Лейкоциты, или белые кровяные тельца, представляют собой бесцветные клетки, содержащие ядро и протоплазму. Количество лейкоцитов в периферической крови колеблется в пределах 4х109/л - 9х109/л в зависимости от баланса гормонов, нервного напряжения, сезона, времени суток. Содержание лейкоцитов может быть увеличено (лейкоцитоз) или уменьшено (лейкопения). Среди физиологических лейкоцитозов различают пищевой, миогенный, эмоциональный, а также лейкоцитоз, возникающий при беременности. Физиологические лейкоцитозы носят перераспределительный характер и, как правило, не достигают высоких показателей. Имеется две группы лейкоцитов: гранулоциты (нейтрофилы, эозинофилы, базофилы) и агранулоциты (моноциты, лимфоциты). Размеры лейкоцитов варьируют от 4 мкм до 20 мкм. Продолжительность жизни гранулоцитов и моноцитов от 4-5 дней до 20 дней, лимфоцитов - до 100-120 дней. Лейкоциты обладают амебовидной подвижностью, миграцией (диапедезом) - способностью проникать через стенку неповрежденных капилляров - и фагоцитозом - способностью поглощать и переваривать микробов, чужеродные частицы и отмирающие клетки. Эти свойства определяют функции лейкоцитов: защитная (фагоцитоз, бактерицидное и антитоксическое действие, участие в иммунных реакциях, противоопухолевое действие); регенеративная -лейкоциты способствуют заживлению поврежденных тканей; транспортная - они являются носителями ряда ферментов.
Лейкограмма (лейкоцитарная формула) - это процентное соотношение различных видов лейкоцитов в крови: нейтрофилы -46-76%; эозинофилы - 1-5%; базофилы - 0-1%; моноциты -2-10%; лимфоциты - 18-40% (табл.2, рис.2).
Таблица 2

В том случае, когда отмечается сдвиг влево, но при этом фиксируется омоложение, это говорит о раковых заболеваниях органов кроветворения с метастазами в клетках костного мозга. Сдвиг формулы вправо возникает, когда количество зрелых клеток на порядок выше остальных. Это характерно для патологий печени и почек, лучевой болезни и частом переливании крови, при котором естественный баланс сдвигается. В результате проведения анализа также получают индекс сдвига, которые отображает цифру, определяющую соотношение общего числа юных лейкоцитов к зрелым кровяным тельцам. В норме у здорового человека это индекс находится в пределах 0,05-0,1.

Рис.2. Виды лейкоцитов
В отличие от взрослых количество лейкоцитов в крови, смотря какого возраста ребенок, разное. На протяжении всего детского периода жизни у ребенка дважды происходит перекрест лейкоцитарной формулы. Первый раз это случается после рождения малыша. Поскольку организм матери выполнял основную защитную функцию для плода, то состав крови новорожденного приближен к показателю нормы у взрослых людей. Находясь в возрасте от года до трех, детский организм отличается нестабильным составом крови. То есть время от времени происходит сдвиг лейкоцитарной формулы влево у детей или же вправо. При этом концентрация лимфоцитов и нейтрофилов может меняться в течение всего дня. Также причиной такой смены могут послужить определенные условия: переохлаждение; длительная прогулка на солнце; хронические заболевания; изменения на генном уровне. С 4 до 6 лет нейтрофилы выходят на лидирующую позицию. Однако у детей старше 6-7 лет состав крови идентичен параметрам взрослых. В течение всего этого периода гормональных изменений может наблюдаться сдвиг формулы на 10-15%, что является нормой.
Таблица 3
Лейкоцитарная формула у детей

Нейтрофилы (Нф) являются самой многочисленной группой. Основная их функция – фагоцитоз бактерий и продуктов распада тканей. Зрелые Нф содержат сегментированное ядро и три типа гранул. Гранулы содержат миелопероксидазу, катионные белки (КБ), кислые и щелочные фосфатазы, которые позволяют Нф переварить фагоцитированные объекты. Вне процессов фагоцитоза Нф секретируют нейтрофилокины: фактор некроза опухоли (ФНО), интерлейкины (ИЛ)-1, -6, -11. Нф первыми приходят в очаг повреждения благодаря хемоатрактантам и вазоактивным факторам. Нф оказывают цитотоксическое действие, а также продуцируют интерферон, обладающий противовирусным действием. Перед выходом в ткани Нф адгезируются к стенке сосудов при помощи адгезивных молекул – интегринов и селектинов. Бактерицидность Нф связана с респираторным взрывом. При контакте с агентом в Нф образуется супероксидные ионы и Н2О2, которые окисляют у поглощенных бактерий клеточную мембрану. Нф могут выбрасывать в окружающее пространство вещества, обладающие бактериальным действием (лактоферрин, лизоцим, КБ, гистоны).
Нф в инфицированных тканях способны формировать внеклеточные бактериоцидные ловушки, представленные выделяемыми нейтрофилами нитями ДНК.В 2004 году описан новый механизм антимикробного действия нейтрофилов – образование нейтрофильных внеклеточных ловушек (neutrophilextracellulartraps, NETs или НВЛ). Нейтрофильные гранулоциты после взаимодействия с микробными агентами (бактериями, простейшими, клетками грибов), а также с различными индукторами биологической или химической природы (опсонинами, цитокинами, активными формами кислорода при дыхательном взрыве, форбол12-миристат-13-ацетатом и т.д.) выбрасывают во внеклеточное пространство сетеподобные структуры – НВЛ, в состав которых входят ДНК, гистоны, а также различные белки и ферменты гранул. Было установлено, что образование НВЛ представляет собой один из базовых механизмов противоинфекционной защиты, биологическая функция которого не менее важна, чем фагоцитоз и секреция медиаторов нейтрофилами.
Эозинофилы при стыковке IgG- и IgE-опсонированных антигенов могут выделять из своих гранул цитотоксически действующие вещества и за счет этого повреждать покровы многочисленных паразитов. При инфекции организма личинками паразитов обнаруживается повышение эозинофилов в крови, которое в экстремальных случаях может составлять до 90% всех лейкоцитов. Эозинофилы обладают способностью к фагоцитозу, но это не имеет серьезного значения из-за их небольшого количества в крови, продуцируют фермент гистаминазу, который разрушает гистамин, освобождающийся из поврежденных базофилов и тучных клеток при различных аллергических состояниях.
При этом увеличивается количество эозинофилов в крови (эозинофилия). Эозинофилы продуцируют плазминоген, который является предшественником плазмина – главного фактора фибринолитической системы крови. Содержание эозинофилов в периферической крови подвержено суточным колебаниям, что связано с уровнем глюкокортикоидов. В конце второй половины дня и рано утром их на 20~ меньше среднесуточного уровня, а в полночь – на 30% больше.
Базофилы продуцируют и содержат биологически активные вещества (гепарин, гистамин и др.), чем и обусловлена их функция в организме. Гепарин препятствует свертыванию крови в очаге воспаления. Гистамин расширяет капилляры, что способствует рассасыванию и заживлению. В базофилах содержатся также гиалуроновая кислота, влияющая на проницаемость сосудистой стенки; фактор активации тромбоцитов (ФАТ); цитокины – факторы роста новых сосудов, активаторов кроветворения; тромбоксаны, способствующие агрегации тромбоцитов; лейкотриены и простагландины. Базофильные гранулоциты содержат также гранулы и во многих свойствах сходны с тучными клетками. Дегрануляция базофилов осуществляется после контакта IgE и антигена. Содержащийся в этих гранулах гистамин принимает участие в аллергических реакциях.
Некоторые микроорганизмы проявляют устойчивость к фагоцитозу или перевариванию в макрофагах. Например, возбудители туберкулеза, тифа, гонореи и проказы. Для борьбы с такими возбудителями необходимо активировать макрофаги с помощью цитокинов. Активация макрофагов приводит в том числе к синтезу цитотоксического пептида, так называемого дефензина, который может образовывать в мембране бактерии ионные каналы и таким образом уничтожать возбудителя. Активация макрофагов ведет также к экспрессии высокоактивной NO-синтазы, которая отщепляет от L-аргинина высоко активный NO. NO и сам обладает антимикробным действием, но его взаимодействие с О2-, приводит к образованию еще более активных соединений, таких как пероксинитрит (ONOO-), так что вместе с многочисленными бактериями могут быть успешно атакованы также грибы, простейшие и даже паразитные черви. Моноциты обладают выраженной фагоцитарной функцией. Это самые крупные клетки периферической крови и их называют макрофагами. Моноциты находятся в крови 2-3 дня, затем они выходят в окружающие ткани, где, достигнув зрелости, превращаются в тканевые макрофаги (гистиоциты). Моноциты способны фагоцитировать микробы в кислой среде, когда нейтрофилы не активны. Фагоцитируя микробы, погибшие лейкоциты, поврежденные клетки тканей, моноциты очищают место воспаления и подготавливают его для регенерации. Моноциты синтезируют отдельные компоненты системы комплемента. Активированные моноциты и тканевые макрофаги продуцируют цитотоксины, интерлейкин (ИЛ-1), фактор некроза опухолей (ФНО), интерферон, тем самым осуществляя противоопухолевый, противовирусный, противомикробный и противопаразитарный иммунитет; участвуют в регуляции гемопоэза. Макрофаги принимают участие в формировании специфического иммунного ответа организма. Они распознают антиген и переводят его в так называемую иммуногенную форму (презентация антигена). Моноциты продуцируют как факторы, усиливающие свертывание крови (тромбоксаны, тромбопластины), так и факторы, стимулирующие фибринолиз (активаторы плазминогена).
Клетки врожденных иммунных реакций принимают участие в процессах воспаления, поглощают и переваривают чужеродный материал. Проникающие микроорганизмы в жидких средах организма быстро захватываются фагоцитирующими клетками. К ним принадлежат нейтрофильные полиморфноядерные лейкоциты крови и встречающиеся в крови и тканях мононуклеарные фагоциты (моноциты, макрофаги). Если при ранении патогенные микробы проникли в ткани организма, то в первую очередь к месту повреждения привлекаются клетки неспецифической системы защиты. Это происходит за счет хемотаксиса, что означает направленное передвижение неспецифических воспалительных клеток, которое запускается и поддерживается за счет градиентов концентраций химических веществ. Хемотаксически активные вещества крайне многочисленны и здесь перечислена лишь их небольшая часть: некоторые из них продуцируются эндотелием поврежденных сосудов (простагландин, лейкотриен В4), часть тромбоцитами (PlateletActivatingFactor = PAF), некоторые входят в состав системы комплемента (белки C3 и C5). Кроме того, известны более чем 30 различных, так называемых хемокинов, которые привлекают определенные типы клеток.
Фагоцитоз начинается с захвата микроорганизмов и их связывания с мембранной поверхностью фагоцитов. Нагруженные C3b или антителами частицы (бактерии, поврежденные клетки организма) связываются с мембраной фагоцитов через C3b- или Fc-рецепторы (рис.3). После связывания фагоцит образует псевдоподии, которые окружают чужеродное тело (образование фагосомы). Непосредственное разрушение чужеродного тела происходит, когда фагосомы сливаются с лизосомами в фаголизосому, и ферменты лизосом вступают в контакт с фагоцитируемым материалом. Лизосомальные ферменты включают протеазы, пептидазы, оксидазы дезоксирибонуклеазы и липазы. Кроме того фагоциты (прежде всего нейтрофильные гранулоциты) продуцируют реактивные метаболиты кислорода, такие как перекись водорода (Н2О2), пероксид анионы (О2-) и гидроксилрадикалы (ОН.). Они повреждают мембраны бактерий и тем самым облегчают доступ лизосомальным ферментам.

Рис. 3.Фаза 1: Чужеродное тело, несущее антитела (например, IgG) или фактор системы комплемента C3b, распознается соответствующими рецепторами фагоцитов (Fc- и C3b-рецепторами) как нечто чужое. Фаза 2: После вступления в контакт с чужеродным организмом фагоциты образуют псевдоподии, которыми они «обхватывают» чужеродное тело. Фаза 3: После полного захвата чужеродного тела (фагоцитоз в собственном смысле) происходит образование фагосом. Фаза 4: Лизосомы, богатые гидролазой, сливаются с фагосомами и образуют фаголизосомы, в которых переваривается чужеродное тело. Фаза 5: Непереваренный материал выделяется наружу; на поверхности клетки появляются вновь Fc- и C3b-рецепторы, которые были расщеплены перед образованием фагосом (вторичная переработка)
Лимфоциты являются центральным звеном иммунной системы организма. Они осуществляют формирование специфического иммунитета, синтез защитных антител, лизис чужеродных клеток, реакцию отторжения трансплантата, обеспечивают иммунную память. Лимфоциты образуются в костном мозге, а дифференцировку проходят в тканях.
Выделяют отдельный класс лимфоцитов, формируются в результате дифференцировки лимфобластов (рис.4). NK-большие гранулярные лимфоциты, обладающие цитотоксичностью против опухолевых клеток и клеток, зараженных вирусами. Не имеют Т-клеточных рецепторов. Выполняют цитотоксические и цитокин-продуцирующие функции. Быстро активизируются, один из важнейших компонентов врожденного иммунитета. Главная функция – уничтожение клеток организма с дефектами или без молекул главного комплекса гистосовместимости I типа.
Лимфоциты, созревание которых происходит в вилочковой железе, называются Т-лимфоцитами (тимусзависимые). Тh0-лимфоциты («наивные» недифференцированные Т-лимфоциты-хелперы), которые содержат молекулы CD-4 или Т-клеточный рецептор (TCR) дифференцируются в Th1 (Т-хелперы 1) и Th2 (Т-хелперы 2) соответственно. Если образуются Тh1-лимфоциты, активизируется клеточный ответ, направленный на уничтожение вирусных агентов, опухолевых клеток, паразитов. Th1-клетки продуцируют интерлейкин-2 (IL-2) - цитокин, выполняющий функцию фактора роста, в результате происходит размножение Т-эффекторов, которые могут быть двух видов: ТЦТ (цитотоксические лимфоциты) и ТГЗТ (лимфоциты гиперчувствительности замедленного типа). Также Th1-клетки продуцируют интерферон γ - эффекторный цитокин, обладающий прямой противовирусной и противоопухолевой активностью. Если образуются Th2-лимфоциты, активизируется гуморальный ответ, направленный против растворимых и клеточных антигенов. Th-лимфоциты, которые преобразуются в Th2-лимфоциты, взаимодействуют с рецепторами В-лимфоцитов, которые являются встроенными в мембрану иммуноглобулинами (IgM-мономер, IgD). В результате взаимодействия происходит передача антигенной детерминанты от Th2 к B-клетке и продукция Th2 клетками ростовых факторов ИЛ-4,5,6. Под действием этих факторов антиген-специфические B-лимфоциты начинают размножаться и дифференцироваться в плазматические клетки, которые продуцируют Ig (антитела). Антитела связываются с растворимыми антигенами, образуют иммунные комплексы, элиминируемые, в последствии, из организма. Второй вариант эффекторной фазы гуморального иммунного ответа может быть направлен на вирусинфицированные или опухолевые клетки.
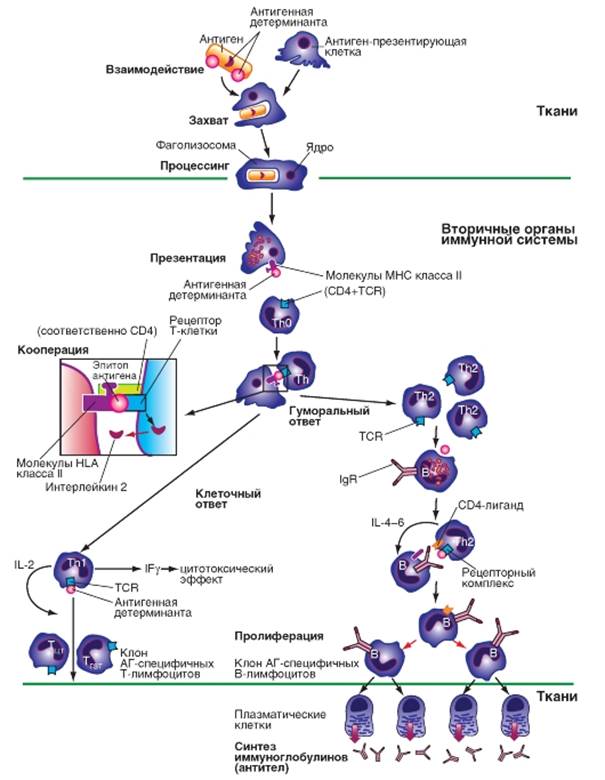
Рис.4. Дифференцировка лимфоцитов
В этом случае антитела связывается с антигеном на поверхности клетки; происходит активация комплемента и нарушение целостности цитоплазматической мембраны. Различают также Т–киллеры (убийцы) осуществляют реакции клеточного иммунитета, лизируя чужеродные клетки, возбудителей инфекционных заболеваний, опухолевые клетки, клетки-мутанты. Т-супрессоры (угнетатели) блокируют чрезмерные реакции В-лимфоцитов. Имеются также Т-хелперы и Т-супрессоры, регулирующие клеточный иммунитет. Т-клетки памяти хранят информацию о ранее действующих антигенах.
В-лимфоциты (бурсозависимые) проходят дифференцировку у человека в лимфоидной ткани кишечника, небных и глоточных миндалин. В-лимфоциты осуществляют реакции гуморального иммунитета. Большинство В-лимфоцитов являются антителопродуцентами. В-лимфоциты в ответ на действие антигенов в результате сложных взаимодействий с Т-лимфоцитами и моноцитами превращаются в плазматические клетки. Плазматические клетки вырабатывают антитела, которые распознают и специфически связывают соответствующие антигены. В абсолютном большинстве случаев для «узнавания» антигена В-клетками и для их превращения в плазматические клетки, выделяющие антитела, необходимы еще антиген-презентирующие клетки и Т-хелпера.
Различают 5 основных классов антител, или иммуноглобулинов: JgA, JgG, JgМ, JgD, JgЕ.
JgМ- это наиболее «ранние» из всех классов Ig активируют систему комплемента.
JgЕ - это мономеры, содержание которых в сыворотке крови ничтожно мало. Связывается со специфическими рецепторами на поверхности тучных клеток и базофилов с высвобождением из этих клеток медиаторов аллергии.
JgA - это секреторные иммуноглобулины, секретируется в различные жидкости организма, обеспечивая секреторный иммунитет.
JgD - это мономеры, функционирует в основном в качестве мембранных рецепторов для антигена.
JgG - это мономеры, которые отличаются друг от друга по аминокислотному составу и антигенным свойствам. IgG проявляет разнообразные виды активности, в том числе способность проникать через плаценту.
Наряду с плазматическими клетками при контакте с антигеном возникают В-клетки памяти, которые после контакта с антигеном не выделяют иммуноглобулины, а сохраняют информацию о структуре антигена. При последующем контакте с антигеном они под влиянием Т-хелперов и Т-клеток памяти, могут незамедлительно продуцировать большие количества антител. Эта «функция памяти» иммунной системы не столько связана со специфическими клетками памяти, сколько является результатом постоянного и повторяющегося контакта малейших количеств антигена с субпопуляцией В- и Т-клеток, которая держит антиген в «поле зрения», чтобы не забыть его.
О-лимфоциты (нулевые) не проходят дифференцировку и являются как бы резервом Т- и В-лимфоцитов.
Регуляция лейкопоэза.
Все лейкоциты образуются в красном костном мозге из единой стволовой клетки (рис.5), однако родоначальницей миелопоэза является бипотенциальная колониеобразующая единица гранулоцитарно-моноцитарная (КОЕ-ГМ) или клетка-предшественница. Для ее роста и дифференцировки необходим особый колониестимулирующий фактор (КСФ), вырабатываемый у человека моноцитарно-макрофагальными клетками, костным мозгом и лимфоцитами. КСФ является гликопротеидом и состоит из двух частей — стимулятора продукции эозинофилов (Эо-КСФ) и стимулятора продукции нейтрофилов и моноцитов (ГМ-КСФ), относящихся к ранним гемопоэтическим ростовым факторам. Содержание ГМ-КСФ стимулируется Т-хелперами и подавляется Т-супрессорами. На более поздних этапах на лейкопоэз влияют гранулоцитарный колониестимулирующий фактор — Г-КСФ (способствует развитию нейтрофилов) и макрофагальный колониестимулирующий фактор — М-КСФ (приводит к образованию моноцитов), являющиеся поздно действующими специфическими ростовыми факторами. Из костного мозга и отдельных видов лейкоцитов (гранулоцитов и агранулоцитов) выделен комплекс полипептидных факторов, выполняющих функции специфических лейкопоэтинов.
Важная роль в регуляции лейкопоэза отводится интерлейкинам. В частности, ИЛ-3 не только стимулирует гемопоэз, но и является фактором роста и развития базофилов. ИЛ-5 необходим для роста и развития эозинофилов. Многие интерлейкины (ИЛ-2, ИЛ-4, ИЛ-6, ИЛ-7 и др.) являются факторами роста и дифференцировки Т- и В-лимфоцитов.
Ингибиторы миелопоэза - лактоферрин, содержащийся в мембране макрофагов, кислый изоферритин, гранулоцитарные кейлоны. Угнетают гранулоцитопоэзкейлоны, выделяющиеся зрелыми нейтрофилами. Кейлоны тормозят синтез ДНК в стволовых клетках белого ростка костного мозга. Задерживают созревание гранулоцитов и моноцитов простагландины Е, интерфероны. Лимфоцитарные кейлоны - тканевоспецифические ингибиторы клеточного деления. Лимфоцитарные кейлоны представляют собой гликопротеиды, они вырабатываются в тимусе, селезенке, лимфобластами. Иммунодепрессивное действие кейлонов связано с подавлением синтеза ДНК и пролиферации лимфоцитарных клеток. Процессы дифференцировки лимфоцитов регулируют лимфопоэтины.
Лейкоциты являются наиболее «подвижной» частью крови, быстро реагирующей на различные изменения в окружающей среде и организме развитием лейкоцитоза, что обеспечивается существованием клеточного резерва. Известны два типа гранулоцитарных резервов — сосудистый и костномозговой. Сосудистый гранулоцитарныи резерв представляет собой большое количество гранулоцитов, расположенных вдоль стенок сосудистого русла, откуда они мобилизуются при повышении тонуса симпатического отдела автономной (вегетативной) нервной системы. Количество клеток костномозгового гранулоцитарного резерва в 30—50 раз превышает их количество в кровотоке. Мобилизация этого резерва происходит при инфекционных заболеваниях, сопровождается сдвигом лейкоцитарной формулы влево и обусловлена в основном воздействием эндотоксинов. Своеобразные изменения претерпевают лейкоциты в разные стадии адаптационного синдрома, что обусловлено действием гормонов гипофиза (АКТГ) и надпочечника (адреналина, кортизона, дезоксигидрокортизона). Уже через несколько часов после стрессорного воздействия развивается лейкоцитоз, который обусловлен выбросом нейтрофилов, моноцитов и лимфоцитов из депо крови. При этом число лейкоцитов не превышает 16—18 тыс. в 1 мкл. В стадии резистентности число и состав лейкоцитов мало отличаются от нормы.В стадии истощения развивается лейкоцитоз, сопровождающийся увеличением числа нейтрофилов и снижением числа лимфоцитов и эозинофилов.

Рис.5. Регуляция лейкопоэза
Эстрогены активируют клетки иммунной системы, повышают устойчивость организма женщин к инфекции.Соматотропный гормон тормозит созревание Нф. Андрогены стимулируют гранулоцитопоэз. Адреналин вызывает перераспределительный/физиологический лейкоцитоз. Лейкопоэз стимулируют продукты распада самих лейкоцитов и тканей, микроорганизмы и их токсины, некоторые гормоны гипофиза, нуклеиновые кислоты,
Лейкоциты разрушаются в слизистой оболочке пищеварительного тракта, а также в ретикулярной ткани.






