

Семя – орган полового размножения и расселения растений: наружи у семян имеется плотный покров – кожура...

Типы оградительных сооружений в морском порту: По расположению оградительных сооружений в плане различают волноломы, обе оконечности...
Семя – орган полового размножения и расселения растений: наружи у семян имеется плотный покров – кожура...
Типы оградительных сооружений в морском порту: По расположению оградительных сооружений в плане различают волноломы, обе оконечности...
Топ:
Методика измерений сопротивления растеканию тока анодного заземления: Анодный заземлитель (анод) – проводник, погруженный в электролитическую среду (грунт, раствор электролита) и подключенный к положительному...
Комплексной системы оценки состояния охраны труда на производственном объекте (КСОТ-П): Цели и задачи Комплексной системы оценки состояния охраны труда и определению факторов рисков по охране труда...
Устройство и оснащение процедурного кабинета: Решающая роль в обеспечении правильного лечения пациентов отводится процедурной медсестре...
Интересное:
Искусственное повышение поверхности территории: Варианты искусственного повышения поверхности территории необходимо выбирать на основе анализа следующих характеристик защищаемой территории...
Аура как энергетическое поле: многослойную ауру человека можно представить себе подобным...
Берегоукрепление оползневых склонов: На прибрежных склонах основной причиной развития оползневых процессов является подмыв водами рек естественных склонов...
Дисциплины:
|
из
5.00
|
Заказать работу |
|
|
|
|
Тема 1.2. Учение о клетке
Основой строения эукариотических организмов является наименьшая единица живого - клетка (cellula).
Клетка - это ограниченная активной мембраной, упорядоченная структурированная система биополимеров (нуклеиновые кислоты, белки, полисахариды, липиды) и их макромолекулярных комплексов, образующих ядро и цитоплазму, осуществляющих поддержание и воспроизведение всей системы в целом.
Кроме клеток, в организме находятся их производные: симпласт, синцитий, межклеточное вещество.
Содержимое клетки отделено от внешней среды плазматической мембраной (плазмолеммой).
Все эукариотические клетки состоят из двух основных компонентов: ядра и цитоплазмы.
В ядре различают хроматин (хромосомы), ядрышки, ядерную оболочку, нуклеоплазму (кариоплазму) и ядерный белковый остов (матрикс). Цитоплазма неоднородна по своему составу и строению и включает гиалоплазму (или основную плазму), в которой находятся органеллы; каждая из них выполняет обязательную функцию. Часть органелл имеет мембранное строение: эндоплазматическая сеть, комплекс Тольджи, лизосомы, пероксисомы и митохондрии. Немембранные органеллы цитоплазмы представлены рибосомами, клеточным центром, ресничками, жгутиками и компонентами цитоскелета. Кроме того, в гиалоплазме могут встретиться и иные необязательные структуры, или включения (жировые капли, пигментные гранулы и др.). Такое разделение клетки на отдельные компоненты не означает их структурной и функциональной обособленности. Все эти компоненты выполняют отдельные внутриклеточные функции, необходимые для существования клетки как целого, как элементарной единицы живого. Изучением общих черт строения и функционирования клеток занимается наука цитология, или, как ее теперь называют, биология клетки. Она исследует отдельные клеточные структуры, их участие в общеклеточных физиологических процессах, пути регуляции этих процессов, воспроизведение клеток и их органелл, приспособление клеток к условиям среды, реакции на действие различных факторов. Изучение цитологии имеет большое значение для медицины, так как практически все болезни человека являются результатом различных клеточных поражений или нарушений функций клеток в составе тканей различных органов.
|
|

Ультрамикроскопическое строение клетки животных организмов (схема): 1 - ядро; 2 - плазмолемма; 3 - микроворсинки; 4 - агранулярная эндоплазма-тическая сеть; 5 - гранулярная эндоплазматическая сеть; 6 - комплекс Гольджи; 7 - центриоль и микротрубочки клеточного центра; 8 - митохондрии; 9 - цито-плазматические пузырьки; 10 - лизосомы; 11 - микрофиламенты; 12 - рибосомы; 13 - выделение гранул секрета.
Клеточная теория
Клеточная теория - это обобщенное представление о строении клеток как единиц живого, об их воспроизведении и роли в формировании многоклеточных организмов.
Появлению и формулированию отдельных положений клеточной теории предшествовал довольно длительный (более 300 лет) период накопления знаний о строении различных одноклеточных и многоклеточных организмов, растений и позвоночных животных. Все это легло в основу сформулированной Т. Шванном (1838) клеточной теории строения организмов. Большую роль в развитии клеточной теории сыграли работы немецкого патолога Р. Вирхова.
В книге «Целлюлярная патология как учение, основанное на физиологической и патологической гистологии» (1855-1859) он обосновал фундаментальное положение о преемственности клеточного развития. Р. Вирхов, в противоположность Т. Шванну и М. Шлейдену, отстаивал взгляд на образование новых клеток не из «цитобластемы» - бесструктурной живой субстанции, а путем деления предсуществующих клеток (omnis cellula e cellula).
|
|
Создание клеточной теории и ее дальнейшее развитие стало важнейшим событием в биологии, одним из решающих доказательств единства происхождения всей живой природы. Клеточная теория оказала значительное влияние на развитие биологии и медицины, послужила главным фундаментом для становления таких дисциплин, как эмбриология, гистология. Принятие принципа клеточного строения организма оказало огромное влияние на физиологию, переведя ее на изучение реально функционирующих единиц - клеток. Она дала основы для научного понимания жизни, для понимания индивидуального развития и возникновения патологических изменений организмов.
Клеточная теория сохраняет свое значение до настоящего времени. Основные положения клеточной теории:
1. Клетка - наименьшая единица живого. Согласно одному из современных определений, живые организмы представляют собой открытые (т. е. обменивающиеся с окружающей средой веществами и энергией), саморегулирующиеся и самовоспроизводящиеся системы, важнейшими функционирующими компонентами которых являются белки и нуклеиновые кислоты. Все проявления жизни связаны с белками. Белки - функционирующие молекулы, обладающие сложной организацией и строгой функциональной специфичностью, которая определяется нуклеиновыми кислотами, несущими в себе информацию о строении тех или других белков. Живому свойствен ряд совокупных признаков: генетическая индивидуальность, способность к воспроизведению (репродукции), использование и трансформация энергии, метаболизм, реактивность и раздражимость, адаптивная изменчивость. Такую совокупность этих признаков впервые можно обнаружить только на клеточном уровне. Именно клетка как таковая является наименьшей единицей, обладающей всеми свойствами, отвечающими определению «живое».
2. Сходство клеток разных организмов по строению. Клетки могут иметь разнообразную внешнюю форму: шаровидную (лейкоциты), многогранную (клетки железистого эпителия), звездчатую и разветвленно-отростчатую (нервные и костные клетки), веретеновидную (гладкие мышечные клетки, фибробласты), столбчатую (кишечный эпителиоцит), уплощенную (эндо-телиоцит, мезотелиоцит) и др. Однако при изучении клеток тканей различных растений или животных обращает на себя внимание существование общего плана их организации (рис. 4.1). Такое сходство в строении клеток определяется общеклеточными функциями, связанными с поддержанием самой живой системы (синтез нуклеиновых кислот и белков, биоэнергетика клетки и др.). Одновременно это сходство указывает на общность происхождения всех эукариотических организмов.
Различие клеток в многоклеточном организме, обусловленное специализацией их функций, связано с развитием органелл специального значения. Так, если рассматривать мышечную клетку, то в ней, кроме общеклеточных структур (мембранные системы, рибосомы и др.), встречаются в большом количестве фибриллярные компоненты - миофиламенты и миофибриллы, обеспечивающие движение, сокращение. В нервной клетке, кроме общеклеточных компонентов, можно увидеть большое количество микротрубочек и промежуточных филаментов в клеточных отростках. Вся совокупность этих отличительных черт нервной клетки связана с ее специализацией - генерацией и передачей нервного импульса (подробно эти вопросы рассматриваются в разделе «Учение о тканях»).
|
|
3. Размножение клеток путем деления исходной клетки. Размножение прокариотических и эукариотических клеток происходит только путем деления исходной клетки, которому предшествует воспроизведение ее генетического материала (репликация ДНК). У эукариотических клеток единственно полноценным способом деления является митоз, или непрямое деление. При этом по двум дочерним клеткам распределяется равное количество хромосом, которые до этого удвоились в числе.
Митоз наблюдается у всех эукариотических (растительных и животных) клеток. Современная наука отвергает иные пути образования клеток и увеличения их числа в норме.
4. Клетки имеют одинаковый объем генетической информации. Это положение основано на том, что все клетки произошли от зиготы - одноклеточного зародыша. Однако морфологически и функционально клетки разных тканей значительно отличаются друг от друга. Несмотря на то, что потомки одноклеточного зародыша должны обладать одинаковыми генетическими потенциями, по мере развития зародыша его клетки все больше и больше отличаются друг от друга как по свойствам, так и по строению. Это связано с тем, что в разных клетках развивающегося организма одинаковая по объему генетическая информация реализуется не полностью (вследствие их детерминации и дифференциальной активности генов).
Индивидуальное развитие от одной клетки до многоклеточного зрелого организма - результат последовательного, избирательного включения работы разных генов в различных клетках. Это приводит к появлению клеток со специфическими для них структурами и особыми функциями, к процессу, называемому дифференцировкой. Дифференцировка обусловлена активностью разных генов в разных клетках, проявляемой по мере развития многоклеточного организма. Другими словами, сходство в строении клеток как данного организма, так и разных организмов определяется сходством общеклеточных функций, направленных на поддержание жизни самих клеток и их размножение. Разнообразие же в строении клеток - это результат их функциональной специализации, дифференцировки в процессе развития.
|
|
5. Клетки как части целостного организма. Каждое проявление деятельности целого организма, будь то реакция на раздражение или движение, иммунные реакции и многое другое, осуществляется специализированными клетками различных тканей. Однако, хотя клетка и является единицей функционирования в многоклеточном организме, деятельность ее не обособлена от других клеток и от межклеточного вещества. Специализированные клетки объединяются в системы тканей и органов, подчиненные и связанные межклеточными, тканевыми, гуморальными и нервными формами регуляции. Вот почему мы говорим об организме как о целом, а о клетках - как об элементарных единицах живого, специализированных на выполнении строго определенных функций, осуществляющих их в комплексе со всеми элементами, входящими в состав сложно организованной системы многоклеточного организма.
Цитоплазма
Цитоплазма (cytoplasma), часть клетки, отделенная от окружающей среды плазмолеммой, включает гиалоплазму и находящиеся в ней обязательные клеточные компоненты - органеллы, а также различные непостоянные структуры - включения.
Гиалоплазма
Гиалоплазма (от греч. hyalinos - прозрачный), или матрикс цитоплазмы, представляет собой очень важную часть клетки, ее истинную внутреннюю среду.
В электронном микроскопе матрикс цитоплазмы имеет вид гомогенного или тонкозернистого вещества с низкой электронной плотностью. Эта система способна переходить из золеобразного (жидкого) состояния в гелеобразное и обратно.
В организованной, упорядоченной многокомпонентной системе гиалоплазмы отдельные зоны могут менять свое агрегатное состояние в зависимости от условий или от функциональной задачи; в бесструктурной на взгляд гиалоплазме могут возникать и распадаться различные фибриллярные, нитчатые комплексы белковых молекул.
|
|
В состав гиалоплазмы входят главным образом различные глобулярные белки. Они составляют 20-25 % общего содержания белков в эукариотической клетке. К важнейшим ферментам гиалоплазмы относятся ферменты метаболизма сахаров, азотистых оснований, аминокислот, липидов и других важных соединений. В гиалоплазме располагаются ферменты активации аминокислот при синтезе белков, транспортные (трансферные) рибонуклеиновые кислоты (тРНК).
В гиалоплазме при участии рибосом и полирибосом (полисом) происходит синтез белков, необходимых для собственно клеточных нужд, для поддержания и обеспечения жизни данной клетки. Осмотические и буферные свойства клетки в значительной степени определяются составом и структурой гиалоплазмы.
Важнейшая роль гиалоплазмы заключается в том, что эта полужидкая среда объединяет все клеточные структуры и обеспечивает химическое взаимодействие их друг с другом. Через гиалоплазму осуществляется большая часть внутриклеточных транспортных процессов: перенос аминокислот, жирных кислот, нуклеотидов, сахаров. В ней идет постоянный поток ионов к плазматической мембране и от нее к митохондриям, к ядру и вакуолям.
В гиалоплазме происходит анаэробный синтез аденозинтрифосфата (АТФ) - гликолиз. Она является основным вместилищем и зоной перемещения массы молекул АТФ. В гиалоплазме происходит отложение запасных продуктов: гликогена, жировых капель, некоторых пигментов.
Органеллы
Органеллы - постоянно присутствующие и обязательные для всех клеток микроструктуры, выполняющие жизненно важные функции.
Эндоплазматическая сеть
Эндоплазматическая сеть (эндо-плазматический ретикулум) была открыта К. Р. Портером в 1945 г. Этот компонент вакуолярной системы клетки представляет собой совокупность вакуолей, плоских мембранных мешков или трубчатых образований, создающих трехмерную мембранную сеть. В состав сети входят гранулярные и агранулярные участки, которые могут чередоваться.
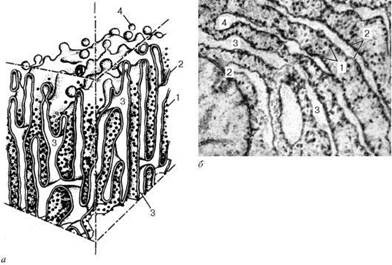
Строение гранулярной эндоплазматической сети: а - схема; б - электронная микрофотография участка среза эпителиальной клетки печени. 1 - рибосомы; 2 - пластинки; 3 - внутренние полости цистерн; 4 - отщепляющиеся мембранные пузырьки, лишенные рибосом.
Гранулярная эндоплазматическая сеть (reticulum endoplasmicum granulosum) на ультратонких срезах представлена замкнутыми мембранами, которые на сечениях образуют уплощенные мешки, цистерны, трубочки.
Диаметр цистерн значительно варьирует и в зависимости от функциональной активности клетки колеблется от 20 нм до несколько микрометров. Отличительной чертой мембран гранулярной эндоплазматической сети является то, что они со стороны гиалоплазмы покрыты многочисленными рибосомами.
Гранулярная эндоплазматическая сеть имеет разное строение. Для малоспециализированных клеток или для клеток с низкой метаболической активностью характерно наличие редких и разрозненных цистерн. Если возникают локальные скопления гранулярной эндоплазматической сети, то это свидетельствует об активном синтезе секреторных белков. Так, в клетках печени и некоторых нервных клетках гранулярная эндоплазматическая сеть собрана в отдельные зоны. В клетках поджелудочной железы гранулярная эндоплазмати-ческая сеть в виде плотно упакованных друг около друга мембранных цистерн занимает базальную и околоядерную зоны клетки. Рибосомы, связанные с мембранами эндоплазматической сети, участвуют в синтезе белков, выводимых из данной клетки («экспортируемые» белки). Кроме того, гранулярная эндоплазматическая сеть принимает участие в синтезе белков - ферментов, необходимых для организации внутриклеточного метаболизма, а также используемых для внутриклеточного пищеварения.
Белки, накапливающиеся в полостях эндоплазматической сети, могут, минуя гиалоплазму, транспортироваться в вакуоли комплекса Гольджи, где они модифицируются и входят в состав либо лизосом, либо секреторных гранул, содержимое которых остается изолированным от гиалоплазмы мембраной. Внутри канальцев или вакуолей гранулярной эндоплазматической сети происходит модификация белков, например, связывание их с сахарами (первичное глюкозилирование).
В гранулярной эндоплазматической сети на ее рибосомах происходит синтез мембранных интегральных белков, которые встраиваются в толщу мембраны. Здесь же со стороны гиалоплазмы идет синтез липидов и их встраивание в мембрану. В результате этих двух процессов наращиваются сами мембраны эндоплазматической сети и другие компоненты вакуоляр-ной системы клетки.
Агранулярная (гладкая) эндоплазматическая сеть (reticulum endoplasmaticum nongranulosum) также представлена мембранами, образующими мелкие вакуоли, трубки, канальцы, которые могут ветвиться, сливаться друг с другом. В отличие от гранулярной эндоплазматической сети на мембранах гладкой эндоплазматической сети нет рибосом. Диаметр вакуолей и канальцев гладкой эндоплазматической сети обычно около 50-100 нм. Гладкая эндоплаз-матическая сеть возникает и развивается на основе гранулярной эндоплаз-матической сети. В отдельных участках гранулярной эндоплазматической сети образуются новые липопротеидные мембранные участки, лишенные рибосом. Эти участки могут разрастаться, отщепляться от гранулярных мембран и функционировать как самостоятельная вакуолярная система.
Деятельность гладкой эндоплазматической сети связана с метаболизмом липидов и некоторых внутриклеточных полисахаридов. Гладкая эндоплазматическая сеть участвует в заключительных этапах синтеза липидов. Она сильно развита в клетках, секретирующих стероиды, например, в эндокринных клетках коркового вещества надпочечников, в эпителиальных клетках извитых семенных канальцев.
Тесная топографическая связь гладкой эндоплазматической сети с отложениями гликогена (запасной внутриклеточный полисахарид животных) в гиалоплазме различных клеток (клетки печени, мышечные волокна) указывает на ее возможное участие в метаболизме углеводов.
В поперечнополосатых мышечных волокнах гладкая эндоплазматиче-ская сеть способна депонировать ионы кальция, необходимые для функции мышечной ткани.
Очень важна роль гладкой эндоплазматической сети в дезактивации различных вредных для организма веществ за счет их окисления с помощью ряда специальных ферментов. Особенно четко она проявляется в клетках печени. Так, при некоторых отравлениях в клетках печени появляются ацидофильные зоны (не содержащие РНК), сплошь заполненные гладким эндо-плазматическим ретикулумом.
Комплекс Гольджи
Комплекс Гольджи (пластинчатый комплекс) был открыт в 1898 г. К. Гольджи. Автор, используя свойства связывания тяжелых металлов (осмия или серебра) с клеточными структурами, выявил в нервных клетках сетчатые образования, которые он назвал внутренним сетчатым аппаратом (apparatus reticularis internus). В дальнейшем его стали называть аппаратом, или комплексом Гольджи (complexus Golgiensis). Подобные структуры затем были описаны во всех клетках эукариот.
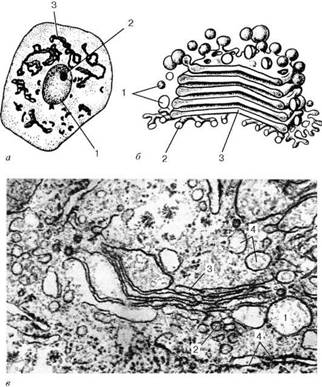
Комплекс Гольджи: а - нервная клетка спинного мозга, импрегнация серебром по методу Гольджи: 1 - ядро; 2 - ядрышко; 3 - комплекс Гольджи; б - схема ультрамикроскопического строения (трехмерная реконструкция); в - комплекс Гольджи на ультратонком срезе (печеночная клетка): 1 - пузырьки; 2 - трубочки; 3 - уплощенные мешочки (цистерны); 4 - фрагменты гранулярной эндоплазматической сети
Отдельная зона скопления этих мембран называется диктиосомой (стопкой Гольджи). Таких зон в клетке может быть несколько. В диктиосоме плотно друг к другу (на расстоянии 20-25 нм) расположены 5-10 плоских цистерн, между которыми находятся тонкие прослойки гиалоплазмы. Каждая цистерна имеет переменную толщину: в центре ее мембраны могут быть сближены (до 25 нм), а на периферии иметь расширения - ампулы, ширина которых непостоянна. Кроме плотно расположенных плоских цистерн, в зоне комплекса Гольджи наблюдается множество мелких пузырьков (везикул), которые встречаются главным образом в его периферических участках. Иногда они отшнуровываются от ампулярных расширений на краях плоских цистерн. В зоне диктиосомы различают проксимальную (cis) и дистальную (trans) поверхности. В секретирующих клетках обычно комплекс Гольджи поляризован: его проксимальная поверхность обращена к ядру, в то время как дистальная - к поверхности клетки.
В клетках отдельные диктиосомы могут быть связаны друг с другом системой везикул и цистерн, примыкающих к дистальной поверхности, так что образуется рыхлая трехмерная сеть, выявляемая в световом и электронном микроскопах («транс-сеть» комплекса Гольджи).
Комплекс Гольджи участвует в сегрегации и накоплении продуктов, синтезированных в эндоплазматической сети, в их химической перестройке, созревании; в его цистернах происходят синтез полисахаридов, их комплексирование с белками, что приводит к образованию пептидогликанов. С помощью комплекса Гольджи осуществляется процесс выведения готовых секретов за пределы секреторной клетки. Кроме того, комплекс Гольджи обеспечивает формирование лизосом. Внутри полостей комплекса Гольджи с помощью различных ферментов (транс-глюкозидаз) по разному модифицируются лизосомные белки и белки секретов: происходят последовательная замена и наращивание олигосахаридных цепочек.
В цистернах дистальной (trans) поверхности происходит сортировка белков: на внутренних поверхностях мембран цистерн располагаются рецепторы, узнающие или секреторные белки, или белки, входящие в состав лизосом (гидролазы). В результате от цистерн дистальной поверхности диктиосом отщепляются два типа мелких вакуолей: а) содержащие гидролазы - лизосомы (первичные); б) секреторные белки.
Секреторная функция комплекса Гольджи заключается в том, что синтезированный на рибосомах белок, накапливающийся внутри цистерн эндо-плазматической сети, транспортируется далее в вакуоли комплекса Гольджи.
Лизосомы
Лизосомы (lysosomae) - это разнообразный класс вакуолей размером 0,2- 0,4 мкм, ограниченных одиночной мембраной. Характерным признаком лизосом является наличие в них гидролитических ферментов - гидролаз (протеиназы, нуклеазы, фосфатазы, липазы и др.), расщепляющих различные биополимеры при кислом значении рН. Лизосомы были открыты в 1949 г. де Дювом.
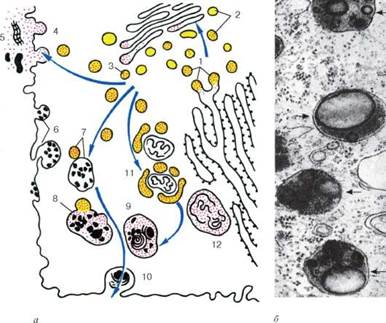
Строение лизосом: а - схема участия структур клетки в образовании лизосом и во внутриклеточном пищеварении: 1 - образование из гранулярной эндоплазматической сети мелких пузырьков, содержащих гидролитические ферменты; 2 - перенос ферментов в комплекс Гольджи; 3 - образование первичных лизосом; 4 - выделение и использование (5) гидролаз при внеклеточном расщеплении; 6 - эндоцитозные пузырьки; 7 - слияние первичных лизосом и эндоцитозных пузырьков; 8 - образование вторичных лизосом; 9 - телолизосомы; 10 - экскреция остаточных телец; 11 - слияние первичных лизосом с разрушающимися структурами клетки; 12 - аутофаголизосома; б - электронная микрофотография среза гетерофаголизо-сом (обозначены стрелками)
Кроме собственно лизосом (первичных) различают аутофаголизосомы, или гетеролизосомы (вторичные лизосомы), и телолизосомы (остаточные тельца).
Разнообразие морфологии лизосом объясняется тем, что эти частицы участвуют в процессах внутриклеточного переваривания, образуя сложные пищеварительные вакуоли как экзогенного (внеклеточного), так и эндогенного (внутриклеточного) происхождения.
Лизосомы (первичные) представляют собой мелкие мембранные пузырьки размером около 0,2-0,5 мкм, заполненные бесструктурным веществом, содержащим гидролазы, в том числе активную кислую фосфатазу, которая является маркерным ферментом для лизосом. Эти мелкие пузырьки практически очень трудно отличить от мелких везикул на периферии зоны комплекса Гольджи, которые также содержат кислую фосфатазу. Местом ее синтеза является гранулярная эндоплазматическая сеть. Затем этот фермент появляется в цистернах проксимальной поверхности диктиосомы, а затем в мелких везикулах по периферии диктиосомы и, наконец, в лизосомах. Таким образом, весь путь образования лизосом очень сходен с образованием секреторных (зимогенных) гранул в клетках поджелудочной железы, за исключением последнего этапа.
Гетерофаголизосомы (вторичные лизосомы), или внутриклеточные пищеварительные вакуоли, формируются при слиянии лизосом с фагоцитарными или пиноцитозными вакуолями. Если происходит слияние лизосомы с измененными органеллами самой клетки, то такая структура называется аутофаголизосома. При этом ферменты лизосомы получают доступ к субстратам, которые они и начинают расщеплять. Вещества, попавшие в состав гетероили аутофаголизосом (вторичных лизосом), расщепляются гидролазами до мономеров, которые транспортируются через мембрану лизосомы в гиалоплазму, где они реутилизируются, т. е. включаются в различные обменные процессы.
Однако расщепление, переваривание макромолекул лизосомой может идти в ряде клеток не до конца. В этом случае в вакуолях лизосом накапливаются непереваренные продукты. Такая органелла носит название тело-лизосома, или остаточное тельце (corpusculum residuale). Остаточные тельца содержат меньше гидролитических ферментов, в них происходит уплотнение содержимого, его перестройка. Часто в остаточных тельцах наблюдается вторичная структуризация неперевариваемых липидов, которые образуют слоистые структуры. Там же откладываются пигментные вещества. Например, у человека при старении организма в клетках мозга, печени и в мышечных волокнах в телолизосомах происходит отложение «пигмента старения» - липофусцина.
При участии лизосом (аутофаголизосом) может происходить модификация продуктов, которые синтезированы самой клеткой. Так, с помощью лизосомальных ферментов в клетках щитовидной железы гидролизуется тиреоглобулин, что приводит к образованию тиреоидных гормонов, которые затем выводятся в кровеносное русло путем экзоцитоза.
Пероксисомы
Пероксисомы (peroxysomae) в клетках тканей человека - это небольшие (размером 0,3-1,5 мкм) овальной формы тельца, ограниченные мембраной, содержащие гранулярный матрикс, в центре которого часто видны кри-сталлоподобные структуры, состоящие из фибрилл и трубок (сердцевина). Пероксисомы особенно характерны для клеток печени, почек. Во фракции пероксисом обнаруживаются ферменты окисления аминокислот, при работе которых образуется перекись водорода, а также выявляется фермент катала-за, разрушающий ее. Каталаза пероксисом играет важную защитную роль, так как Н2О2 является токсичным веществом для клетки.
Митохондрии
Митохондрии (mitochondrial) - энергетическая система клетки, органеллы синтеза АТФ. Их основная функция связана с окислением органических соединений и использованием освобождающейся при распаде этих соединений энергии для синтеза молекул АТФ. Исходя из этого, митохондрии часто называют энергетическими станциями клетки, или органеллами клеточного дыхания.
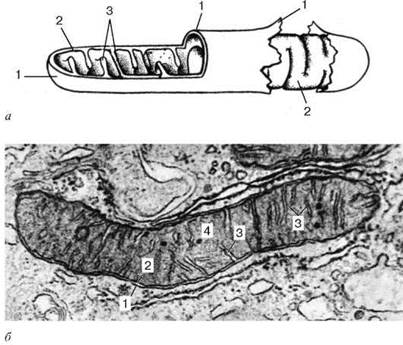
Ультрамикроскопическое строение митохондрии: а - схема; б - электронная микрофотография среза митохондрии печеночной клетки. 1 - наружная митохондриальная мембрана; 2 - внутренняя митохондриальная мембрана; 3 - кристы; 4 - митохондриальный матрикс.
Термин «митохондрия» был введен Бенда в 1897 г. для обозначения зернистых и нитчатых структур в цитоплазме разных клеток. Митохондрии можно наблюдать в живых клетках, так как они обладают достаточно высокой плотностью. Форма и размеры митохондрий животных клеток разнообразны, но в среднем толщина их около 0,5 мкм, а длина - от 1 до 10 мкм. Подсчеты показывают, что количество их в клетках сильно варьирует - от единичных элементов до сотен. Так, в клетке печени они составляют более 20 % общего объема цитоплазмы и содержат около 30-35 % общего количества белка в клетке. Площадь поверхности всех митохондрий печеночной клетки в 4-5 раз больше поверхности ее плазматической мембраны.
Обычно митохондрии скапливаются вблизи тех участков цитоплазмы, где возникает потребность в АТФ. Так, в сердечной мышце митохондрии находятся вблизи миофибрилл. В сперматозоидах митохондрии образуют спиральный футляр вокруг оси жгутика и т. п. Увеличение числа митохондрий в клетках происходит путем деления, или почкования, исходных митохондрий.
Митохондрии ограничены двумя мембранами толщиной около 7 нм.
Наружная митохондриальная мембрана (membrana mitochondrialis externa) отделяет их от гиалоплазмы. Обычно она имеет ровные контуры и замкнута, так что представляет собой мембранный мешок. Внешнюю мембрану от внутренней отделяет межмембранное пространство шириной около 10-20 нм. Внутренняя митохондриальная мембрана (membrana mitochondrialis interna) ограничивает собственно внутреннее содержимое митохондрии, ее матрикс (matrix mitochondrialis). Характерной чертой внутренних мембран митохондрий является их способность образовывать многочисленные выпячивания внутрь митохондрий. Такие выпячивания чаще всего имеют вид плоских гребней, или крист.
Матрикс митохондрий имеет тонкозернистое строение, в нем иногда выявляются тонкие нити (толщиной около 2-3 нм) и гранулы размером около 15-20 нм. Нити матрикса митохондрий представляют собой молекулы ДНК, а мелкие гранулы - митохондриальные рибосомы.
Основная функция митохондрий - синтез АТФ, происходящий в результате процессов окисления органических субстратов и фосфорилирования аденозиндифосфата (АДФ).
Выявлено, что в матриксе митохондрии локализуется автономная система митохондриального белкового синтеза. Она представлена молекулами ДНК, свободными от гистонов, что сближает их с ДНК бактериальных клеток. На этих ДНК происходит синтез молекул РНК разных типов: информационных, трансферных (транспортных) и рибосомных. В матриксе митохондрий наблюдается образование рибосом, отличных от рибосом цитоплазмы. Эти рибосомы участвуют в синтезе ряда митохондриальных белков, не кодируемых ядром.
Рибосомы
Рибосомы ( ribosomae) - элементарные аппараты синтеза белковых, полипептидных молекул - обнаруживаются во всех клетках. Рибосомы - это сложные рибонуклеопротеиды, в состав которых входят белки и молекулы рибосо-мальных РНК (рРНК) примерно в равных весовых отношениях. Размер функционирующей рибосомы эука риотических клеток 25x20x20 нм. Такая рибосома состоит из большой и малой субъединиц. Каждая из субъединиц построена из рибонуклеопро-теидного тяжа, где рРНК взаимодействует с разными белками и образует тело рибосомы.
Различают единичные рибосомы и комплексы рибосом (полисомы). Рибосомы могут располагаться свободно в гиалоплазме или быть связанными с мембранами эндоплазматической сети. В малоспециализированных и быстрорастущих клетках в основном обнаруживаются свободные рибосомы. В специализированных клетках рибосомы располагаются в составе гранулярной эндоплазматической сети. Синтетическая деятельность свободных рибосом направлена в основном на собственные нужды клетки. Связанные рибосомы обеспечивают синтез белков «на экспорт», т. е. на обеспечение нужд организма. Содержание РНК и соответственно степень белковых синтезов коррелируют с интенсивностью базофилии цитоплазмы, т. е. со способностью окрашиваться основными красителями.
Цитоскелет
Цитоскелет (cytoskeleton) - опорно-двигательная система клетки, включающая немембранные белковые нитчатые органеллы, выполняющие как каркасную, так и двигательную функции в клетке. Эти структуры являются динамическими образованиями, они могут быстро возникать в результате полимеризации их элементарных молекул и так же быстро разбираться, исчезать при деполимеризации. К этой системе относятся фибриллярные структуры и микротрубочки.
Фибриллярные структуры цитоплазмы. К фибриллярным компонентам цитоплазмы эукариотических клеток относятся микрофиламенты (micro-filamenti) толщиной 5-7 нм и так называемые промежуточные филаменты (filamenti intermedii) толщиной около 10 нм.
Микрофиламенты встречаются практически во всех типах клеток. Они располагаются в кортикальном слое цитоплазмы, непосредственно под плазмолеммой, пучками или слоями. Их можно видеть в псевдоподиях амеб или в движущихся отростках фибробластов, в микроворсинках кишечного эпителия. Микрофиламенты часто образуют пучки, направляющиеся в клеточные отростки.
С помощью иммунофлюоресцентных методов показано, что в состав микрофиламентов кортикального слоя и пучков входят белки: актин, миозин, тропомиозин, альфа-актинин. Следовательно, микрофиламенты не что иное, как внутриклеточный сократительный аппарат, обеспечивающий не только подвижность клеток при активном амебоидном их перемещении, но, вероятно, и большинство внутриклеточных движений, таких как токи цитоплазмы, движение вакуолей, митохондрий, деление клетки. Кроме того, актиновые микрофиламенты играют и каркасную роль. Соединяясь с рядом стабилизирующих белков, они могут образовывать временные или постоянные (как в микроворсинках кишечного эпителия) пучки или сети, играющие большую роль в структурировании цитоплазмы.
Промежуточные филаменты. Это тонкие (10 нм) неветвящиеся, часто располагающиеся пучками нити. Характерно, что в клетках разных тканей их белковый состав различен. Например, в эпителии кожного типа в состав промежуточных филаментов входит кератин. Пучки кератиновых промежуточных филаментов в эпителиальных клетках образуют тонофиламенты, которые подходят к десмосомам. В состав промежуточных филаментов клеток, производных мезенхимы (например, фибробластов), входит другой белок - виментин; в мышечных клетках обнаруживается десмин; в нервных клетках в состав нейрофиламентов также входит особый белок. Роль промежуточных микрофиламентов, скорее всего, опорно-каркасная; эти фибриллярные структуры не так лабильны, как микротрубочки и микрофиламенты.
Микротрубочки (microtubuli). В клетках микротрубочки принимают участие в создании ряда временных (цитоскелет интерфазных клеток, веретено деления) или постоянных (центриоли, реснички, жгутики) структур.

Строение микротрубочек: а - Тб-субъединица, димер тубу-лина в составе микротрубочек; б - микротрубочки в цитоплазме клетки (стрелки).
Микротрубочки представляют собой прямые, неветвящиеся длинные полые цилиндры. Их внешний диаметр составляет около 24 нм, внутренний просвет имеет ширину 15 нм, а толщина стенки - 5 нм. Стенка микротрубочек построена за счет плотно уложенных округлых субъединиц диаметром около 5 нм. В электронном микроскопе на поперечных сечениях микротрубочек видны большей частью 13 субъединиц, выстроенных в виде однослойного кольца. Микротрубочки, выделенные из разных источников (реснички простейших, клетки нервной ткани, веретено деления), имеют сходный состав и содержат белки - тубулины.
Клеточный центр
Клеточный центр (центросома) состоит из центриолей и связанных с ними микротрубочек - центросферы. Термин «центриоли» был предложен Т. Бо-вери в 1895 г. для обозначения очень мелких телец, размер которых находится на границе разрешающей способности светового микроскопа. В некоторых объектах удавалось видеть, что мелкие плотные тельца - центриоли (centriolum) окружены зоной более светлой цитоплазмы, от которой радиально отходят тонкие фибриллы. Эти органеллы в делящихся клетках принимают участие в формировании веретена деления и располагаются на его полюсах. В неделящихся клетках центриоли часто определяют полярность клеток эпителия и располагаются вблизи комплекса Гольджи.
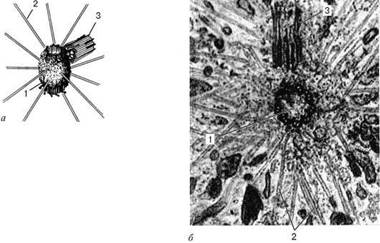
Строение клеточного центра в полюсе митотического веретена клетки: а - схема; б - электронная микрофотография. 1 - активная материнская центриоль, окруженная тонкофибриллярным матриксом, от которого отходят микротрубочки полярной лучистости (2); 3 - неактивная дочерняя центриоль
Основой строения центриоли являются расположенные по окружности 9 триплетов микротрубочек, образующих таким образом полый цилиндр. Его диаметр составляет около 0,2 мкм, а длина - 0,3- 0,5 мкм (хотя встречаются центриоли, достигающие в длину нескольких микрометров).
Обычно в интерфазных клетках присутствуют две центриоли - рядом друг с другом, образующие диплосому (diplosoma). В диплосоме центриоли располагаются под прямым углом по отношению друг к другу. Из двух центриолей различают материнскую и дочернюю. Обе центриоли сближены, конец дочерней центриоли направлен к поверхности материнской центриоли.
Вокруг каждой центриоли расположен бесструктурный, или тонковолокнистый, матрикс. Часто можно обнаружить несколько дополнительных структур, связанных с центриолями: спутники (сателлиты), фокусы схождения микротрубочек, дополнительные микротрубочки, образующие особую зону - центросф<
|
|
|
Типы оградительных сооружений в морском порту: По расположению оградительных сооружений в плане различают волноломы, обе оконечности...

Индивидуальные и групповые автопоилки: для животных. Схемы и конструкции...

История развития пистолетов-пулеметов: Предпосылкой для возникновения пистолетов-пулеметов послужила давняя тенденция тяготения винтовок...

Индивидуальные очистные сооружения: К классу индивидуальных очистных сооружений относят сооружения, пропускная способность которых...
© cyberpedia.su 2017-2024 - Не является автором материалов. Исключительное право сохранено за автором текста.
Если вы не хотите, чтобы данный материал был у нас на сайте, перейдите по ссылке: Нарушение авторских прав. Мы поможем в написании вашей работы!