Лекция 6.
Биофизика клеточных процессов.
Искусственные липидные мембран.
Липосомы. При набухании сухих фосфолипидов в воде или при впрыскивании их раствора в воду происходит происходит самосборка молекул и образование фосфолипидных везикул – липосом. При этом неполярные хвосты молекул находятся внутри би- и или полимолекулярных слоев и не соприкасаются с водой. Обычно получаются многослойные липосомы, т.е. состоящие из нескольких бимолекулярных слоев. В таких структурах, отдельные бимолекулярные слои отделены друг от друга слоями воды. Толщина билипидных слоев в многослйных липосомах составляет 6,5 – 7,5 нм, водных слоев - 1,5 – 2 нм. Диаметр многослойных липосом может достигать до 400 -500 нм. Для получения однослойных липосом суспензию многослойных липосом обрабатывают ультразвуком. Диаметр таких структур составляет 25-30 нм. Современные методы позволяют получит однослойные липосомы большего диаметра, до 500 нм. Такие структуры можно рассматривать как прототипы клеток, а билипидный слой – как модель мембраны для исследований различных свойств клеточных мембран. Другой тип искусственных мембран, используемых в качестве модельных систем – плоские бислойные липидные мембраны (БЛМ). Такие мембраны получают на маленьких отверстиях диаметром 1 мм, проделанных на фторопластовых пластинах.

Рис.4. Схема строения однослойной липосомы (Антонов, 28 с)
На отверстие пластины, погруженного в воду, наносят каплю раствора липидов, растворенных в неполярном растворителе (спирты, хлороформ, ацетон). Растворитель диффундирует в воду и на отверстии образуется липидная пленка. Она спонтанно утончается до бимолекулярного слоя. БЛМ (наряду с липосомами) широко используются в качестве модельных систем для изучения проницаемости, электрических свойств мембран, мембранного транспорта и других свойств и функций мембран. современные методики позволяют вводить в состав БЛМ молекулы белков и конструировать модели биологических мембран, выполняющих разнообразные функции.
Пассивный транспорт
Пассивный транспорт – это перенос вещества в сторону уменьшения химического и электрохимического потенциалов. Как отмечали выше, пассивный транспорт веществ может осуществляться в процессе простой или облегченной диффузии. Движущей силой пассивного транспорта являются градиенты концентрации и электрического потенциала. В случае транспорта неэлектролитов (Z = 0) или в отсутствие электрического поля dφ/dx = 0 плотность потока вещества (количество вещества в единицу времени через единицу площади поверхности, перпендикулярной направлению переноса) описывается уравнением dm/dt = -URTS dc/dx, где U - подвижность частиц, с - концентрация частиц, S – площадь переноса.. Согласно соотношению Эйнштейна, коэффициент диффузии D = URT, тогда
dm/dt = -DS dc/dx (1 закон Фика).
Простая диффузия веществ может осуществляться через липидный бислой, через поры в билипидном слое и через белковые поры. Скорость простой диффузии определяется градиентом концентрации вещества в мембране (разностью концентрации снаружи и внутри клетки) и коэффициентом проницаемости мембраны (Р). Коэффициент проницаемости зависит от свойств мембраны и переносимых веществ и определяется как P = DK/l, где D - коэффициент диффузии, К – коэффициент распределения, l -толщина мембраны. Как видно из этой формулы, коэффициент проницаемости тем больше, чем больше коэффициент диффузии, чем тоньше мембрана, и чем лучше вещество растворяется в липидном бислое.
В липидной фазе мембран хорошо растворяются неполярные соединения, например, органические жирные кислоты, эфиры. Эти вещества хорошо проникают через липидный бислой.
Вода и полярные водорастворимые соединения (соли, основания, сахара, аминокислоты, спирты) плохо растворяются в липидной фазе и поэтому не должны проходить через билипидный слой мембран. Однако, известно, что все биологические мембраны очень хорошо проницаемы для молекул воды и малых гидрофильных молекул. Этот факт свидетельствует о том, что существуют специальные пути транспорта таких молекул через мембраны. В настоящее время известно три способа транспорта воды и водорастворимых молекул через мембраны: образование кинков, через липидные поры, при помощи переносчиков (облегченная диффузия).
Образование кинков.
Проникновение гидрофильных молекул и воды через липидный бислой связывают с образованием между жирно-кислотными хвостами липидных молекул свободных полостей вследствие их теплового движения, так называемых кинков (от англ. kink – петля). Образование кинков происходит за счет гош-транс-гош –конфигураций липидных молекул (рис.?).
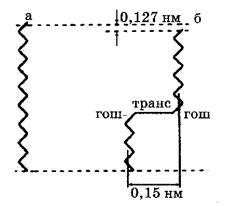
Рис. 5. Схематичное изображение образования кинка липидной фазе биологических мембран Антонов, 37 стр)
а. Углеводордная цепь липида в транс-конформации;
б. Углеводордная цепь липида в гош- транс- гош -конформации
Вследствие теплового движения молекул, кинки могут перемещаться поперек мембраны и переносить попавшие в них небольшие молекулы, в первую очередь, молекулы воды.
Второй путь проникновения нерастворимых в липидах молекул – через липидные и белковые поры в мембранах.
Липидные поры
Барьерные и механические свойства клеточных и внутриклеточных мембран обуславливаются непрерывностью липидного бислоя. Однако, в процессе жизнедеятельности клетки, непрерывность бислоя может нарушаться и приводить к образованию структурных дефектов типа сквозных гидрофильных пор. Естественно, при этом изменяется и параметры биомембран, в частности проницаемость, стабильность. Сквозные поры в липидном бислое появляются в результате действия различных факторов: тепловых флуктуаций поверхности бислоя, энергетического пробоя, замораживания, действия ПАВ, осмотического давления, окисления молекул липидов и т.д. Один из наиболее типичных и хорошо изученных примеров дестабилизации мембран – гемолиз эритроцитов. Это явление начинается с набухания клеток в результате осмотического давления, при помещении их в гипотонический раствор. При набухании мембрана эритроцита растягивается, и при определенном пороговом уровне натяжения мембраны появляются гидрофильные липидные поры. (Рис.3 Антонов,с. 49). Через эти поры гемоглобин и низкомолекулярные соединения выходят из клетки, что приводит к снижению разности осмотического давления на мембране. Натяжение мембраны снижается, и поры затягиваются. Белки цитоскелета позволяют эритроциту сохранить форму и при этом образуется так называемая «тень эритроцита». В отсутствие цитоскелета и его недостаточного развития прочность клетки определяется прочностью мембраны, т.е. наличием и размерами липидных пор. Если размеры поры меньше критического размера, она затягивается и зарастает. Неограниченный рост поры приводит к разрушению мембраны. По своему происхождению, структуре и функциям липидные поры принципиально отличаются от белковых каналов. Белковые каналы характеризуются определенными размерами, которые не изменяются в течение всей жизни клетки. Размеры липидных пор не постоянны, они варьируют в широких пределах в процессе образования и зарастания (табл. 3). Если размер поры меньше критического, то пора, в процессе зарастания, проходит все промежуточные радиусы и достигает минимального размера.
Таблица 3
Размеры липидных пор в модельных и клеточных мембранах
| Тип мембраны
| Радиус поры, нм
| Тип воздействия на мембрану
|
| Эритроцит
Эритроцит
L- клетка
Липосомы
Липосомы
Билипидная мембрана
| 3,0 – 4,0
2,0
1,2
0,2- 2,0
0,6 – 0,8
1,2-1,8
| Электрический пробой
Осмотический гемолиз
Электрический пробой
Осмотический гемолиз
Фазовый переход
Фазовый переход
|
Предполагается, что липидные поры полностью не затягиваются, так как этому препятствуют мощные силы гидратации, проявляющиеся при сближении стенок гидрофильных пор. Липидные поры, в отличие от белковых каналов, не обладают избирательной проницаемостью по отношению к тем или иным молекулам и ионам. Однако, по мере затягивания размеры липидных могут стать соизмеримыми с размерами ионных каналов. Экспериментально показано, что через определенное время после снятия стрессового воздействия, проводимость мембраны возвращается в исходное состояние. Это объясняется тем, что образованные при стрессе липидные поры затягиваются до маленьких размеров и не способны пропускать гидратированные ионы. На последних этапах затекания липидные поры пропускают только молекулы и ионы воды. Как видно, через гидрофильные липидные поры могут проходить и высокомолекулярные вещества, и низкомолекулярные соединения, органические и неорганические ионы, молекулы воды. Показано, что большие поры могут пропускать в клетку такие гигантские молекулы как молекулы ДНК. Механизм этого явления пока непонятен. Средний диаметр статистического клубка ДНК достигает до 2000 нм. Критической размер липидной поры около 9 нм. Предполагается, что липидная пора в этом случае служит якорем для фиксации свободных концов ДНК, что удерживает молекулу на определенном участке мембраны. Дальнейший перенос молекулы ДНК в клетку происходит по механизму пиноцитоза.
Облегченная диффузия
В ряде случаев, посредниками для переноса органических молекул служат специальные молекулы. Эти молекул по химической природе являются белками, полипептидами или олигопептидами. Эти молекулы способны узнавать определенные вещества и транспортировать их через мембрану. Этот вид транспорта называют облегченной диффузией. Широко известна облегченная диффузия ионов калия при помощи антибиотика валиномицина. Молекула валиномицина состоит из 6 остатков аминкислот и 6 остатков кетокислот, имеет цилиндрическую форму. Внутри цилиндра располагаются полярные группировки, снаружи – неполярные. Поэтому эта молекула хорошо растворяется в липидной фазе и может свободно диффундировать через липидный бислой мембран. Каждая молекула валиномицина способна связывать один ион калия. При взаимодействии валиномицина с К+ происходит разрушение гидратной оболочки иона и образование комплекса. Комплекс валиномицин – К+ по градиенту концентрации диффундирует через мембрану. При низкой концентрации ионов калия в среде комплекс разрушается и свободный валиномицин диффундирует обратно. Облегченная диффузия имеет свои характерные особенности. Как видно, перенос ионов калия валиномицином может происходить в ту или иную сторону в сторону меньшей концентрации ионов. Облегченная диффузия имеет ряд характерных отличий от простой диффузии.
При облегченной диффузии скорость переноса значительно выше.
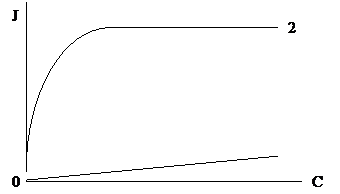
Процесс облегченной диффузии характеризуется свойством насыщения. При повышении концентрации переносимого вещества скорость переноса возрастает лишь до определенного предела, когда все молекулы переносчика связаны с переносимым веществом. Затем скорость транспорта не повышается (Рис. 6).
Рис. 6. Зависимость скорости переноса в клетку через мембрану от концентрации вещества во внеклеточном пространстве
1 – простая диффузия; 2 – облегченная диффузия
Облегченная диффузия характеризуется высокой специфичностью и избирательностью. Так, показано, что через плазматическую мембрану таким путем транспортируется только L – аминокислоты, но аминокислоту. Переносчики сахаров, наоборот, транспортируют через мембрану, только D-изомеры. Если переносчик способен транспортировать разные типы молекул, то наблюдается конкуренция между ними. При этом одни типы молекул переносятся лучше, чем другие. Например, через мембрану эритроцитов, глюкоза переносится лучше, чем фруктоза, фруктоза лучше, чем ксилоза, ксилоза лучше, чем арабиноза и.т.д.
Работа переносчика может блокироваться веществом, имеющим сходную структуру с транспортируемым соединением. Например, флоридзин подавляет транспорт сахаров через мембрану.
Как видно, работа белковых переносчиков на мембранах имеют много общего с работой ферментов.
Свойства ионных каналов.
Ионных каналы биологических мембран обладают определенными свойствами, которые характеризуют их работу.
Селективность (избирательность)– способность канала избирательно пропускать ионы одного типа. Эксперименты показали, что ионные каналы обладают абсолютной селективностью по отношению ионам с различными зарядами, т.е. существуют катион-селективные каналы и анион-селективные каналы. Любой ионный канал способен пропускать ионы одного знака, но с различной скоростью. Так, через натриевый канал на мембране могут проходить и ионы калия, но коэффициент проницаемости для них в 20 раз ниже, чем для ионов натрия. В то же время, натриевый канал абсолютно непроницаем для анионов, например, для ионов хлора. Способность ионного канала пропускать с неодинаковой скоростью различные ионы одного знака называется относительной селективностью канала.
Независимость работы отдельных каналов. Прохождение ионов через канал не зависит от того, идет ли ток ионов через другие каналы на этой мембране. Например, включение или выключение калиевого канала не влияет на работу натриевого канала, и наоборот. Влияние канала на функционирование других каналов может происходит опосредованно, за изменения значения электрического потенциала на мембране.
Дискретный характер проводимости. Как уже отмечалось, ионный канал может находиться в двух состояниях: открытом или закрытом. Переходы между этими состояниями происходят в случайные моменты времени и подчиняются статистическим закономерностям. Нельзя утверждать, что данный ионный канал откроется точно в какой-то момент времени, можно говорить лишь о вероятности открывания канала в определенный интервал времени. Интервал времени, в течение которого канал находится в открытом состоянии, называют временем открытого состояния канала. Среднее значение этой величины для Na+ - каналов составляет около 0,7 мс (0,3 – 1,5 мс для Na+ -каналов различных мембран). Промежуток времени, в течение которого вероятность открывания отдельного канала велика, называют временем жизни канала. Для натриевых каналов она составляет около 2 мс.
Управляемость канала. Ионные каналы чувствительны к изменению мембранного потенциала. Чувствительный к изменению электрического поля участок молекулы белка называют сенсором напряжения. Изменение напряжения на мембране приводит к конформационной перестройке белковой молекулы, что меняет вероятность открывания или закрывания ворот канала. Экспериментально показано, что деполяризация мембраны нервных клеток приводит к открыванию натриевых каналов. Ионные каналы могут быть чувствительны и к другим воздействиям: механическим, химическим, термическим и т.д. Такие свойства характерны для ионных каналов мембран рецепторных клеток, соответственно, механорецепторов, хеморецепторов, терморецепторов.
Вопросы и задания для самоконтроля.
- Назовите наиболее распространенных представителей липидов, входящих в состав биологических мембраны.
- Нарисуйте схематично структуру биомембраны. Объясните характерное расположение молекул липидов и белков в мембранах.
- Какова роль воды в формировании и стабилизации мембранных структур?
- Остатки каких молекул выступают в качестве полярной части молекулы липида?
- Чем отличаются ионные каналы от мембранных пор?.
- Опишите процесс получения липосом, искусственных мембран.
- Как определить температуру плавления определенной биологической мембраны?
- Объясните выражение «скорость латеральной диффузии фосфатидилсерина составляет 3 мкм/с».
- Опишите, каким путем транспортируются в мембрану молекулы: глюкозы, серина, аденина, валиномицина, воды.
- Какие ионы будут проходить через натриевый канал на плазматической мембране?
Лекция 6.
Биофизика клеточных процессов.