КУРСОВАЯ РАБОТА
НА ТЕМУ: "КОМПЛЕКСНАЯ ОЦЕНКА ЖЕЛЕЗОДЕФИЦИТНЫХ СОСТОЯНИЙ"
Выполнила: студентка 3 курса
факультета естественных наук
специальности "Лабораторная диагностика"
Шаповалова И.И.
Луганск, 2014
Содержание
Введение
1. Железосодержащие органические соединения в организме человека
1.1 Клеточное железо
1.2 Внеклеточное железо
2. Медико-биологические основы обеспечения организма железом
2.1 Эритропоэз
2.1.1 Эритроидный ряд клеток.
2.1.2 Регуляция эритропоэза
2.2 Синтез гемоглобина
2.3 Регенерация красной крови
2.4 Метаболизм железа
2.4.1 Обмен железа
2.4.2 Этапы обмена железа в организме
2.4.3 Потери железа
2.4.4 Регуляция метаболизма железа
3. Клинико-лабораторные характеристики красной крови и обмена железа
4. Железодефицитные состояния
4.1 Определение
4.2 Этиология и патогенез железодефицитных состояний
4.2.1 Алиментарная недостаточность
4.2.2 Нарушение усвоения железа
4.2.3 Кровопотери
4.2.4 Перераспределение и конкурентное потребление железа
4.2.5 Нарушения транспорта железа
4.3 Клиническая картина железодефицитных состояний
4.3.1 Сидеропения (гипосидероз)
4.3.2 Анемический синдром.
4.4 Диагностика железодефицитных состояний
4.4.1 Доказательство наличия анемии
4.4.2 Доказательство наличия латентного дефицита железа
4.4.3 Доказательство гипохромности анемии
4.4.4 Доказательство дефицита железа, исключение нежелезодефицитных причин гипохромии и микроцитоза
4.4.5 Определение причины дефицита железа
Заключение
Список использованной литературы
Введение
Для нормального роста и выполнения биологических функций человеку и животным кроме витаминов необходим целый ряд неорганических элементов. Эти элементы можно разделить на две класса. Один из них - макроэлементы, а другой - микроэлементы.
Макроэлементы, к которым относятся кальций, магний, натрий, калий, фосфор, сера и хлор, требуются организму в относительно больших количествах (порядка нескольких граммов в сутки). Часто они выполняют более чем одну функцию.
Более непосредственное отношение к действию ферментов имеют независимые микроэлементы, суточная потребность в которых не превышает нескольких миллиграммов, то есть, сопоставима с потребностью в витаминах.
Известно, что в пище животных обязательно должно содержаться около 15 микроэлементов, большинство из которых выполняет, по меньшей мере, какую-нибудь одну из трёх возможных функций. Во-первых, незаменимый микроэлемент сам по себе может обладать каталитической активностью по отношению к той или иной химической реакции, скорость которой значительно возрастает в присутствии ферментного белка. Это особенно характерно для ионов железа и меди. Во-вторых, ион металла может образовывать комплекс одновременно с активным центром фермента, в результате чего происходит их сближение и переход в активную форму. И, в-третьих, ион металла может играть роль мощного акцептора электронов на определённой стадии.
Железо относится к тем микроэлементам, биологические функции которых изучены наиболее полно. Значение железа для организма человека трудно переоценить. Подтверждением этому может быть не только большая его распространенность в природе, но и важная роль в сложных процессах, происходящих в живом организме. Биологическая ценность железа определяется многократностью его функций, незаменимостью другими металлами в сложных биохимических процессах, активным участием в клеточном дыхании, обеспечивающем нормальное функционирование тканей и организма человека.
Железо является одним из важнейших элементов в организме человека и входит в состав многих субстратов и ферментов, отвечающих за транспорт кислорода к клеткам, функционирование дыхательной цепи митохондрий, окислительно-восстановительные клеточные реакции, антиоксидантную защиту, функционирование нервной и иммунной систем и др.
Клеточное железо
Гемоглобин, содержащийся в эритроцитах, выполняет важную для организма газотранспортную функцию - переносит экзогенный кислород и эндогенный углекислый газ. Эритроцит по отношению к гемоглобину играет роль буферной системы, способной регулировать общую величину газотранспортной функции. Дыхательный пигмент крови - сложный белок, состоящий из белковой молекулы - глобина, соединенной полипептидными цепочками с 4 комплексами гема. Глобин состоит из 2 пар полипептидных цепочек, каждая из которых содержит 141-146 аминокислот. Гем, составляющий 4% веса молекулы гемоглобина, содержит железо в центре порфиринового кольца. У здорового человека гемоглобин гетерогенен. Нормальный эритроцит содержит приблизительно 30 пг. гемоглобина, в котором находится 0,34% железа.
Миоглобин - дыхательный белок сердечной и скелетной мускулатуры. Он состоит из единственной полипептидной цепочки, содержащей 153 аминокислоты и соединенный с гемпростетической группой. Основной функцией миоглобина является транспортировка кислорода через клетку и регуляция его содержания в мышце для осуществления сложных биохимических процессов, лежащих в основе клеточного дыхания. Он содержит 0,34% железа. Миоглобин депонирует кислород во время сокращения мышц, а при их поражении он может попадать в кровь и выделяться с мочой.
Железосодержащие ферменты и негеминовое железо клетки находится главным образом в митохондриях. Наиболее изученными и важными для организма ферментами являются цитохромы, каталаза и пероксидаза.
Цитохромы делятся на 4 группы в зависимости от строения геминовой группы:
А - цитохромы с гем - группой, соединяющей формилпорфин;
В - цитохромы с протогем - группой;
С - цитохромы с замещенной мезогем - группой;
Д - цитохромы с гем - группой, соединяющей дегидропорфин.
В организме человека содержатся следующие цитохромы:
а1, аз, в, в5, с, с1, Р450. Они представляют собой липидные комплексы гемопротеинов и прочно связаны с мембраной митохондрии. Однако, цитохромы в5 и Р450 находятся в эндоплазматическом ретикулюме, а микросомы содержат НАДН - цитохром С - редуктазу. Существует мнение, что митохондриальное дыхание необходимо для процессов дифференцировки тканей, а внемитохондриальное играет важную роль в процессах роста и дыхания клетки. Основной биологической ролью большинства цитохромов является участие в переносе электронов, лежащих в основе процессов терминального окисления в тканях.
Цитохромоксидаза является конечным ферментом митохондриального транспорта электронов - электронотранспортной цепочки, ответственным за образование АТФ при окислительном фосфолировании в митохондриях. Показана тесная зависимость между содержанием этого фермента в тканях и утилизацией ими кислорода. Каталаза, как и цитохромоксидаза, состоит из единственной полипептидной цепочки, соединенной с гем - группой. Она является одним из важнейших ферментов, предохраняющих эритроциты от окислительного гемолиза. Каталаза выполняет двойную функцию в зависимости от концентрации перекиси водорода в клетке. При высокой концентрации перекиси водорода фермент катализирует реакцию ее разложения, а при низкой - и в присутствии донора водорода (метанол, этанол и др.) становится преобладающей пероксидазная активность каталазы.
Пероксидаза содержится преимущественно в лейкоцитах и слизистой тонкого кишечника у человека. Она также обладает защитной ролью, предохраняя клетки от их разрушения перекисными соединениями.
Миелопероксидаза - железосодержащий геминовый фермент, находящийся в азурофильных гранулах нейтрофильных лейкоцитов и освобождается в фагоцитирующие вакуоли в течение лизиса гранул. Активированное этим ферментом разрушение белка клеточной стенки бактерий является смертельным для микроорганизма, а активированное им йодинирование частиц относится к бактерицидной функции лейкоцитов.
К железосодержащим относятся ифлавопротеиновые ферменты, в которых железо не включено в геминовую группу и необходимо только для реакций переноса.
Наиболее изученной является сукцинатдегидрогеназа, которая наиболее активна в цикле трикарбоновых кислот. Митохондриальные мембраны свободно проницаемы для субстрата фермента.
железодефицитное состояние дефицит железо
Негеминовое железо, локализующееся главным образом в митохондриях клетки, играет существенную роль в дыхании клетки, участвуя в окислительном фосфолировании и транспорте электронов при терминальном окислении, в цикле трикарбоновых кислот.
Ферритин и гемосидерин - запасные соединения железа в клетке, находящиеся главным образом в ретикулоэндотелиальной системе печени, селезенки и костного мозга. Приблизительно одна треть резервного железа организма человека, преимущественно в виде ферритина, падает на долю печени. Запасы железа могут быть при необходимости мобилизованы для нужд организма и предохраняют его от токсичного действия свободно циркулирующего железа.
Известно, что гепатоциты и купферовские клетки печени участвуют в создании резервного железа, причем в нормальной печени большая часть нового железа обнаружена в гепатоцитах в виде ферритина. При парентеральном введении железа как гепатоциты, так и купферовские клетки печени аккумулируют большое количество дополнительного ферритина, хотя последние имеют тенденцию запасать относительно больше из лишнего негеминового железа в виде гемосидерина.
Сферическая белковая оболочка молекулы ферритина состоит из 24 субъединиц, имеющих молекулярный вес 18500 - 19000. Общий молекулярный вес апоферритина 445000. Электронно-микроскопические исследования показали, что ферритин имеет полую оболочку с внутренним диаметром 70 - 80 А. Оболочка имеет 6 каналов, расширяющихся внутри (их диаметр 9-12 А).
Ядро ферритина состоит из мицелл железо-фосфатного комплекса, имеющих кристаллическую структуру. Захват и освобождение железа осуществляется через белковые каналы путем свободного пассажа, а его отложение и мобилизация происходят на поверхности микрокристаллов. Стимуляция синтеза ферритина железом является хорошо установленным фактом.
Как известно, печень является основным компонентом ретикулоэндотелиальной системы. В конце жизнедеятельности эритроциты фагоцитируются макрофагами этой системы, а освобождающееся железо или оседает в печени в виде ферритина (гемосидерина), или возвращается в плазму крови и захватывается в паренхиматозных клетках печени и мышц, а также в макрофагах ретикулоэндотелиальной системы печени, селезенки и костного мозга.
Гемосидерин является вторым запасным соединением железа в клетке и содержит значительно больше железа, чем ферритин. В отличие от ферритина он нерастворим в воде. Существует достаточно аргументированное предположение, что преобразование ферритина в гемосидерин происходит путем постепенного перенасыщения ферритиновой молекулы железом с последующим ее разрушением и образованием зрелого гемосидерина.
Внимание исследователей в последнее время привлекает циркулирующий в крови ферритин. Вероятно, он происходит из клеток ретикулоэндотелиальной системы. Имеются предположения, что сывороточный ферритин является отражением активной секреции ферритина из печеночных клеток, возможно из
связанных полисом. Таким образом, его присутствие в сыворотке в небольшом количестве не является результатом разрушения клеток печени. Не только его происхождение, но и биологическая роль в организме человека до настоящего времени изучены недостаточно. Не вызывает сомнений точно установленный факт концентрация сывороточного ферритина отражает состояние запасного фонда железа в организме человека. Отметим, что хорошая зависимость отмечена между уровнем сывороточного ферритина и мобилизуемыми запасами железа в организме человека, изученных с помощью количественных кровопусканий, а также между ферритином и концентрацией негеминового железа в тканях печени, полученных с помощью биопсии у людей. Средняя концентрация его в сыворотке крови у мужчин выше, чем у женщин, с колебаниями от 12 до 300 мкг/л.
Внеклеточное железо
Во внеклеточных жидкостях железо находится в связанном состоянии - в виде железо - белковых комплексов. Концентрация его в плазме широко варьирует у здорового человека, составляет 10,8 - 28,8 мкмоль/л. с достаточно большими суточными колебаниями, достигающими 7,2 мкмоль/л. Общее содержание железа во всем объеме циркулирующей плазмы у взрослого человека составляет 3 - 4 мг. Уровень железа в плазме крови зависит от ряда факторов: взаимоотношения процессов разрушения и образования эритроцитов, состояния запасного фонда железа в желудочно - кишечном тракте. Однако наиболее важной причиной, определяющей уровень плазменного железа, является взаимодействие процессов синтеза и распада эритроцитов.
Железо-связывающий белок трансферрин, открытый шведскими учеными, содержится в небольшом количестве в плазме крови. Общая железо-связывающая способность плазмы, характеризующаяся практически концентрацией трансферрина, колеблется от 44,7 до 71,6 мкмоль/л, а свободная железо-связывающая способность - резервная емкость трансферрина - составляет 28.8 - 50.4 мкмоль/л у здорового человека.
В плазме здорового человека трансферрин может находиться в 4 молекулярных формах:
) апотрансферрина;
) моножелезистого трансферрина А - железо занимает только А - пространство;
) моножелезистого трансферрина В - железо занимает только В-пространство;
) дижелезистого транферрина - заняты А и В пространства.
Молекулярный вес трансферрина 76000 - 80000, он состоит из единственной полипептидной цепочки с расположенными на ней двумя значительно схожими, если не идентичными, металлсвязывающими пространствами. Эти пространства (А и В) наиболее прочно связывают железо по сравнению с ионами других металлов. Около 6% железо-связывающего белка составляют углеводные остатки, находящиеся в 2 ответвляющихся цепочках и заканчивающихся сиаловой кислотой. Углеводы, вероятно, не участвуют в механизме захвата железа. Синтезируется трансферрин преимущественно в паренхиматозных клетках печени. Функции трансферрина в организме представляют значительный интерес. Он не только переносит железо в различные ткани и органы, но и "узнает" синтезирующие гемоглобин ретикулоциты и, возможно другие нуждающиеся в железе клетки. Трансферрин отдает железо им только в том случае, если клетки имеют специфические рецепторы, связывающие железо. Таким образом, этот железо-связывающий белок функционирует как транспортное средство для железа, обмен которого в организме человека зависит как от общего поступления железа в плазму крови, так и от его количества, захваченного различными тканями соответственно количеству в них специфических рецепторов для железа. Кроме того трансферрин обладает защитной функцией - предохраняет ткани организма от токсического действия железа.
Анализируя биологическую роль трансферрина в организме, следует упомянуть о результатах экспериментальных исследований, свидетельствующих о способности этого белка регулировать транспорт железа из лабильных его запасов в эпителии клеток желудочно-кишечного тракта в плазму крови. Из плазмы железо захватывается преимущественно костным мозгом для синтеза гемоглобина и эритроцитов, в меньшей степени - клетками ретикулоэндотелиальной системы и откладывается в виде запасного железа, некоторое количество его поступает в неэритропоэтические ткани и используется для образования миоглобина и ферментов тканевого дыхания (цитохромы, каталаза и т.д.). Все эти процессы являются сложными и до конца не изученными.
Однако некоторые этапы наиболее важного процесса передачи железа трансферрином клеткам костного мозга можно представить следующим образом:
) адсорбция трансферрина рецепторными участками на поверхности ретикулоцитов;
) образование прочного соединения между трансферрином и клеткой, возможно проникновение белка в клетку;
) перенос железа от железо-связывающего белка к синтезирующему гемоглобин - аппарату клетки;
) освобождение трансферрина в кровь.
Известно, что количество связывающих трансферрин
пространств максимально в ранних эритроидных
предшественниках и уменьшается по мере созревания этих клеток.
Железо-связывающий белок лактоферрин обнаружен во многих биологических жидкостях: молоке, слезах, желчи, синовиальной жидкости, панкреатическом соке и секрете тонкого кишечника. Кроме того, он находится в специфических вторичных гранулах нейтрофильных лейкоцитов, образуясь в клетках миелоидного ряда со стадии промиелоцита. Подобно трансферрину, лактоферрин способен связывать 2 атома железа специфическими пространствами. Он состоит из одной полипептидной цепочки, молекулярный вес приблизительно равен 80000. В физиологических условиях этот железо-связывающий белок насыщен железом до 20% в ничтожных количествах он содержится в плазме крови, освобождаясь в нее из нейтрофильных лейкоцитов. Несмотря на схожесть лактоферрина и трансферрина, эти железо-связывающие белки отличаются друг от друга по антигенным свойствам, составу аминокислот, белков и углеводов.
В настоящее время известны следующие функции этого белка: бактериостатическая, участие в иммунных процессах и абсорбции железа в желудочно-кишечном тракте. Свободный от железа лактоферрин - аполактоферрин обладает бактериостатическими свойствами, которые теряются при насыщении его железом. Аполактоферрин тормозит in vitro рост бактерий и грибов, и возможно, играет роль во внутриклеточной гибели микроорганизмов. При низкой концентрации лактоферрина в нейтрофильных лейкоцитах может уменьшаться их бактерицидная активность.
Растворимые рецепторы трансферрина (sTfR) - это белки, расположенные на поверхности клеток, которые обеспечивают перенос ионов железа внутрь клетки с её поверхности. Часть из них растворима и находится в крови. Рецептор состоит из двух пептидных цепей, проходящих сквозь мембрану клетки. Молекула транферрина, несущая до двух атомов железа, присоединяется к внешнему, экстрацеллюлярному концу рецептора, после чего поглощается клеткой путем эндоцитоза. В сформированной везикуле происходит изменение рН, железо меняет степень окисления (с Fe3+ на Fe2+) и, в дальнейшем, используется для синтеза гемоглобина или сохраняется в форме депонированного железа. Белковая часть трансферрина, освободившись от железа, вместе с рецептором выходит на поверхность клетки, где апотрансферрин отделяется и весь цикл повторяется. При повышенной потребности в железе цикл рецептора трансферрина ускоряется и все больше рецепторов располагается на поверхности клетки. При этом все чаще внешняя (внеклеточная) часть рецептора подвергается расщеплению экстрацеллюлярными протеазами. В результате воздействия протеаз от рецептора отделяется и попадает в кровь довольно стабильный фрагмент - пептид с молекулярным весом 95 a, называемый растворимым рецептором трансферрина, концентрацию которого можно определить с помощью иммуноферментного метода. Приблизительно 80% рецепторов трансферрина находится на плазматической мембране эритропоэтических клеток. Кроме того, наличие рецептора показано в клетках плаценты, в лимфоцитах и даже в некоторых опухолевых клетках. Плотность рецептора трансферрина на поверхности клеток-предшественников эритроцитов повышается, по мере развития клеток, вплоть до ретикулоцита, однако на поверхности зрелого эритроцита рецепторов трансферрина не обнаружено. Мониторинг уровня sTfR позволяет определить терапевтический успех применения эритропоэтина. Обычно при адекватной стимуляции эритропоэтической системы, sTfR начинает повышаться. При локальном дефиците железа уровень ферритина и sTfR изменяются разнонаправлено: ферритин снижается, sTfR повышается. При повышении ферритина при хронических воспалениях, опухолях, эффективную потребность железа можно оценить только при определении концентрации sTfR.
Железосерные ферменты - это еще один важный класс железосодержащих ферментов, участвующих в переносе электронов в клетках животных, растений и бактерий. Железосерные ферменты не содержат гемогрупп, они характеризуются тем, что в их молекулах присутствует равное число атомов железа и серы, которые находятся в особой лабильной форме, расщепляющейся под действием кислот. К железо - серным ферментам относится, например, ферредоксин хлоропластов, осуществляющий перенос электронов от возбужденного светом хлорофилла на разнообразные акцепторы электронов.
Эритропоэз
Эритропоэз - один из самых мощных процессов образования клеток в организме. За 70 лет человек производит около 3,5 тонн эритроцитов. У взрослого человека эритропоэз протекает в костном мозге. Родоначальниками кроветворения являются стволовые клетки, способные давать все ростки клеток крови.
Эритроидный ряд клеток.
Собственно эритроидный ряд (см. табл. 2.1.1) начинается с проэритробластов - потомков стволовых клеток, у которых появляется чувствительность к эритропоэтину. В проэритробластах начинается синтез гемоглобина, продолжающийся в ряду созревающих клеток. По мере накопления гемоглобина в последующих клетках деление приостанавливается, ядро уменьшается в размере. На последнем этапе ядро из клетки удаляется, затем исчезают остатки РНК, еще выявляемые при специальной окраске в молодых эритроцитах - ретикулоцитах. Наиболее известным фактором, регулирующим эритропоэз, является гормон эритропоэтин (ЭПО), синтезируемый в почках. Уровень ЭПО повышается при быстром развитии гипоксии - обильная кровопотеря, острый гемолиз, острая ишемия почек, подъем в горы. При хронических анемиях уровень ЭПО, как правило, нормален, исключая апластическую анемию, сопровождающуюся чрезвычайно высоким уровнем гормона.
табл. 2.1.1
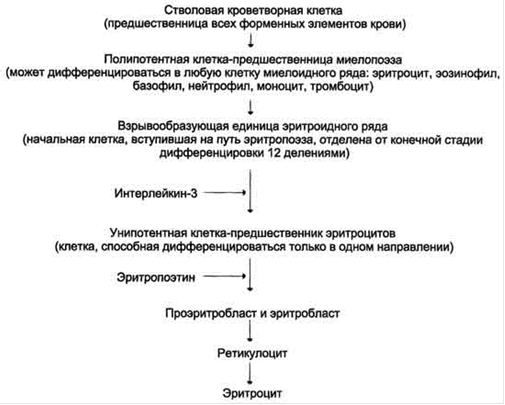
Регуляция эритропоэза
Наряду с ЭПО, в крови присутствуют ингибиторы эритропоэза (ИЭПО). Это разнообразные вещества, часть которых относится к среднемолекулярным токсинам, образующимся в нормальных и патологических процессах. Баланс активности ЭПО и ИЭПО регулирует эритропоэз. При необходимости экстренной регенерации индуцируется эритропоэтиновый механизм - значительное преобладание активности ЭПО над активностью ИЭПО. При почечной недостаточности и ряде хронических заболеваний, сопровождающихся повышенным образованием или нарушением выведения среднемолекулярных токсинов, активность ИЭПО доминирует над ЭПО, что ведет к угнетению эритропоэза и развитию анемии хронических заболеваний.
Синтез гемоглобина
Гемоглобин составляет 95% белка эритроцитов. Молекула гемоглобина состоит из двух пар различных по структуре цепей глобинового белка, в каждой из которых находится железосодержащая протопорфириновая группа - гем. Синтез гемоглобина начинается с ранних этапов эритропоэза и идет путем синхронной продукции гема и глобиновых цепей с образованием законченной молекулы.
Образование гемоглобина регулируется ЭПО, рост концентрации которого ускоряет синтез глобина. Активность поступления железа в клетки костного мозга также регулирует образование гемоглобина. Уровень железа в клетках определяет скорость образования гема. В свою очередь, свободный гем через белки-регуляторы усиливает синтеза глобина. Эта связь систем синтеза гема и глобина синхронизирует их работу - в норме лишь незначительное количество свободного протопорфирина и глобина остается не связанным в гемоглобине.
Регенерация красной крови
Действие многих факторов ведет к нарушению мембран и внутренней структуры эритроцитов. Физиологический гемолиз измененных эритроцитов - постоянный процесс в организме. Основная масса эритроцитов разрушается в селезенке. Здесь имеется открытый кровоток, и эритроциты плотно контактируют с макрофагами в паренхиме органа. У здорового человека разрушается 180-270 млн. эритроцитов в сутки. Железо этих эритроцитов накапливается в макрофагах и обменивается через трансферрин крови с костным мозгом и другими органами и тканями. В норме скорость регенерации красной крови соответствует уровню физиологического гемолиза. При массивной кровопотере или повышенном кроверазрушении активность эритропоэза за счет выброса ЭПО может возрастать в сотни раз. Адекватный уровень эритропоэза невозможен при недостатке или нарушениях обмена ряда незаменимых, поступающих только с пищей веществ. Витамин В12 и фолиевая кислота, участвуя в обмене нуклеотидов, являются незаменимыми факторами для синтеза ДНК и деления любых клеток. Недостаток в организме этих витаминов приводит к развитию мегалобластных анемий: В12-дефицитной и фолиеводефицитной анемии. Недостаточное поступление железа в клетки костного мозга ограничивает возможность синтеза гема и, как следствие, - гемоглобина. Угнетение синтеза гема наблюдается и при недостатке витамина В6, в связи с тем, что этот процесс осуществляется ферментами, зависимыми от пиридоксина. Недостаток в организме железа и витамина В6 сопровождается развитием микроцитарных анемий: сидеропенической (ЖДА) и сидеробластной, соответственно.
Метаболизм железа
Обмен железа
В организме здорового человека содержится около 3-4 г железа (40-50 мг Fe/кг). Выделяют три фонда железа: - функциональное железо находится в составе гемоглобина, миоглобина и железосодержащих энзимов. - транспортная форма представлена железом в составе трансферрина. - депонированное железо находится в составе ферритина и гемосидерина внутренних органов. Обмен железа является высокоорганизованным процессом, в котором все железо, выделяемое при распаде гемоглобина и других железосодержащих белков, вновь утилизируется. Поэтому, несмотря на то, что ежедневно абсорбируется и выводится лишь малая часть железа (0,03-0,05%), его метаболизм очень динамичный. Кинетика железа здорового мужчины весом 70 кг имеет следующие параметры: эритроциты - 2000-2500 мг; плазма - 4 мг; запасы - 800-1000 мг; миоглобин и ферменты - 300 мг; всасывание - 1-2 мг/сутки; потери - 1-2 мг/сутки; высвобождение при гемолизе - 25 мг/сутки; потребление при эритропоэзе - 25 мг/сутки.
Содержание железа в сыворотке в норме приведено ниже (Табл.2.4.1).
Табл. 2.4.1
| Возраст
| Содержание железа
|
|
| Мкг/дл
| Ммоль/л
|
| Новорожденные
| 100-200
| 17,9-44,75
|
| Дети до 2 лет
| 40-100
| 7,16-17,9
|
| Дети старше 2 лет
| 50-120
| 8,95-21,48
|
| Взрослые:
|
|
|
| Мужчины женщины
| 50-160 40-150
| 8,95-28,64 7,16-26,85
|
|
|
|
|
| | | | | |
Потери железа
Потери железа из организма в норме происходят, главным образом, через кишечник: часть выделяется вместе с желчью, часть - с отторгающимся эпителием. Кроме того, железо теряется с эпителием кожи, волосами, с мочой, а у женщин репродуктивного возраста - с менструальными кровопотерями.
Железодефицитные состояния
Определение
Железодефицитное состояние или недостаточность железа определяется как дефицит общего железа, обусловленный несоответствием между его поступлением и расходом (потери или возросшее потребление), приводящим к отрицательному балансу.
Железодефицитная анемия (ЖДА) - синдром, характеризующийся снижением наполнения гемоглобина железом с последующим уменьшением содержания гемоглобина в эритроците и угнетением эритропоэза из-за дефицита железа.
Нарушение усвоения железа
Любые патологические состояния двенадцатиперстной и проксимальных отделов тонкой кишки могут приводить к развитию дефицита железа. Значительное нарушение всасывания железа может быть последствием гастроэктомии с выключением части двенадцатиперстной кишки, резекции тонкой кишки или хронического панкреатита и обусловленного им синдрома мальабсорбции. У некоторых больных, нарушенная абсорбция железа может маскироваться общими синдромами, такими как стеаторрея, спру, целиакия или диффузный энтерит. Атрофический гастрит и сопутствующая ахлоргидрия также снижают всасывание железа. Недостаточность железа часто возникает после операций на желудке и гастроэнтеростомии.
Нарушение всасывания железа может быть обусловлено избыточным содержанием в продуктах питания ингибиторов абсорбции железа (фитины, танины, фосфаты, оксалаты, кальций). Специфической причиной низкого усвоения железа может являться дефицит меди в организме меди, который нередко развиваться при длительном, несбалансированном применении препаратов цинка, блокирующих усвоение меди. Угнетение абсорбции железа может развиваться и вследствие длительного применение при язвенной болезни и гастродуоденитах антацидов и препаратов с обволакивающим механизмом действия. Как самостоятельный фактор, нарушение всасывания железа является редкой причиной развития дефицита железа, но он приобретает большое значение при сочетании с другими факторами, нарушающими метаболизм железа.
Кровопотери
Острые потери крови, снижая количество эритроцитов, ведут к развитию острой постгеморрагической анемии. В дальнейшем, в зависимости от степени кровопотери, компенсаторной реакции эритропоэза и наличия резервов железа, может развиваться картина железодефицитного состояния. Но, тем не менее, ведущее место среди причин дефицита железа занимают хронические кровопотери.
Маточные кровопотери являются основной причиной дефицита железа у женщин репродуктивного возраста. Они могут наблюдаться при меноррагиях различной этиологии, яичниковой дисфункции, аденоматозе, миоме матки, эндометриозе, злокачественных опухолях матки, наличии внутриматочных контрацептивов. Кровопотери из кишечной трубки являются наиболее частой причиной дефицита железа у мужчин и неменструирующих женщин. Потери крови могут возникать на всех уровнях пищеварительной системы. При воспалительных заболеваниях десен, стоматитах, тромбоцитопатиях, ДВС-синдроме, ангиоматозах могут наблюдаются хронические десневые кровотечения. Эрозивный эзофагит (чаще - при недостатке кардии или грыже пищеводного отверстия диафрагмы), варикоз вен пищевода и кардиального отдела желудка (при циррозе печени и других формах портальной гипертензии) могут стать причинами хронических пищеводных кровопотерь. Эрозии желудка, язвы желудка и двенадцатиперстной кишки, опухоли и полипы желудка могут стать причиной хронических кровопотерь, связанных с патологией желудка. Значение хронических кровопотерь из кишечника как причины дефицита железа увеличивается с возрастом из-за роста частоты дивертикулеза тонкого кишечника, дивертикулярной болезни, опухолей тонкого кишечника, чаще - полипоза и опухолей толстого кишечника. Реже потери крови из кишечника обусловлены болезнью Крона и неспецифическим язвенным колитом.
Кровоточащий геморрой является также одной из самых распространенных причин дефицита железа. Носовые кровотечения, как и десневые, являются причиной развития дефицита железа, главным образом, у больных с геморрагическими диатезами (телеангиоэктазия, тромбоцитопатия, тромбоцитопенические пурпуры, ДВС - синдром) или при ангиопатиях-ангиоматозах. Относительно редкой причиной дефицита железа являются кровопотери в замкнутые полости и ткани, которые могут наблюдаться при эндометриозе, изолированном легочном сидерозе, ангиопатиях, гломических опухолях. Причиной недостаточности железа также может являться гематурия, наблюдаемая при хроническом гематурическом нефрите, мочекаменной болезни, поликистозе почек, геморрагическом цистите, а также перманентном внутрисосудистом гемолизе (болезнь Маркиафавы), пароксизмальной ночной гемоглобинурии, механическом внутрисосудистом гемолизе (маршевая анемия).
Значительную и часто неоцениваемую адекватно роль играют ятрогенные кровопотери. К развитию дефицита железа могут приводить частые, повторные заборы крови для исследований.
Нарушения транспорта железа
Картина железодефицитного состояния без истинного дефицита железа может развиваться при снижении в крови уровня трансферрина. Это может иметь место при гипопротеинемиях различной этиологии (нефротический синдром, нарушение белок-синтетической функции печени, синдром мальабсорбции, алиментарная белковая недостаточность). В редких случаях концентрация трансферрина или нарушения его строения могут быть генетически детерминированы.
Сидеропения (гипосидероз)
Гипосидероз связан с тканевым дефицитом железа, необходимого для нормальной трофики органов и тканей. Уже на ранних стадиях развития дефицита железа снижаются его запасы в тканях и наблюдаются изменения со стороны максимально чувствительных к дефициту тканевого железа систем. К трофическим нарушениям вследствие гипосидероза относится и вторичный иммунодифецит, проявляющийся склонностью к хронизации воспалительных процессов и частыми инфекционно-воспалительными заболеваниями. Симптоматика гипосидероза зависит от глубины тканевого дефицита железа. При значительном дефиците железа, сопровождающем анемию, ведущими проявлениями являются изменения со стороны кожи и ее придатков, общая слабость, невозможность сосредоточиться, снижение памяти, утомляемость, длительная астенизация после перенесенных заболеваний, появление специфической бледности кожи, часто с алебастровым или зеленоватым оттенком.
Гипосидероз без анемического синдрома имеет большое самостоятельное значение, так как имеются доказательства того, что клинически не манифестированный дефицит железа играет роль в развитии умственной отсталости детей, сердечной недостаточности, болезни Паркинсона и Альцгеймера.
Анемический синдром.
Анемический синдром при железодефицитных состояниях проявляется хорошо известными и неспецифическими для анемии любого происхождения симптомами, выраженность которых во многом зависит от скорости снижения уровня гемоглобина. Как правило, ЖДА развивается постепенно, и больные адаптируются к ней, часто приписывая свои недомогания переутомлению, психическим и физическим перегрузкам. В связи с этим в клинической картине ЖДА доминируют симптомы гемосидероза, а не малокровия.
Нормальные значения
| Возраст
| Женщины
| Мужчины
|
| 0-2
| 24,0 - 31,0пг
| 24,5 - 29,0пг
|
| 3-6
| 25,0 - 30,5пг
| 25,5 - 29,5пг
|
| 7-12
| 25,5 - 31,0пг
| 26,0 - 31,0пг
|
| 13-16
| 27,0 - 32,0пг
| 26,5 - 32,0пг
|
| 17-19
| 27,0 - 33,0пг
| 27,0 - 32,5пг
|
| 20-29
| 27,5 - 33,0пг
| 27,5 - 33,0пг
|
| 30-39
| 27,0 - 34,0пг
| 27,5 - 33,5пг
|
| 40-49
| 27,0 - 34,0пг
| 27,5 - 34,0пг
|
| 50-59
| 27,0 - 34,5пг
| 27,5 - 34,0пг
|
| 60-65
| 26,5 - 33,5пг
| 27,0 - 34,5пг
|
| >65
| 26,0 - 34,0пг
| 26,0 - 35,0пг
|
Материал для исследования: венозная или капиллярная кровь. Кровь берется с ЭДТА.
МСН (mean corpuscular hemoglobin) характери






