Задание 9
Назовите к какому механическому результату приводит:
а) увеличение физиологического поперечника мышцы
Сила тяги мышцы зависит от совокупности механических, анатомических и физиологических условий.
Основным механическим условием, определяющим тягу мышцы, является нагрузка. Без нагрузки на мышцу не может быть ее силы тяги. Нагрузка растягивает мышцу при ее уступающей работе. Против нагрузки мышца выполняет преодолевающую работу. С нарастанием нагрузки сила тяги мышцы увеличивается, но не беспредельно. Нагрузка может быть представлена весом отягощения, а также его силой инерции и другими силами. Большее ускорение отягощения вызывает большую силу инерции. Следовательно, и при не очень большом отягощении, увеличивая его ускорение, можно увеличивать нагрузку, а значит, и силу тяги мышцы.
Движение звеньев в кинематической цепи как результат приложения тяги мышцы зависит также от: а) закрепления звеньев; б) соотношения сил, вызывающих движение, и сил сопротивления; в) начальных условий движения. При различных условиях закрепления звеньев в паре одна и та же тяга приводит к неодинаковому результату — разным движениям звеньев в суставе. В биокинематической паре может быть закреплено одно или другое звено, либо оба свободны, либо оба закреплены Соответственно возникнут ускорения одного из звеньев, либо обоих вместе (встречные движения), либо соединение будет фиксировано. Для двусуставной мышцы, не говоря уже о многосуставных, возможных вариантов намного больше. Наконец, особо важны для эффекта тяги мышцы начальные условия движения — положение звеньев пары и их скорость (направление и величина) в момент приложения силы тяги мышцы.
Из анатомических условий проявления тяги мышцы надо назвать строение мышцы и ее расположение (в данный момент движения). Физиологический поперечник мышцы (площадь сечения через все волокна перпендикулярно к их продольным осям) определяет
суммарную тягу всех волокон с учетом их взаимного расположения. От расположения волокон зависит и величина их упругой деформации при растягивании всей мышцы, а значит, и величина возникающих упругих сил. Расположение мышцы в каждый момент движения определяет угол ее тяги относительно костного рычага и величину растягивания, что влияет на величину момента силы тяги мышцы. При углах, отличающихся от прямого, как уже говорилось, кроме вращающей имеется и укрепляющая составляющая тяги мышцы; с увеличением укрепляющей уменьшается вращающая.
Физиологические условия проявления тяги мышцы в основном можно свести к ее возбуждению и утомлению. Эти два фактора отражаются на возможностях мышцы, повышая или снижая ее силу тяги.
Величина силы тяги мышцы связана с быстротой ее продольной деформации. С увеличением скорости сокращения мышцы при преодолевающей работе ее сила тяги уменьшается. При уступающей же работе увеличение скорости растягивания мышцы увеличивает ее силу тяги. Это очень важно для оценки силы тяги мышцы при быстрых движениях.
В биокинематических цепях действуют те же факторы, определяющие результат тяги каждой мышцы. Но так как в биокинематической цепи все звенья так или иначе взаимосвязаны, то в каждом конкретном случае лишь совокупность всех факторов определяет результат работы мышц в целом.
Разновидности работы мышц определяются сочетанием изменений их силы тяги и длины.
Виды работы мышц (преодолевающая, уступающая) определяются только характером изменения длины всей мышцы: укорочением, удлинением или же сохранением длины. Для каждого из этих трех случаев существует возможность по меньшей мере трех вариантов изменения силы тяги: увеличения, уменьшения или сохранения ее постоянной. Таким образом, схематически можно выделить девять типичных разновидностей работ
б) увеличение длины мышцы
Мышцы могут проявлять силу: без изменения своей длины (изометрический режим), при уменьшении длины (изотонический режим), при удлинении (эксцентрический режим), при использовании специальной аппаратуры возможно соблюдение изокинетического режима (в ходе сокращения мышц соблюдается либо постоянная скорость, либо сила).
Силовое проявление мышцы зависит от:
— интенсивности активации мотнейронного пула спинного мозга данной мышцы;
— количества активированных двигательных единиц и мышечных волокон;
— количества миофибрилл в каждом мышечном волокне;
— скорости сокращения миофибрилл, которая зависти от активности миозиновой АТФ-азы и величины внешнего сопротивления;
— законов механики мышечного сокращения (сила — длина мышцы, сила — скорость сокращения);
— начального состояния исполнительного аппарата (утомленные мышечные волокна демонстрируют меньшую силу).
Спортсмен при желании сократить какую-либо мышцу активизирует соответствующий двигательный нейрон в коре головного мозга, который посылает импульсы в спинной мозг к мотонейронному пулу, обслуживающему данную мышцу. Поскольку в мотонейронном пуле размеры мотонейронов различаются, то при низкой частоте импульсации из ЦНС могут активироваться только низкопороговые мотонейроны. Каждый мотонейрон иннервирует свои мышечные волокна. Поэтому активация мотонейрона приводит к рекрутированию или возбуждению соответствующих мышечных волокон. Каждое активное мышечное волокно под влиянием электрических импульсов выпускает из СПР ионы кальция, которые снимают ингибитор с активных центров актина. Это обеспечивает образование актин-миозиновых мостиков и начало их поворота и мышечного сокращения. На поворот мостиков и отсоединение актина от миозина тратится энергия одной молекулы АТФ. Продолжительность работы мостика составляет 1 мс. Вероятность образования мостиков зависит от взаимного расположения между собой нитей актина и миозина, отсюда возникает зависимость сила — длина активной мышца, а также от скорости взаимного перемещения (скольжения) их одной по отношению к другой, соответственно, имеем зависимость «сила-скорость».
Зависимость «сила — длина активного мышечного волокна» определяется, как правило, относительным расположением между собой головок миозина и активных центров актина. Максимальное количество мостиков возникает при некоторой средней длине мышцы. Отклонение от этой длины в большую или меньшую сторону ведет к снижению силовых проявлений мышечного волокна (мышцы). Однако, в случае растяжения некоторых мышц, еще не в активном состоянии, могут возникать значительные силы сопротивления растяжению, например, в мышцах сгибателях голеностопного или лучезапястного сустава. Эти силы связаны с растяжением соединительных тканей, например, перемезиума. В биомеханике в таком случае говорят о параллельном упругом компоненте мышцы. Упругостью обладают сухожилия, зет-пластинки саркомеров и нити миозина, к которым прикреплены головки. Такую упругость называют последовательной упругой компонентой.
Наличие последовательной упругой компоненты в мышечных волокнах приводит к тому, что с ростом числа рекрутированных МВ увеличивается жесткость мышцы -коэффициент упругости (В. М. Зациорский с соав., 1981).
Растягивание активной мышцы приводит не только к накоплению энергии упругой деформации в последовательной упругой компоненте, но и к прекращению работы мостиков, а именно, они перестают отцепляться за счет энергии молекул АТФ. Разрыв мостиков происходит благодаря действию внешней — механической силы. В итоге отрицательная работа мышц выполняется с очень высоким коэффициентов полезного действия, с минимальными затратами АТФ, а значит и кислорода.
в) увеличение длины саркомера
Существует ли какое-нибудь свидетельство механических свойств мышцы, поддерживающее существование сверхрастянутых саркомеров в поврежденный мышце? Было выдвинуто предположение, что присутствие сверхрастянутых саркомеров увеличивает последовательную податливость, приводя к изменению зависимости «длина-напряжение» активной мышцы в направлении б о льших значений длины мышцы (Morgan, 1990). Такое изменение было сначала описано Katz (1939) а затем показано для изолированных мышечных волокон лягушки (Morgan et al. 1996), целой мышцы земноводных (Wood et al. 1993; Talbot & Morgan, 1996) и мышцы человека (Jones et al. 1997; Brockett et al. 2001). Моделируемый пример показан на рис. 3A.
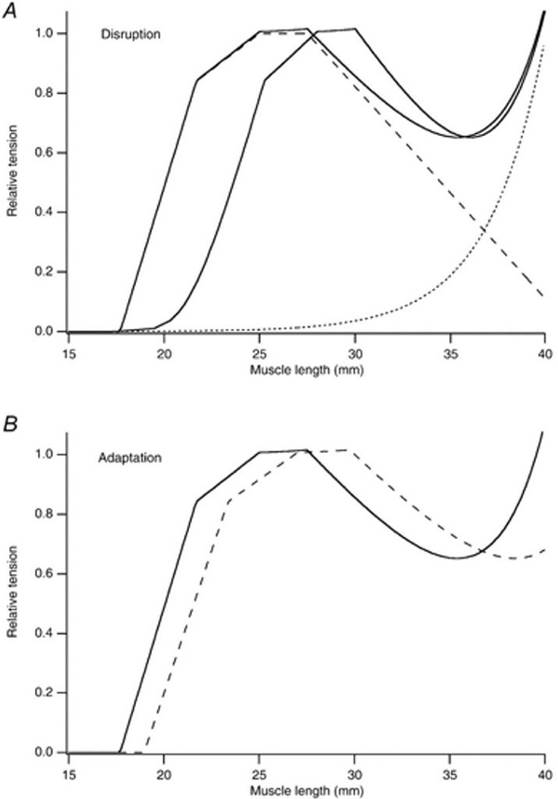
Рис.3. Изменения в механических свойствах мышцы после ряда эксцентрических сокращений.
A, разрушение саркомеров. Компьютер моделировал зависимость длина-напряжение; для саркомера. Штриховая линия; зависимость длина-напряжение для саркомера, взятое из статьи Gordon et al. (1966).
Пунктир — показательная кривая, представляющая пассивную напряженность; непрерывная линия — полная напряжение. Напряжение нормализовано относительно максимума активного напряжения. Волокно состоит из 10 000 саркомеров, имеющих среднюю длину 2,5 мкм. Исходная зависимость непрерывная кривая слева. После ряда эксцентрических сокращений 10 % саркомеров не могут развивать силу, поэтому чтобы моделировать разрушение, происходит изменение в оптимальной длине зависимости длина-напряжение на 3 мм вправо (непрерывная кривая справа).
B, адаптация мышечного волокна после повреждения, вызванного эксцентрическим упражнением. Непрерывная кривая — кривая напряжения общего количества контроля как в верхней группе, расплющенной кривой, что после того, как число саркомеров последовательно было увеличено на 10 %, не изменяя длину сухожилия. Это привело к увеличению оптимальной длины на 2 мм.
Если основной причиной дефицита напряжения после эксцентрических сокращений был отказ в сцепления E-C (Warren et al. 1993), приводя к сниженному, но однородному выходу ионов Ca2+, то изменение в отношении «длина-напряжение» могло бы попасть на сниженный уровень активации, так, чтобы мышцу нужно было бы больше растянуть, чтобы она достигла максимальной активации (Endo, 1973). Существуют доказательства, что такое объяснение не всегда правильно. Для изолированного мышечного волокна лягушки (Morgan et al. 1996) и целой мышцы крысы (Рис. 4), могут быть найдены примеры, где кривые «длина-напряжение» до и после эксцентрического сокращения пересекаются при значительной длине (Katz, 1939; see also Brockett et al. 2001a, b).
При этих больших значениях длины напряжение после выполнения упражнения выше, чем до упражнения. Таким образом, чтобы объяснить изменение в кривой (рис. 4) нельзя использовать ссылку на неполную активацию мышцы.
г) увеличение активности миозиновой АТФ - азы. 6
В гликолитических мышечных волокнах имеется запас молекул АТФ в миофибриллах, запас молекул АТФ около митохондрий, запас молекул АТФ в саркоплазме. Имеется запас молекул КрФ, глобул гликогена и капелек жира. Масса митохондрий в гликолитических МВ (ГМВ) мала, поскольку необходима только для жизни этих клеток в покое.
Активизация биохимических процессов начинается с момента прохождения электрических импульсов по мембранам МВ. Открываются поры в цистернах, выходит кальций в саркоплазму, кальций прикрепляется к актину, образуются актин-миозиновые мостики, тратится АТФ и КрФ. Свободный креатин и неорганический фосфат выходят из миофибрилл и используют энергию саркоплазматических молекул АТФ для ресинтеза КрФ. Молекулы АТФ ресинтезируются в ходе анаэробного гликолиза. Гликолиз начинается с разрушения молекулы глюкозы или гликогена, а заканчивается образованием пирувата. Пируват, из-за отсутствия митохондрий, преобразуется в лактат. Соединение аниона лактата с протоном водорода приводит к образованию молочной кислоты, которая может в таком виде выходить в кровь. В крови молекула молочной кислоты диссоциирует, поэтому между концентрацией водорода и лактата имеется высокая корреляционная связь (R = 0,99).
Ионы водорода образуются при распаде саркоплазматических и других молекул АТФ.
Активность ГМВ приводит к накоплению в саркоплазме продуктов метаболизма Н, Кр, Ф, Ла, Пир и др.
Запасов миофибриллярных АТФ хватает на 1–2 с, КрФ 5–20 с (в зависимости от режима сокращения и расслабления МВ). Затем усиливается гликолиз, но мощность его не более 50 % от максимума, а из-за накопления ионов водорода нарушается процесс образования актин-миозиновых мостиков и через 30 с они практически полностью перестают образовываться. Это явление обычно определяют как локальное мышечное утомление. ГМВ определяют как утомляемые мышечные волокна.
Окислительные мышечные волокна устроены точно также как и гликолитические мышечные волокна. Основное различие связано с массой митохондрий. В ОМВ масса митохондрий находится в предельном соотношении с миофибриллами, что обеспечивает максимальное потребление кислорода одним килограммом ОМВ около 0,3 л/мин.
Активизация ОМВ приводит к образованию актин — миозиновых мостиков и затратам энергии молекул АТФ. Концентрация миофибриллярных молекул АТФ поддерживается КрФ. Поддержание концентрации КрФ обеспечивается двумя путями:
— молекулами АТФ ресинтезируемыми в митохондриях,
— молекулами АТФ ресинтезируемыми в аэробном гликолизе.
Этот процесс развивается в течение 45–60 с. К этому времени одновременно может идти как гликолиз, так и окисление жиров. Но по мере функционирования митохондрий в саркоплазме накапливается цитрат, поэтому начинается ингибирование ферментов гликолиза и ОМВ полностью переходит на липолиз.
Липолиз использует запасы жира в капельках, запаса этого жира у нормальных людей хватает на 30–50 мин. Жирные кислоты крови медленно поступают в МВ, поэтому не могут полностью обеспечить мышечную деятельность высокой интенсивности.
Митохондрии поглощают АДФ, Ф, кислород, пируват, жирные кислоты, глицерол, ионы водорода и выделяют ресинтезированные молекулы АТФ, углекислый газ и воду. Поэтому ОМВ не закисляются, не утомляются.
Окисление жиров в ОМВ может прекратиться, если в саркоплазме появятся ионы лактата. В этом случае окисление жиров ингибируется, а лактат становится субстратом окисления. Лактат с помощью лактатдегидрогеназы сердечного типа превращается в пируват, а тот, через ацетил-коэнзима, поступает в митохондрии. Пируват также начинает образовываться в ходе гликолиза из глюкозы и гликогена.
Лактат может попасть в ОМВ только при одновременном функционировании ГМВ и ОМВ.
Биомеханические свойства мышечных волокон связаны с эмпирическими законами:
— «сила — длина»,
— «сила — скорость»,
— «сила — время активации»,
— «сила — время расслабления»,
— «сила — энергия упругой деформации».
Эти законы надо учитывать при анализе соревновательной деятельности
Список литературы
1. Григорьева Л.С., Козловская И.Б. Влияние невесомости и гипокинезии на скоростно-силовые свойства мышц человека // Космич. биол. и авиакосмич. мед. - 1987. - 21. - C. 27-30.
2. Коряк Ю.А. Исследование скоростно-силовых свойств мышечного аппарата человека // Резерв. возможности организма спортсменов (Каражанов Б.Б., ред.). - Алма-Ата: КазИФК, 1985. - C. 86-97.
3. Христова Л.Г., Гидиков А.А., Асланова И.Ф. и др. Влияние иммерсионной гипокинезии на потенциалы двигательных единиц мускулатуры человека // Косм. биол. и авиакосмич. мед. - 1988. - 22. - P. 39-43.
4. Antonutto G., Capelli C., Cirardis M. et al. Effects of microgravity on maximal power of lower limbs during very short efforts in humans // J. Appl. Physiol. - 1999. - 86. - P. 85-92.
5. Bell D.G. Jacobs I. Eleciro-mechanical response limes and rate of force development in males and females // Med. Sci. Sports Exerc. - 1986. - 18. - P. 31-36.
Задание 9
Назовите к какому механическому результату приводит:
а) увеличение физиологического поперечника мышцы
Сила тяги мышцы зависит от совокупности механических, анатомических и физиологических условий.
Основным механическим условием, определяющим тягу мышцы, является нагрузка. Без нагрузки на мышцу не может быть ее силы тяги. Нагрузка растягивает мышцу при ее уступающей работе. Против нагрузки мышца выполняет преодолевающую работу. С нарастанием нагрузки сила тяги мышцы увеличивается, но не беспредельно. Нагрузка может быть представлена весом отягощения, а также его силой инерции и другими силами. Большее ускорение отягощения вызывает большую силу инерции. Следовательно, и при не очень большом отягощении, увеличивая его ускорение, можно увеличивать нагрузку, а значит, и силу тяги мышцы.
Движение звеньев в кинематической цепи как результат приложения тяги мышцы зависит также от: а) закрепления звеньев; б) соотношения сил, вызывающих движение, и сил сопротивления; в) начальных условий движения. При различных условиях закрепления звеньев в паре одна и та же тяга приводит к неодинаковому результату — разным движениям звеньев в суставе. В биокинематической паре может быть закреплено одно или другое звено, либо оба свободны, либо оба закреплены Соответственно возникнут ускорения одного из звеньев, либо обоих вместе (встречные движения), либо соединение будет фиксировано. Для двусуставной мышцы, не говоря уже о многосуставных, возможных вариантов намного больше. Наконец, особо важны для эффекта тяги мышцы начальные условия движения — положение звеньев пары и их скорость (направление и величина) в момент приложения силы тяги мышцы.
Из анатомических условий проявления тяги мышцы надо назвать строение мышцы и ее расположение (в данный момент движения). Физиологический поперечник мышцы (площадь сечения через все волокна перпендикулярно к их продольным осям) определяет
суммарную тягу всех волокон с учетом их взаимного расположения. От расположения волокон зависит и величина их упругой деформации при растягивании всей мышцы, а значит, и величина возникающих упругих сил. Расположение мышцы в каждый момент движения определяет угол ее тяги относительно костного рычага и величину растягивания, что влияет на величину момента силы тяги мышцы. При углах, отличающихся от прямого, как уже говорилось, кроме вращающей имеется и укрепляющая составляющая тяги мышцы; с увеличением укрепляющей уменьшается вращающая.
Физиологические условия проявления тяги мышцы в основном можно свести к ее возбуждению и утомлению. Эти два фактора отражаются на возможностях мышцы, повышая или снижая ее силу тяги.
Величина силы тяги мышцы связана с быстротой ее продольной деформации. С увеличением скорости сокращения мышцы при преодолевающей работе ее сила тяги уменьшается. При уступающей же работе увеличение скорости растягивания мышцы увеличивает ее силу тяги. Это очень важно для оценки силы тяги мышцы при быстрых движениях.
В биокинематических цепях действуют те же факторы, определяющие результат тяги каждой мышцы. Но так как в биокинематической цепи все звенья так или иначе взаимосвязаны, то в каждом конкретном случае лишь совокупность всех факторов определяет результат работы мышц в целом.
Разновидности работы мышц определяются сочетанием изменений их силы тяги и длины.
Виды работы мышц (преодолевающая, уступающая) определяются только характером изменения длины всей мышцы: укорочением, удлинением или же сохранением длины. Для каждого из этих трех случаев существует возможность по меньшей мере трех вариантов изменения силы тяги: увеличения, уменьшения или сохранения ее постоянной. Таким образом, схематически можно выделить девять типичных разновидностей работ
б) увеличение длины мышцы
Мышцы могут проявлять силу: без изменения своей длины (изометрический режим), при уменьшении длины (изотонический режим), при удлинении (эксцентрический режим), при использовании специальной аппаратуры возможно соблюдение изокинетического режима (в ходе сокращения мышц соблюдается либо постоянная скорость, либо сила).
Силовое проявление мышцы зависит от:
— интенсивности активации мотнейронного пула спинного мозга данной мышцы;
— количества активированных двигательных единиц и мышечных волокон;
— количества миофибрилл в каждом мышечном волокне;
— скорости сокращения миофибрилл, которая зависти от активности миозиновой АТФ-азы и величины внешнего сопротивления;
— законов механики мышечного сокращения (сила — длина мышцы, сила — скорость сокращения);
— начального состояния исполнительного аппарата (утомленные мышечные волокна демонстрируют меньшую силу).
Спортсмен при желании сократить какую-либо мышцу активизирует соответствующий двигательный нейрон в коре головного мозга, который посылает импульсы в спинной мозг к мотонейронному пулу, обслуживающему данную мышцу. Поскольку в мотонейронном пуле размеры мотонейронов различаются, то при низкой частоте импульсации из ЦНС могут активироваться только низкопороговые мотонейроны. Каждый мотонейрон иннервирует свои мышечные волокна. Поэтому активация мотонейрона приводит к рекрутированию или возбуждению соответствующих мышечных волокон. Каждое активное мышечное волокно под влиянием электрических импульсов выпускает из СПР ионы кальция, которые снимают ингибитор с активных центров актина. Это обеспечивает образование актин-миозиновых мостиков и начало их поворота и мышечного сокращения. На поворот мостиков и отсоединение актина от миозина тратится энергия одной молекулы АТФ. Продолжительность работы мостика составляет 1 мс. Вероятность образования мостиков зависит от взаимного расположения между собой нитей актина и миозина, отсюда возникает зависимость сила — длина активной мышца, а также от скорости взаимного перемещения (скольжения) их одной по отношению к другой, соответственно, имеем зависимость «сила-скорость».
Зависимость «сила — длина активного мышечного волокна» определяется, как правило, относительным расположением между собой головок миозина и активных центров актина. Максимальное количество мостиков возникает при некоторой средней длине мышцы. Отклонение от этой длины в большую или меньшую сторону ведет к снижению силовых проявлений мышечного волокна (мышцы). Однако, в случае растяжения некоторых мышц, еще не в активном состоянии, могут возникать значительные силы сопротивления растяжению, например, в мышцах сгибателях голеностопного или лучезапястного сустава. Эти силы связаны с растяжением соединительных тканей, например, перемезиума. В биомеханике в таком случае говорят о параллельном упругом компоненте мышцы. Упругостью обладают сухожилия, зет-пластинки саркомеров и нити миозина, к которым прикреплены головки. Такую упругость называют последовательной упругой компонентой.
Наличие последовательной упругой компоненты в мышечных волокнах приводит к тому, что с ростом числа рекрутированных МВ увеличивается жесткость мышцы -коэффициент упругости (В. М. Зациорский с соав., 1981).
Растягивание активной мышцы приводит не только к накоплению энергии упругой деформации в последовательной упругой компоненте, но и к прекращению работы мостиков, а именно, они перестают отцепляться за счет энергии молекул АТФ. Разрыв мостиков происходит благодаря действию внешней — механической силы. В итоге отрицательная работа мышц выполняется с очень высоким коэффициентов полезного действия, с минимальными затратами АТФ, а значит и кислорода.
в) увеличение длины саркомера
Существует ли какое-нибудь свидетельство механических свойств мышцы, поддерживающее существование сверхрастянутых саркомеров в поврежденный мышце? Было выдвинуто предположение, что присутствие сверхрастянутых саркомеров увеличивает последовательную податливость, приводя к изменению зависимости «длина-напряжение» активной мышцы в направлении б о льших значений длины мышцы (Morgan, 1990). Такое изменение было сначала описано Katz (1939) а затем показано для изолированных мышечных волокон лягушки (Morgan et al. 1996), целой мышцы земноводных (Wood et al. 1993; Talbot & Morgan, 1996) и мышцы человека (Jones et al. 1997; Brockett et al. 2001). Моделируемый пример показан на рис. 3A.
Рис.3. Изменения в механических свойствах мышцы после ряда эксцентрических сокращений.
A, разрушение саркомеров. Компьютер моделировал зависимость длина-напряжение; для саркомера. Штриховая линия; зависимость длина-напряжение для саркомера, взятое из статьи Gordon et al. (1966).
Пунктир — показательная кривая, представляющая пассивную напряженность; непрерывная линия — полная напряжение. Напряжение нормализовано относительно максимума активного напряжения. Волокно состоит из 10 000 саркомеров, имеющих среднюю длину 2,5 мкм. Исходная зависимость непрерывная кривая слева. После ряда эксцентрических сокращений 10 % саркомеров не могут развивать силу, поэтому чтобы моделировать разрушение, происходит изменение в оптимальной длине зависимости длина-напряжение на 3 мм вправо (непрерывная кривая справа).
B, адаптация мышечного волокна после повреждения, вызванного эксцентрическим упражнением. Непрерывная кривая — кривая напряжения общего количества контроля как в верхней группе, расплющенной кривой, что после того, как число саркомеров последовательно было увеличено на 10 %, не изменяя длину сухожилия. Это привело к увеличению оптимальной длины на 2 мм.
Если основной причиной дефицита напряжения после эксцентрических сокращений был отказ в сцепления E-C (Warren et al. 1993), приводя к сниженному, но однородному выходу ионов Ca2+, то изменение в отношении «длина-напряжение» могло бы попасть на сниженный уровень активации, так, чтобы мышцу нужно было бы больше растянуть, чтобы она достигла максимальной активации (Endo, 1973). Существуют доказательства, что такое объяснение не всегда правильно. Для изолированного мышечного волокна лягушки (Morgan et al. 1996) и целой мышцы крысы (Рис. 4), могут быть найдены примеры, где кривые «длина-напряжение» до и после эксцентрического сокращения пересекаются при значительной длине (Katz, 1939; see also Brockett et al. 2001a, b).
При этих больших значениях длины напряжение после выполнения упражнения выше, чем до упражнения. Таким образом, чтобы объяснить изменение в кривой (рис. 4) нельзя использовать ссылку на неполную активацию мышцы.
г) увеличение активности миозиновой АТФ - азы. 6
В гликолитических мышечных волокнах имеется запас молекул АТФ в миофибриллах, запас молекул АТФ около митохондрий, запас молекул АТФ в саркоплазме. Имеется запас молекул КрФ, глобул гликогена и капелек жира. Масса митохондрий в гликолитических МВ (ГМВ) мала, поскольку необходима только для жизни этих клеток в покое.
Активизация биохимических процессов начинается с момента прохождения электрических импульсов по мембранам МВ. Открываются поры в цистернах, выходит кальций в саркоплазму, кальций прикрепляется к актину, образуются актин-миозиновые мостики, тратится АТФ и КрФ. Свободный креатин и неорганический фосфат выходят из миофибрилл и используют энергию саркоплазматических молекул АТФ для ресинтеза КрФ. Молекулы АТФ ресинтезируются в ходе анаэробного гликолиза. Гликолиз начинается с разрушения молекулы глюкозы или гликогена, а заканчивается образованием пирувата. Пируват, из-за отсутствия митохондрий, преобразуется в лактат. Соединение аниона лактата с протоном водорода приводит к образованию молочной кислоты, которая может в таком виде выходить в кровь. В крови молекула молочной кислоты диссоциирует, поэтому между концентрацией водорода и лактата имеется высокая корреляционная связь (R = 0,99).
Ионы водорода образуются при распаде саркоплазматических и других молекул АТФ.
Активность ГМВ приводит к накоплению в саркоплазме продуктов метаболизма Н, Кр, Ф, Ла, Пир и др.
Запасов миофибриллярных АТФ хватает на 1–2 с, КрФ 5–20 с (в зависимости от режима сокращения и расслабления МВ). Затем усиливается гликолиз, но мощность его не более 50 % от максимума, а из-за накопления ионов водорода нарушается процесс образования актин-миозиновых мостиков и через 30 с они практически полностью перестают образовываться. Это явление обычно определяют как локальное мышечное утомление. ГМВ определяют как утомляемые мышечные волокна.
Окислительные мышечные волокна устроены точно также как и гликолитические мышечные волокна. Основное различие связано с массой митохондрий. В ОМВ масса митохондрий находится в предельном соотношении с миофибриллами, что обеспечивает максимальное потребление кислорода одним килограммом ОМВ около 0,3 л/мин.
Активизация ОМВ приводит к образованию актин — миозиновых мостиков и затратам энергии молекул АТФ. Концентрация миофибриллярных молекул АТФ поддерживается КрФ. Поддержание концентрации КрФ обеспечивается двумя путями:
— молекулами АТФ ресинтезируемыми в митохондриях,
— молекулами АТФ ресинтезируемыми в аэробном гликолизе.
Этот процесс развивается в течение 45–60 с. К этому времени одновременно может идти как гликолиз, так и окисление жиров. Но по мере функционирования митохондрий в саркоплазме накапливается цитрат, поэтому начинается ингибирование ферментов гликолиза и ОМВ полностью переходит на липолиз.
Липолиз использует запасы жира в капельках, запаса этого жира у нормальных людей хватает на 30–50 мин. Жирные кислоты крови медленно поступают в МВ, поэтому не могут полностью обеспечить мышечную деятельность высокой интенсивности.
Митохондрии поглощают АДФ, Ф, кислород, пируват, жирные кислоты, глицерол, ионы водорода и выделяют ресинтезированные молекулы АТФ, углекислый газ и воду. Поэтому ОМВ не закисляются, не утомляются.
Окисление жиров в ОМВ может прекратиться, если в саркоплазме появятся ионы лактата. В этом случае окисление жиров ингибируется, а лактат становится субстратом окисления. Лактат с помощью лактатдегидрогеназы сердечного типа превращается в пируват, а тот, через ацетил-коэнзима, поступает в митохондрии. Пируват также начинает образовываться в ходе гликолиза из глюкозы и гликогена.
Лактат может попасть в ОМВ только при одновременном функционировании ГМВ и ОМВ.
Биомеханические свойства мышечных волокон связаны с эмпирическими законами:
— «сила — длина»,
— «сила — скорость»,
— «сила — время активации»,
— «сила — время расслабления»,
— «сила — энергия упругой деформации».
Эти законы надо учитывать при анализе соревновательной деятельности
Режим одиночного и тетанического сокращения. При каких условиях реализуется каждый из них; тетанический индекс и отчего он зависит
Режим одиночного и тетанического сокращения Одиночное – это сокращение, которое возникает на одиночный стимул, достаточный для вызова возбуждения мышцы. После короткого скрытого периода (латентный период) начинается процесс сокращения. При регистрации сократительной активности в изометрических условиях (два конца неподвижно закреплены) в первую фазу происходит нарастание напряжения (силы), а во вторую – ее падение до исходной величины. Соответственно эти фазы называют фазой напряжения и фазой расслабления. При регистрации сократительной активности в изотоническом режиме (например, в условиях обычной миографической записи) эти фазы будут называться соответственно фазой укорочения и фазой удлинения. Фаза укорочения всегда меньше по времени, чем фаза расслабления. Если на мышцу действует серия прямых раздражений (минуя нерв) или непрямых раздражений (через нерв), но с большим интервалом, при котором всякое следующее раздражение попадает в период после окончания 2-ой фазы, то мышца будет на каждый из этих раздражителей отвечать одиночным сокращением. В режиме одиночного сокращения мышца способна работать длительное время без развития утомления.
Суммированные возникают в том случае, если на мышцу наносятся 2 и более раздражения, причем всякое последующее раздражение (после предыдущего) наносится либо во время 2-ой фазы (расслабления или удлинения), либо во время 1-ой фазы (укорочения или напряжения). В случае, когда всякое второе раздражение попадает в период фазы расслабления (удлинения), возникает частичная суммация – сокращение еще полностью не закончилось, а уже возникло новое. Если попадается много раздражителей с подобным интервалом, то возникает явление зубчатого тетануса. Если раздражители наносятся с меньшим интервалом и каждое последующее раздражение попадает в фазу укорочения, то возникает так называемый гладкий тетанус.
Механическая работа, выполняемая мышцей, равна произведению развиваемой ею силы и расстояния, на протяжении которого она действует. Мощность мышечного сокращения отличается от силы мышцы, поскольку мощность является мерой общего количества работы, выполняемой в единицу времени. Следовательно, мощность определяется не только силой мышечного сокращения, но также расстоянием сокращения и числом сокращений в минуту. Мышечная мощность обычно измеряется в килограммометрах (кгм) в минуту. Например, о мышце, которая может поднимать вес, равный 1 кг, на высоту 1 м или сдвигать некий объект в сторону с силой 1 кг на расстояние 1 м за 1 мин, говорят, что ее мощность равна 1 кгм/мин.
Мышечная утомляемость – неспособность мышц поддерживать мышечное сокращение заданной интенсивности – связано с присутствием избытка аммиака, усиливающего анаэробный гликолиз, блокируя выход молочной кислоты. Повышение уровня аммиака и ацидоз лежат в основе метаболических нарушений при мышечной утомляемости. Эти процессы играют определяющую роль в формировании усталости, связанной с физиологическим утомлением.
Работоспособность скелетной мускулатуры и скорость развития утомления зависят от уровня умственной деятельности: высокий уровень умственного напряжения уменьшает мышечную выносливость.
При каких условиях реализуется каждый из них;
Основой всех типов мышечного сокращения служит взаимодействие актина и миозина. В скелетных мышцах за сокращение отвечают миофибриллы (примерно две трети сухого веса мышц). Миофибриллы — структуры толщиной 1-2 мкм, состоящие из саркомеров — структур длиной около 2,5 мкм, состоящих из актиновых и миозиновых (тонких и толстых) филаментов и Z-дисков, соединённых с актиновыми филаментами. Сокращение происходит при увеличении концентрации в цитоплазме ионов Ca2+ в результате скольжения миозиновых филаментов относительно актиновых. Источником энергии сокращения служит АТФ. КПД мышечной клетки около 50 %, мышцы в целом не более 20%. Максимальная сила мышц не достигается в реальных условиях; не все клетки мышцы используются одновременно и сокращаются с максимальной силой, иначе при сокращении многих скелетных мышц будут повреждены сухожилия или кости (что иногда и наблюдается при сильных судорогах). КПД мышцы также зависит от внешних условий; например, на холоде он значительно снижается, так как для организма важнее сохранить температуру тела[1].
Изометрическое сокращение. Изометрическое сокращение мышцы создаёт напряжение без изменения длины. Пример можно найти, когда мышцы руки и предплечья захватывают объект; суставы руки не двигаются, но мышцы генерируют достаточную силу, чтобы предотвратить падение объекта.
Изотоническое сокращение. При изотоническом сокращении напряжение в мышцах остаётся постоянным, несмотря на изменение длины мышц. Это происходит, когда сила сокращения мышц соответствует общей нагрузке на мышцы.
Концентрическое сокращение. При концентрическом сокращении мышечное напряжение является достаточным для преодоления нагрузки, а мышца укорачивается при сокращении. Это происходит, когда сила, создаваемая мышцей, превышает нагрузку, противодействующую её сокращению.
Эксцентрическое сокращение. При эксцентрическом сокращении напряжение недостаточно для преодоления внешней нагрузки на мышцы и мышечные волокна, удлиняются при их сокращении. Вместо того, чтобы тянуть сустав в направлении сокращения мышц, мышца действует так, чтобы замедлить сустав в конце движения или иным образом контролировать перемещение груза. Это может происходить невольно (например, при попытке переместить вес, слишком тяжёлый для подъёма мышцы) или добровольно (например, когда мышца «сглаживает» движение или сопротивляется гравитации, как, например, во время ходьбы вниз). В





