

Кормораздатчик мобильный электрифицированный: схема и процесс работы устройства...

История создания датчика движения: Первый прибор для обнаружения движения был изобретен немецким физиком Генрихом Герцем...
Кормораздатчик мобильный электрифицированный: схема и процесс работы устройства...
История создания датчика движения: Первый прибор для обнаружения движения был изобретен немецким физиком Генрихом Герцем...
Топ:
Комплексной системы оценки состояния охраны труда на производственном объекте (КСОТ-П): Цели и задачи Комплексной системы оценки состояния охраны труда и определению факторов рисков по охране труда...
Теоретическая значимость работы: Описание теоретической значимости (ценности) результатов исследования должно присутствовать во введении...
Когда производится ограждение поезда, остановившегося на перегоне: Во всех случаях немедленно должно быть ограждено место препятствия для движения поездов на смежном пути двухпутного...
Интересное:
Лечение прогрессирующих форм рака: Одним из наиболее важных достижений экспериментальной химиотерапии опухолей, начатой в 60-х и реализованной в 70-х годах, является...
Инженерная защита территорий, зданий и сооружений от опасных геологических процессов: Изучение оползневых явлений, оценка устойчивости склонов и проектирование противооползневых сооружений — актуальнейшие задачи, стоящие перед отечественными...
Финансовый рынок и его значение в управлении денежными потоками на современном этапе: любому предприятию для расширения производства и увеличения прибыли нужны...
Дисциплины:
|
из
5.00
|
Заказать работу |
Осуществляется интракардиальными периферическими рефлексами. Интракардиальные рефлекторные дуги включают афферентные интрамуральные нейроны Догеля II типа, дендриты которых образуют рецепторы растяжения миокарда и коронарных сосудов, а также эфферентные нейроны, аксоны которых иннервируют миокард и гладкую мускулатуру коронарных сосудов.
Скопления тел эфферентных нейронов представляют собой интрамуральные сердечные ганглии, их составляют не только холинергические, но и адренергические клетки, причем последние обладают большей возбудимостью по сравнению с холинергическими. Эффекты раздражения эфферентных интрамуральных нейронов могут быть противоположными в зависимости от степени кровенаполнения сердца. При слабом кровенаполнении афферентация от рецепторов сердца ведет к возбуждению адренергических нейронов, а выделяемый ими медиатop норадреналин оказывает стимулирующее влияние на сердце. При чрезмерном наполнении камер сердца кровью и интенсивном раздражении рецепторов возбуждаются холинергические эфферентные нейроны, оказывая тормозные эффекты на сердце.
Внутрисердечные рефлекторные дуги — часть метасимпатической нервной системы. Эфферентные нейроны являются также общими с дугой классического парасимпатического рефлекса.
В эксперименте показано, что увеличение растяжения миокарда правого предсердия (в естественных условиях оно возникает при увеличении притока крови к сердцу) приводит к усилению сокращений левого желудочка. Таким образом, усиливаются сокращения не только того отдела сердца, миокард которого непосредственно растягивается притекающей кровью, но и других отделов, чтобы «освободить место» притекающей крови и ускорить выброс ее в артериальную систему. Доказано, что эти реакции осуществляются с помощью внутрисердечных периферических рефлексов.
+Подобные реакции наблюдаются лишь на фоне низкого исходного кровенаполнения сердца и при незначительной величине давления крови в устье аорты и коронарных сосудах. Если камеры сердца переполнены кровью и давление в устье аорты и коронарных сосудах высокое, то растяжение венозных приемников в сердце угнетает сократительную активность миокарда. При этом сердце выбрасывает в аорту в момент систолы меньшее, чем в норме, количество содержащейся в желудочках крови. Задержка даже небольшого дополнительного объема крови в камерах сердца повышает диастолическое давление в его полостях, что вызывает снижение притока венозной крови к сердцу. Излишний объем крови, который при внезапном выбросе его в артерии мог бы вызвать пагубные последствия, задерживается в венозной системе. Подобные реакции играют важную роль в регуляции кровообращения, обеспечивая стабильность кровенаполнения артериальной системы.
Опасность для организма представляло бы и уменьшение сердечного выброса — оно могло бы вызвать критическое падение артериального давления. Такую опасность также предупреждают регуляторные реакции внутрисердечной системы.
Недостаточное наполнение кровью камер сердца и коронарного русла вызывает усиление сокращений миокарда посредством внутрисердечных рефлексов. При этом в момент систолы в аорту выбрасывается большее, чем в норме, количество содержащейся в них крови. Это и предотвращает опасность недостаточного наполнения кровью артериальной системы. К моменту расслабления желудочки содержат меньшее, чем в норме, количество крови, что способствует усилению притока венозной крови к сердцу.
В естественных условиях внутрисердечная нервная система не является автономной. Опалишь низшее звено в сложной иерархии нервных механизмов, регулирующих деятельность сердца. Более высоким звеном в иерархии являются сигналы, поступающие по симпатическим и блуждающим нервам, экстракардиальной нервной системе регуляции сердца.


55. Внутриклеточные механизмы интракардиальной регуляции сердца: гетерометрическая и гомеометрическая регуляция сердца.
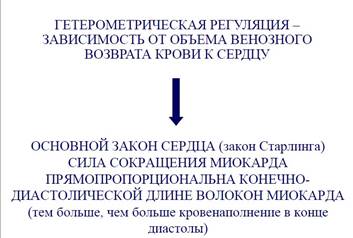
- гетерометрическая регуляция (закон Старлинга);
Эффект Старлинга (Франка-Старлинга)
• Сущность: чем больше растяжение сердечной мышцы перед сокращением, тем сильнее сокращение.
• Механизм: в нерастянутом состоянии центральные концы актиновых нитей частично перекрываются друг с другом, что уменьшает площадь их контакта с миозиновыми нитями. Чем больше растянута мышца, тем меньше перекрываются друг с другом актиновые нити, следовательно, тем больше площадь их контакта с миозиновыми нитями и больше образуется актомиози- новых мостиков; соответственно, больше и общая энергия сокращения
Значение механизма Франка— Старлинга заключается в усилении сердечной деятельности в случае увеличения притока крови к сердцу (преднагрузкй). Чем больше крови скапливается в желудочках во время диастолы, тем сильнее растягиваются волокна и энергичнее сокращаются в систолу. Благодаря этому
1) Устанавливается соответствие между притоком крови к сердцу и оттоком от него.
2) Осуществляется синхронное сокращение правой и левой половины сердца.
- гомеометрическая регуляция (феномен Анрепа).

 Эффект Анрепа — это увеличение силы сокращения и сократимости при увеличенном давлении в аорте и лёгочном стволе (постнагрузки), при неизменной частоте сокращения и неизменном ударном объёме. Это изменение силы сокращений при неменяющейся исходной длине волокон миокарда. Это прежде всего ритмозависимые изменения силы сокращений. Если стимулировать полоску миокарда при равном растяжении со все увеличивающейся частотой, то можно наблюдать увеличение силы каждого последующего сокращения («лестница» Боудича)
Эффект Анрепа — это увеличение силы сокращения и сократимости при увеличенном давлении в аорте и лёгочном стволе (постнагрузки), при неизменной частоте сокращения и неизменном ударном объёме. Это изменение силы сокращений при неменяющейся исходной длине волокон миокарда. Это прежде всего ритмозависимые изменения силы сокращений. Если стимулировать полоску миокарда при равном растяжении со все увеличивающейся частотой, то можно наблюдать увеличение силы каждого последующего сокращения («лестница» Боудича)
56. Экстракардиальная нервная регуляция нагнетательной функции сердца. Хронотропный, инотропный, батмотропный, дромотропный регуляторные эффекты.
Работа сердца регулируется двумя нервами: блуждающим (или вагусом), относящимся к парасимпатической нервной системе, и симпатическим. Эти нервы образованы двумя нейронами. Тела первых нейронов, отростки которых составляют блуждающий нерв, расположены в продолговатом мозге. Отростки этих нейронов заканчиваются в инграмуральных ганглиях сердца. Здесь находятся вторые нейроны, отростки которых идут к проводящей системе, миокарду и коронарным сосудам.
Первые нейроны симпатической нервной системы, регулирующей работу сердца, лежат в боковых рогах I-V грудных сегментов спинного мозга. Отростки этих нейронов заканчиваются в шейных и верхних грудных симпатических узлах. В этих узлах находятся вторые нейроны, отростки которых идут к сердцу. Большая часть симпатических нервных волокон направляются к сердцу от звездчатого ганглия. Нервы, идущие от правого симпатического ствола, в основном подходят к синусному узлу и к мышцам предсердий, а нервы левой стороны — к атриовентрикулярному узлу и мышцам желудочков
Характеристика хронотропного, инотропного, батмотропного, дромотропного и клинотропного регуляторных эффектов на сердце.
· хронотропный (изменяют частоту сердечных сокращений);
· инотропный (изменяют силу сердечных сокращений);
· батмотропный (влияют на возбудимость миокарда);
· дромотропный (влияет на проводимость);
· тонотропный (влияют на тонус миокарда);
Сердце — обильно иннервированный орган. Среди чувствительных образований сердца основное значение имеют две популяции механорецепторов, сосредоточенных, главным образом, в предсердиях и левом желудочке: А-рецепторы реагируют на изменение напряжения сердечной стенки, а В-рецепторы возбуждаются при ее пассивном растяжении.
Афферентные волокна, связанные с этими рецепторами, идут в составе блуждающих нервов. Свободные чувствительные нервные окончания, расположенные непосредственно под эндокардом, представляют собой терминали афферентных волокон, проходящих в составе симпатических нервов. Эфферентная иннервация сердца осуществляется при участии обоих отделов вегетативной нервной системы. Тела симпатических преганглионарных нейронов, участвующих в иннервации сердца, располагаются в сером веществе боковых рогов трех верхних грудных сегментов спинного мозга. Преганглионарные волокна направляются к нейронам верхнего грудного (звездчатого) симпатического ганглия.
Постганглионар-ные волокна этих нейронов вместе с парасимпатическими волокнами блуждающего нерва образуют верхний, средний и нижний сердечные нервы, Симпатические волокна пронизывают весь орган и иннервируют не только миокард, но и элементы проводящей системы. Тела парасимпатических преганглионарных нейронов, участвующих в иннервации сердца, располагаются впродолговатом мозге. Их аксоны идут в составе блуждающих нервов. После вхождения блуждающего нерва в грудную полость от него отходят веточки, которые включаются в состав сердечных нервов. Отростки блуждающего нерва, проходящие в составе сердечных нервов, представляют собой парасимпатические преганглионарные волокна.
С них возбуждение передается на интрамуральные нейроны и далее — преимущественно на элементы проводящей системы. Влияния, опосредованные правым блуждающим нервом, адресованы, в основном, клеткам синоатриального, а левым — клеткам атриовентрикулярного узла. Прямого влияния на желудочки сердца блуждающие нервы не оказывают. Иннервируя ткань водителей ритма, вегетативные нервы способны менять их возбудимость, тем самым вызывая изменения частоты генерации потенциалов действия и сокращений сердца (хронотропный эффект).
Нервные влияния изменяют скорость электротонической передачи возбуждения и, следовательно, длительности фаз сердечного цикла. Такие эффекты называют дромотропными. Поскольку действие медиаторов вегетативной нервной системы заключается в изменении уровня циклических нуклеотидов и энергетического обмена, вегетативные нервы в целом способны влиять и на силу сердечных сокращений (инотропный эффект). В лабораторных условиях получен эффект изменения величины порога возбуждения кардиомиоцитов под действием нейромедиаторов, его обозначают как батмотропный.
Перечисленные пути воздействия нервной системы на сократительную активность миокарда и насосную функцию сердца представляют собой хотя и исключительно важные, но вторичные по отношению к миогенным механизмам модулирующие влияния.
57. Парасимпатическая регуляция сердца. Влияние блуждающего нерва на сердце. Механизм его отрицательного хронотропного действия.
При раздражении сердечных нервов под влиянием медиатора изменяется мембранный потенциал мышечных волокон сердечной мышцы. При раздражении блуждающего нерва происходит гиперполяризация мембраны, т.е. увеличивается мембранный потенциал. Основу гиперполяризации сердечной мышцы составляет увеличение проницаемости мембраны для ионов калия.
Зная, что блуждающий нерв гиперполяризует, а симпатический — деполяризует мембрану, можно объяснить все эффекты действия этих нервов на сердце. Поскольку при раздражении блуждающего нерва увеличивается мембранный потенциал, требуется большая сила раздражения для достижения критического уровня деполяризации и получения ответной реакции, а это свидетельствует об уменьшении возбудимости (отрицательный батмотропный эффект).
Отрицательный хронотропный эффект связан с тем, что при большой силе раздражения вагуса гиперполяризация мембраны столь велика, что возникающая спонтанная деполяризация не может достичь критического уровня и ответ не возникает — наступает остановка сердца.
При малой частоте или силе раздражения блуждающего нерва степень гиперполяризации мембраны меньше и спонтанная деполяризация постепенно достигает критического уровня, вследствие чего наступают редкие сокращения сердца (отрицательный дромотропный эффект).
58. Симпатическая регуляция сердца. Влияние симпатических нервов на сердце. Механизм положительного инотропного действия «усиливающего» нерва И.П. Павлова.
Первые детальные исследования симпатической нервной системы на деятельность сердца принадлежит братьям Цион (1867), а затем И.П. Павлову (1887).
Братья Цион наблюдали увеличение частоты сердечных сокращений при раздражении спинного мозга в области расположения нейронов, регулирующих деятельность сердца. После перерезки симпатических нервов такое же раздражение спинного мозга не вызывало изменений деятельности сердца. Было установлено, что симпатические нервы, иннервирующие сердце, оказывают положительное влияние на все стороны деятельности сердца. Они вызывают положительные хронотропный, инотропный, батморопный, дромотропный и тонотропный эффекты.
Дальнейшими исследованиями И.П. Павлова было показано, что нервные волокна, входящие в состав симпатического и блуждающего нервов, влияют на разные стороны деятельности сердца: одни изменяют частоту, а другие — силу сердечных сокращений. Веточки симпатического нерва, при раздражении которых наступает усиление силы сердечных сокращений, были названы усиливающим нервом Павлова. Было установлено, что усиливающее влияние симпатических нервов связано с повышением уровня обмена веществ.
В составе блуждающего нерва также были найдены волокна, влияющие только на частоту и только на силу сердечных сокращений.
На частоту и силу сокращений влияют волокна блуждающего и симпатического нервов, подходящие к синусному узлу, а сила сокращений изменяется под влиянием волокон, подходящих к атриовентрикулярному узлу и миокарду желудочков.
Блуждающий нерв легко адаптируется к раздражению, поэтому его эффект может исчезнуть, несмотря на продолжающееся раздражение. Это явление получило название «ускользание сердца от влияния вагуса». Блуждающий нерв обладает более высокой возбудимостью, вследствие чего он реагирует на меньшую силу раздражения, чем симпатический, и коротким латентным периодом.
Поэтому при одинаковых условиях раздражения эффект блуждающего нерва появляется раньше, чем симпатического
Ветви симпатических нервов берут начало от грудного отдела спинного мозга и прерываются в верхнем, среднем шейных и звездчатых ганглиях. Постганглионарные волокна иннервируют весь миокард, но в основном предсердия. Раздражение симпатических нервов оказывает влияние, противоположное действию блуждающих нервов: увеличиваются частота и сила сердечных сокращений, улучшается проводимость и повышается возбудимость.
Указанные процессы обусловлены взаимодействием медиатора норадреналина преимущественно с β-адренорецепторами миокарда. Его действие связано с ростом мембранной проницаемости для ионов кальция, что сопровождается повышением степени сопряженности возбуждения и сокращения миокарда. Кроме того, адреналин способствует расщеплению гликогена и образованию глюкозы, служащей источником энергии для сокращения миокарда. Норадреналин разрушается значительно медленнее, чем ацетилхолин, поэтому его взаимодействие с адренорецепторами сердечных клеток, как и раздражение симпатических волокон, сопровождается более продолжительным эффектом. При чрезмерном возбуждении сердечных симпатических нервов возбудимость миокарда может возрасти настолько, что появляются эктопические очаги возбуждения и нарушения ритма.
При раздражении симпатического нерва даже небольшой силой возникает деполяризация мембраны, которая характеризуется уменьшением величины мембранного и порогового потенциалов, что свидетельствует о повышении возбудимости (положительный батмотропный эффект). Поскольку под влиянием симпатического нерва мембрана мышечных волокон сердца деполяризуется, время спонтанной деполяризации, необходимой для достижения критического уровня и возникновения потенциала действия, уменьшается, что приводит к увеличению частоты сердечных сокращений.
59. Рефлекторная регуляция сердца. Роль гипоталамуса, лимбической системы и коры головного мозга в рефлекторной регуляции сердца.

Ядра гипоталамуса являются следующей ступенью в иерархии нервных центров, регулирующих сердечную деятельность. У млекопитающих посредством прямых связей гипоталамус соединяется с ядами блуждающих нервов в продолговатом мозге и боковыми рогами спинного мозга, т.е. с центрами симпатической и парасимпатической иннервации сердца. Непосредственное участие в регуляции работы сердца принимают паравентрикулярное, вентромедиальное и преоптические ядра, сосцевидные тела.
Гипоталамус – интегративный центр, который может изменять параметры сердечной деятельности для обеспечения текущих потребностей организма и всех его систем при различных поведенческих реакциях.
Корковая регуляция
Кора головного мозга является высшей ступенью в иерархии механизмов целенаправленного управления деятельностью сердца. Они проявляются особенно отчетливо при раздра жении моторной и премоторной зон коры, поясной извилины, орбитальной поверхности лобных долей, передней части височной доли. В то же время электрическая стимуляция практически любого участка коры мозга вызывает ответы сер дечно-сосудистой системы. Возможность произвольной корковой регуляции подтверждается экспериментами с БОС.
Кора головного мозга является органом психической деятельности, обеспе чивающим целостные приспособительные реакции организма. Многочислен ные наблюдения показывают, что работа сердца изменяется при действии усло вий, влияющих на деятельность коры мозга: боли, страха, радости, тревоги, ярости и т. п. Например, у спортсменов наблюдается так называемое предстар товое состояние, проявляющееся учащением сердцебиения. Оно связано с активацией корой мозга гипоталамо-гипофизарной системы и надпочечников, выделяющих адреналин в кровь. Сигналы, непосредственно предвещающие возникновение этих ситуаций или возможность их наступления, способны по механизму условного рефлекса вызвать перестройку функций сердца, чтобы обеспечить предстоящую деятельность организма
Корковая регуляция осуществляется посредством гипоталамических влияний и изменения активности лимбической системы, которые, в свою очередь, реализуют свои эффекты через автономную нервную систему.
Все условные рефлексы являются приобретенными в процессе жизни каждого организма и формируются они с участием коры больших полушарий. Трудно, а может быть даже и невозможно выработать условный рефлекс, который бы проявлялся изолированным изменением только деятельности сердца. В целостном организме изменения функций носят системный характер, т.е. наблюдается одновременное изменение ряда показателей. Например, предстартовое изменение ряда функций организма имеет несомненно условнорефлекторный характер, но при этом усиливается не только работа сердца, а происходит повышение АД, гипервентиляция, повышение активности ЦНС, вегетативной нервной системы, увеличение количества эритроцитов и гемоглобина, усиление функций желез внутренней секреции.
Все эти изменения необходимы для достижения полезного приспособительного результата, а в данном случае — высокого спортивного результата.
Или другой пример, который также имеет приобретенный, т.е. условнорефлекторный характер. Изменение функционального состояния перечисленных систем четко проявляется у студентов и школьников во время сдачи экзаменов. Причем уровень психо-эмоционального возбуждения неодинаковый в разные периоды предэкзаминационного напряжения, чем больше дефицит информации, тем больше выраженность реакций (перед взятием билета, начало ответа и т.д.). Уровень мотиваций, выраженность эмоционального компонента имеют чрезвычайно важное значение для выработки условных рефлексов. Поэтому все эти сложные поведенческие реакции протекают с участием не только коры головного мозга, но и подкорковых, и прежде всего лимбических и гипоталамических структур мозга. Роль коры при этом заключается, главным образом, в оценке путей реализации и правильности результата действия.
Условные рефлексы могут быть натуральными (естественными — вышеприведенные примеры относятся к этой категории рефлексов) и искусственными, т.е. выработанные в лабораторных условиях на какие-то ранее индифферентные раздражители. Например, в качестве условного сигнала используется «свет», а подкреплением является болевое раздражение (которое сопровождается в частности тахикардией и повышением АД и т.д.). После выработки условного рефлекса условный сигнал «свет» будет вызывать такие же изменения, как и боль.
Условные рефлексы, образованные путем сочетания условного сигнала с безусловным раздражителем, получили название условных рефлексов первого порядка. Те условные рефлексы, которые образованы на основе сочетания внешнего агента с условным сигналом, вызывающим выработанный ранее прочный постоянный условный рефлекс первого порядка, называются условными рефлексами второго порядка.
Путем сочетания индифферентного раздражителя с условным раздражителем второго порядка можно выработать у собаки условный рефлекс третьего порядка. Последний удалось наблюдать в опытах с оборонительными двигательными рефлексами, которые вызывались электрическим раздражением лапы. Условных рефлексов четвертого порядка у собаки выработать не удается. У детей описаны рефлексы шестого порядка.
Изучение природы временной связи условного рефлекса открывает путь к познанию основы высших функций мозга — явлений памяти. Однако несмотря на многочисленные исследования, до сих пор очень многое остается неясным.
Структура условного рефлекса. Временная связь, лежащая в основе условного рефлекса, имеет «горизонтальную» структуру и замыкается в пределах коры между центром сигнального раздражителя и клетками коркового представительства безусловного рефлекса. Временные связи образуются, как правило, на корковом уровне, однако при исключении такой возможности, например при разрезах коры, могут осуществляться по путям нижележащих уровней (рис. 235, I).
Многие особенности выработки условных рефлексов получают объяснение, если принять, что при этом формируются множественные прямые и обратные связи как в пределах коры, так и между корой и подкорковыми центрами
Важное место в процессе коркового анализа и синтеза раздражений занимает явление условнорефлекторного переключения. Сущность его заключается в том, что эффект условного раздражителя (его сигнальное значение) может быть поставлен в определенную зависимость от той обстановки, в которой он применяется.
Так, если какой-либо звуковой агент, например метроном с частотой 120 ударов в минуту, в утренние часы сопровождать подачей животному пищи, а в дневные — электрическим раздражением конечности, то такой раздражитель после ряда сочетаний приобретает различное сигнальное значение в зависимости от времени дня: утром он будет вызывать условную пищевую реакцию, а днем — оборонительную. Время дня оказывается фактором, определяющим характер условной реакции, как бы переключающим кору большого мозга с одного вида деятельности на другой.
Опыты показывают, что «переключателями» могут быть разнообразные раздражители, связанные с обстановкой экспериментов или самим экспериментатором. В различной обстановке (разные комнаты) один и тот же раздражитель (например, гудок) может являться то условным побуждающим, то тормозным условным сигналом.
60. Собственные рефлексы регуляции сердца с сосудистых рефлексогенных зон.
С механорецепторов легочной артерии, как показал В. В. Парии (1946), возникает рефлекс, аналогичный рефлексам с рефлексогенных зон аорты, каротидного синуса и самого сердца.
Таким образом, механорецепторы эндокарда полостей сердца и ближайших к нему крупных сосудов на выходе (дуга и корень аорты, область каротидного синуса, легочные, коронарные, щитовидные, подключичные артерии) представляют собой единое рецепторное поле, «обуздывающее» артериальное давление. Единое рецепторное поле представляют и хеморецепторы аортального, каротидного и подключичного тельца.
На входе в сердце, в стенках полых вен, возможно, также имеется зона механорецепции. При растяжении устьев полых вен избытком крови наблюдается ответная реакция в виде тахикардии, которая способствует более быстрому перекачиванию крови из венозного отдела сосудистого русла в артериальный. Реакция была описана Бейнбриджем и получила название рефлекса Бейнбриджа. Механизм данной реакции до конца не ясен. Вывод о рефлекторной природе этой реакции сделан на основании того, что тахикардия при растяжении полых вен сопровождается возрастанием импульсации в эфферентных симпатических нервах сердца и исчезает после перерезки блуждающих нервов. Предполагают, что афферентная часть дуги рефлекса Бейнбриджа представлена волокнами блуждающего нерва, а эфферентная — симпатическими нервами. Тахикардия при растяжении полых вен и правого предсердия наблюдается и на полностью денервированном сердце, следовательно, в возникновении этой реакции нельзя исключить механизм Стерлинга и периферические сердечные рефлексы.
Все собственные системные рефлексы сердечно-сосудистой системы осуществляются с использованием механизма отрицательной обратной связи и относятся к рефлексам саморегуляции. Они обеспечивают устойчивое состояние основных параметров системы кровообращения.
Синокаротидный рефлекс (Чермака). В области каротидного синуса (область бифуркации сонной артерии) расположена рецепторная зона, содержащая барорецепторы, реагирующие на изменение давления. При повышении давления в сосуде в результате надавливания в рецепторах возникают импульсы, которые по чувствительным волокнам поступают в продолговатый мозг. В результате увеличивается активность ядер блуждающих нервов, снижается частота сердечных сокращений.
Депрессорный синокаротидный рефлекс развивается при повышении давления крови в синокаротидной зоне, его рефлекторная дуга построена таким образом. Информация об активации (растяжение) барорецепторов по нерву Геринга (увеличено количество ПД) по каналу отрицательной обратной связи поступает в каудальную вентролатерального часть продолговатого мозга и с помощью медиатора глутамата возбуждает чувствительные ядра блуждающего нерва - n. tractus solitarius (NTS). Из них информация поступает в каудальную вентролатеральную L-зону, в которой контактирует с тормозным интернейрон, передающий импульсы (ПД) в клювовидный вентролатеральный отдел продолговатого мозга (симпатоактивной зоны М и S), где с помощью медиатора ГАМК тормозит нейроны симпатической нервной системы, которые несут информацию в боковые рога спинного мозга, а оттуда до сосудов и сердца. В результате торможения влияния симпатической нервной системы существенно снижается ее тонизирующее воздействие на сосуды и сердце, в результате чего сосуды расширяются, уменьшается работа сердца и как результат - наступает падение кровяного давления. Параллельно информация по чувствительным волокнам n. tractus solitarius (NTS) направляется в двигательное ядро блуждающего нерва – n. ambiguus (каудальная вентролатеральная зона), а от него к сердцу, которое реагирует отрицательным хроноинотропным эффектом - уменьшением сердечного выброса крови в сосуды, что также приводит к снижению артериального давления.
Прессорный синокаротидного рефлекс развивается при снижении давления крови в синокаротидной зоне (сморщивании барорецепторов). Уменьшение информации от рецепторов при гипотонии (количество ПД в нерве Геринга значительно падает) по каналу отрицательной связи передается в чувствительные ядра NTS, а из них в ростральную вентролатеральную часть, которая резко усиливает свою активность, что приводит к увеличению частоты импульсов в симпатических волокнах, сужение кровеносных сосудов и повышение артериального давления. Параллельно информация направляется к сердцу и усиливает его работу, что приводит к увеличению выброса крови в сосуды, вследствие этого также возрастает артериальное давление.
При повышении кровяного давления в дуге аорты возбуждаются барорецепторы, информация (ПД) от которых по каналу отрицательной обратной связи - депрессорные (парасимпатической) нерва (X пара Ч.Н.) поступает в каудальный вентролатеральном участок. Стимулируя его, он вызывает активацию тормозного вставного нейрона, который блокирует симпатоактивные нейроны рострального вентролатерального участка. В результате частота импульсов, идущих по симпатических нервных волокнах к сосудам, резко уменьшается или прекращается, они расширяются, артериальное давление падает. Одновременно возбуждаются моторные ядра блуждающего нерва, находящихся в каудальном вентролатеральном участке. Информация от них по блуждающих нервах направляется к сердцу и вызывает снижение выброса крови, что также способствует снижению артериального давления. Таким образом, сосудодвигательный центр находится под постоянным воздействием со стороны барорецепторов и благодаря им пристально следит за уровнем артериального давления в организме.
Рефлексы с хеморецепторов синокаротидной и аортального рефлексогенных зон возникают при отклонении от нормы регулируемых параметров организма; увеличении в артериальной крови Рсо2, ионов водорода [Н +], уменьшении в каротидном синусе Ро2 до 60 мм рт. ст. Хеморецепторы каротидного синуса также возбуждаются под влиянием цианидов, никотина и других соединений, которые поступают к ним с кровью.
Возбуждение хеморецепторов приводит к стимуляции как прессорных симпатоактивных М и S долей ростральной вентролатеральной зоны продолговатого мозга, в результате чего сужаются сосуды, так и к каудальной депрессорного L-зоны, что приводит к снижению частоты сердечных сокращений, уменьшения сердечного выброса крови. Однако эффекты, связанные с сужением сосудов, преобладают над снижением МОК, что приводит к росту системного артериального давления и, вследствие этого, ускорение линейной скорости движения крови, ускорение вывода углекислого газа и увеличение напряжения кислорода в артериальной крови, направленных на поддержание газового гомеостаза организма. В природных условиях эти реакции накладывается влияние системы дыхания, а также прямое действие вышеназванных факторов на сосуды.
61. Сопряженные рефлексы регуляции сердца (рефлекс Гольца, рефлекс Ашнера-Данини).
ГЛАЗОСЕРДЕЧНЫЙ (АШНЕРА-ДАНИНИ) РЕФЛЕКС. Надавливание на глазные яблоки вызывает раздражение рецепторов тройничного нерва; импульсы передаются по этому нерву в средний мозг, далее по вставочному нейрону (возможно, по волокнам ретикулярной субстанции) направляются к ядрам блуждающих нервов, от которых импульсы доходят до нервных сплетений сердца. Кроме того, надавливание на глаза приводит к изменению внутриглазного давления и раздражению барорецепторов полостей глаз, импульсы от которых передаются в ствол мозга к ядрам блуждающих нервов.

Умеренное и равномерное надавливание на глазные яблоки приводит к рефлекторному снижению частоты пульса в среднем на 4-8 биений в минуту.
Техника может использоваться для:
- диагностики (самодиагностики) степени возбудимости парасимпатического отдела вегетативной нервной системы, по результатам которой можно предполагать о том, находитесь ли вы в оптимальном адаптивном состоянии, стрессе.
- снятия приступа тахикардии (повышенной частоты сердечных сокращений, например, при панических атаках) – то есть, для нормализации работы сердца, снижения артериального давления, снижения частоты дыхания и, в целом, снижения психоэмоционального возбуждения.
РЕФЛЕКС ГОЛЬТЦА — уменьшение ЧСС или даже полная остановка сердца при раздражении механорецепторов органов брюшной полости или брюшины, что учитывается при хирургических вмешательствах в брюшной полости. В опыте Гольца поколачивание по желудку и кишечнику лягушки ведет к остановке сердца.
62. Экстракардиальная гуморальная регуляция нагнетательной функции сердца. Роль разных гормонов и электролитов в регуляции сердца.
Факторы гуморальной регуляции делят на две группы:
1) вещества системного действия;
2) вещества местного действия.
История создания датчика движения: Первый прибор для обнаружения движения был изобретен немецким физиком Генрихом Герцем...

История развития пистолетов-пулеметов: Предпосылкой для возникновения пистолетов-пулеметов послужила давняя тенденция тяготения винтовок...

Биохимия спиртового брожения: Основу технологии получения пива составляет спиртовое брожение, - при котором сахар превращается...

Особенности сооружения опор в сложных условиях: Сооружение ВЛ в районах с суровыми климатическими и тяжелыми геологическими условиями...
© cyberpedia.su 2017-2024 - Не является автором материалов. Исключительное право сохранено за автором текста.
Если вы не хотите, чтобы данный материал был у нас на сайте, перейдите по ссылке: Нарушение авторских прав. Мы поможем в написании вашей работы!