цитоплазме разных клеток. Митохондрии можно наблюдать в живых клетках, так как они обладают достаточно высокой плотностью. Форма и размеры митохондрий животных клеток разнообразны, но в среднем толщина их около 0,5 мкм, а длина - от 1 до 10 мкм. Подсчеты показывают, что количество их в клетках сильно варьирует - от единичных элементов до сотен. Так, в клетке печени они составляют более 20 % общего объема цитоплазмы и содержат около 30-35 % общего количества белка в клетке. Площадь поверхности всех митохондрий печеночной клетки в 4-5 раз больше поверхности ее плазматической мембраны.
Обычно митохондрии скапливаются вблизи тех участков цитоплазмы, где возникает потребность в АТФ. Так, в сердечной мышце митохондрии находятся вблизи миофибрилл. В сперматозоидах митохондрии образуют спиральный футляр вокруг оси жгутика и т. п. Увеличение числа митохондрий в клетках происходит путем деления, или почкования, исходных митохондрий.
Митохондрии ограничены двумя мембранами толщиной около 7 нм.
Наружная митохондриальная мембрана (membrana mitochondrialis externa) отделяет их от гиалоплазмы. Обычно она имеет ровные контуры и замкнута, так что представляет собой мембранный мешок. Внешнюю мембрану от внутренней отделяет межмембранное пространство шириной около 10-20 нм. Внутренняя митохондриальная мембрана (membrana mitochondrialis interna) ограничивает собственно внутреннее содержимое митохондрии, ее матрикс (matrix mitochondrialis). Характерной чертой внутренних мембран митохондрий является их способность образовывать многочисленные выпячивания внутрь митохондрий. Такие выпячивания чаще всего имеют вид плоских гребней, или крист.
Матрикс митохондрий имеет тонкозернистое строение, в нем иногда выявляются тонкие нити (толщиной около 2-3 нм) и гранулы размером около 15-20 нм. Нити матрикса митохондрий представляют собой молекулы ДНК, а мелкие гранулы - митохондриальные рибосомы.
Основная функция митохондрий - синтез АТФ, происходящий в результате процессов окисления органических субстратов и фосфорилирования аденозиндифосфата (АДФ).
Выявлено, что в матриксе митохондрии локализуется автономная система митохондриального белкового синтеза. Она представлена молекулами ДНК, свободными от гистонов, что сближает их с ДНК бактериальных клеток. На этих ДНК происходит синтез молекул РНК разных типов: информационных, трансферных (транспортных) и рибосомных. В матриксе митохондрий наблюдается образование рибосом, отличных от рибосом цитоплазмы. Эти рибосомы участвуют в синтезе ряда митохондриальных белков, не кодируемых ядром.






