Физиологическая роль цитокининов. — КиберПедия
Биохимия спиртового брожения: Основу технологии получения пива составляет спиртовое брожение, - при котором сахар превращается...
Поперечные профили набережных и береговой полосы: На городских территориях берегоукрепление проектируют с учетом технических и экономических требований, но особое значение придают эстетическим...
Установка замедленного коксования: Чем выше температура и ниже давление, тем место разрыва углеродной цепи всё больше смещается к её концу и значительно возрастает...
2.3. Физиологическая роль цитокининов. …………………….……... 10
а) Стимуляция деления клеток. …………………………………. 10
б) Влияние цитокининов на рост клеток. …………………….... 11
в) Действие цитокининов на органогенез. …………………...… 12
г)Прерывание покоя и стимуляция прорастания семян под
действием цитокининов. …………………………………...…… 14
д) Влияние цитокининов на рост целых растений. ………….… 14
е) Защитное действие цитокининов при неблагоприятных
факторах среды. ………………..…………………………..…… 15
2.4. Механизм действия цитокининов. ……………………………… 18
3. Влияние фактора засоления на растительные организмы. ………… 20
3.1. Типы засоления почв. ……………………………………………. 20
3.2. Причины и последствия влияния засоления на растительные
организмы. ……………………………………………………………. 21
3.3. Механизмы адаптации к засолению. …………………………… 27
3.4. Растения засолённых почв – галофиты. ………………………... 32
3.5. Метод борьбы с засолением почвы и повышения солеустойчивости
растений. ……………………………………………………………….. 33
II. ЭКСПЕРИМЕНТАЛЬНАЯ ЧАСТЬ
1. Объект и методы и исследований. …………………………………….. 34
1.1. Характеристика объекта исследований. …………………………. 34
а) Особенности морфологии; …………………………………….. 34
б) Особенности роста и развития; ……………………………… 35
в) Особенности биологии; ………………………………………… 36
г) Народохозяйственное значение. ……………………………….. 37
1.2. Методы исследований. …………………………………………….. 39
2. Результаты исследований и их обсуждение. ………………………….. 41
2.1. Влияние засоления на растения кукурузы. ………………………. 41
а) Влияние уровня засоления на высоту растений; …………….. 41
б)Влияние засоления на содержание воды и водоудерживающую
способность; ………………………………………………………. 42
в) Влияние засоления на интенсивность транспирации; ………. 45
г) Влияние засоления на анатомо-морфологическую структуру. 47
2.2. Влияние 6-БАП на растения кукурузы в зависимости от уровня
засоления. ………………………………………………………………... 50
а) Влияние 6-БАП на высоту растений в зависимости от уровня
засоления; ……………………………………………………………. 50
б) Влияние 6-БАП на содержание воды и водоудерживающую
способность в зависимости от уровня засоления; ……………….. 53
в) Влияние 6-БАП на интенсивность транспиации в зависимости от
уровня засоления; ……………………………………………………. 54
г) Влияние 6-БАП на анатомо-морфологическую структуру в
зависимости от уровня засоления. ………………………………… 56
ВЫВОДЫ ………………………………………………………………………... 59
СПИСОК ЛИТЕРАТУРЫ ………………………………………………………. 60
ПРИЛОЖЕНИЕ ………………………………………………………………….. 65
ВВЕДЕНИЕ
Одной из самых острых проблем современности является деградация почв - главного фундамента жизни. В нашей стране засолено около 10% поверхности суши. Причем в настоящее время увеличиваются масштабы вторичного засоления почв, которое развивается чаще всего при нерациональном орошении. Таким образом, засоление начинает признаваться как важный фактор, лимитирующий продуктивность сельскохозяйственных культур, который оказывает глубокое воздействие на все стороны жизнедеятельности растений. При этом изменяются как структура, так и функции растений. Известно, что длительное повышенное содержание ионов в почве оказывает значительное влияние на многие физиологические процессы растений, вызывая при этом и анатомические изменения [43,46].
С другой стороны, в настоящее время наблюдается увеличение аридности и ксерофильности климата, установилось новое соотношение СО2 и О2 в атмосфере. По последним оценкам состояния геосистем Северной Евразии будет увеличиваться количество засух. В связи с такими глобальными изменениями климата особое внимание обращается на виды растений с С4 фотосинтезом. Прогнозируется, что общее потепление будет способствовать смещению в более северные области границ районирования таких хозяйственно ценных С4 растений, как кукуруза, сорго, амарант, некоторые формы проса [17].
Таким образом, С4 растения представляют определенный интерес с точки зрения изучение путей снижения отрицательного действия засоления на растительный организм, одним из которых является применение фитогормонов, относящихся к группе цитокининов, которые принимают участие в антистрессовых реакциях растений и повышают их адаптивные возможности.
В связи с этим, целью данной работы являлось изучение влияния обработки синтетическим аналогом цитокинина на физиологические и анатомо-морфологические показатели растений кукурузы, выращенных при разном уровне засоления субстрата.
I. ЛИТЕРАТУРНЫЙ ОБЗОР.
1. КРАТКАЯ ХАРАКТЕРИСТИКА ФИТОГОРМОНОВ.
Важнейшими представителями эндогенных регуляторов роста растений являются фитогормоны. Это вещества, которые синтезируются в растениях, транспортируются по ним в малых концентрациях и способны вызывать ростовые или формативные эффекты [10].
Все фитогормоны обладают тремя основными особенностями:
1. Эндогенное происхождение. Изменения в интенсивности синтеза того или иного фитогормона, вызванное внутренними или внешними причинами, вызывает ответную реакцию растения — переход к другому характеру ростовых или формативных процессов.
2. Возможность транспортировки по растению. Физиологический смысл этой особенности состоите том, что фитогормон, образовавшийся в одном органе (например, в апикальной меристеме стебля), должен обладать свойством регуляции ростовых процессов в других органах (например, в корне). Именно таким образом достигается взаимодействие органов и целостность растения
3. Способность в малых концентрациях (10-12 - 10-7М) вызывать заметные ростовые или формативные эффекты. Примером ростового эффекта может служить ускорение или замедление роста стебля, формативного — дефолиация.
Работы многих ученых [3, 7, 9, 21, 30, 33] показали, что фитогормоны участвуют в регуляции обмена веществ на всех этапах жизни растений — от развития зародыша до полного завершения жизненного цикла и отмирания. Они определяют характер роста и развития растений, формирования новых органов, габитуса, цветения, старения вегетативных частей, перехода к покою и выхода из него и т.п.
В настоящее время известно восемь групп фитогормонов: пять из которых относятся к классическим группам - ауксины, гиббереллины, цитокинины, абсцизовая кислота, этилен и три открытые сравнительно недавно - брассиностероиды, жасминовая и салициловая кислоты. Все они могут активизировать, но могут и тормозить функциональную активность клеток. Общим условием для действия любого фитогормона является наличие в клетках специфичных рецепторов. Все фитогормоны вызывают у компетентных клеток сравнительно быстрые физиологические реакции, связанные, очевидно, с мембранами и более медленные изменения, зависящие от синтеза белков и нуклеиновых кислот [33].
Предполагается наличие в растениях еще неизвестных классов гормонов, о свойствах которых можно судить только по косвенным показателям. Это — факторы цветения (флориген, ВЕНВ), ризогенеза и роста клеток (фузикокцин), повышения устойчивости растений (олигосахариды). Все это свидетельствует о том, что гормональная система растений значительно сложнее, чем представлялось до недавнего времени.
Помимо естественных фитогормонов, получено большое количество синтетических аналогов этих природных соединений, которые часто обладают высокой физиологической активностью. Строго говоря, эти вещества не могут быть отнесены к фитогормонам, так как не образуются в растениях, однако многие из них по активности не уступают фитогормонам или даже превосходят их [32].
2. ОБЩАЯ ХАРАКТЕРИСТИКА ЦИТОКИНИНОВ.
Химическая структура.
Цитокининами называется один из типов фитогормонов, обладающих определенной совокупностью биологической активности, которая весьма разнообразна и проявляется при регуляции роста, органообразования, процессов старения и покоя.
Цитокинины принимают участие наряду с другими фитогормонами в регуляции самых разнообразных физиологических процессов в растении. Для них, как и для других фитогормонов, характерна полифункциональность. Кроме того, действие цитокининов появляется в неразрывной связи с действием других фитогормонов и природных ингибиторов.
В химическом отношении природные цитокинины и их синтетические заменители представляют собой производные 6-аминопурина с заместителем в аминогруппе при шестом атоме углерода пуринового кольца.
Пурин
Основными представителями цитокининового ряда являются: кинетин, 6-бензиламинопурин (6-БАП), 8-азакинетин, бензимидазол, которые принадлежат к синтетическим цитокининам, а представителем природных цитокининов является зеатин, который был выделен из растения кукурузы [32, 48].
Цитокинины пуринового ряда слаборастворимы в воде, но хорошо растворимы в этаноле, этиловом эфире, ряде щелочей и кислот. Цитокинины устойчивы к нагреванию, автоклавированию, действию щелочей и кислот. Образование природных цитокининов происходит в корневой системе, а передвижение в надземные органы идёт по ксилеме [49].
История открытия.
Цитокинины были открыты в 1955 г. Скугом и Миллером с сотрудниками в Висконсинском университете в США. Их обнаружению помог так называемый “дефектный объект”, которым явился каллюс сердцевины стебля табака. Он образовался на кусочках сердцевины стебля в условиях стерильной культуры, но быстро прекращал рост в связи с исчерпанием какого-то фактора, исходно присутствовавшего в тканях стебля. Ввиду того, что вернуть активный рост каллюса удавалось добавлением к питательной смеси, содержащей ИУК, дрожжевого экстракта, а так же кокосового молока, Скугом и сотрудниками была предпринята попытка выделить недостающие для роста каллюса вещество из дрожжевого экстракта. При этом выяснилось, что активное вещество обладает свойствами пурина.
Вместе с тем, проверка природных пуриновых оснований, так же как гидрометода РНК и ДНК показала, что они не способны обеспечить рост стеблевого каллюса табака. Неожиданно активным оказался эфирный экстракт из препарата ДНК сельди. Впоследствии выяснилось, что активное вещество образуется в любом препарате ДНК при ее деградации путем автоклавирования в кислой среде. Это вещество было выделено в виде кристаллов и идентифицировано химически. Оно оказалось 6- фурфуриламинопурином, который был назван кинетином.
6-фурфуриламинопурин
Принцип химического синтеза производных пурина с замещением в аминогруппе у шестого атома углерода пуринового кольца был известен ранее, поэтому вслед за химической идентификацией кинетина последовал его синтез. Затем было синтезировано много других активных соединений, которые отличались от кинетина характером заместителя в аминогруппе при шестом атоме углерода. Все эти соединения, включая кинетин, были объединены под общим названием кинины. Это название было распространено в литературе до 1965 г. Однако ввиду того, что в физиологии животных термин “кинины” еще ранее был применен для совершенно иных соединений - биологически активных полипептидов, Скуг, Стронг и Миллер предложили заменить название кинины на цитокинины. В настоящее время термин цитокинины стал общепризнанным. [22].
Типы засоления почв.
Согласно Б. П. Строгонову [38], по степени засоления различают практически незасоленные, слабозасоленные, среднезасоленные почвы и солончаки. Тип засоления определяется по содержанию анионов в почве: хлоридное, сульфатное, сульфатно-хлоридное, хлоридно-сульфатные и карбонатное. Преобладающим катионом в таких почвах является натрий (поваренная соль (NaCl), сода (Na2CO3), глауберова соль (Na2SO4), но встречаются также карбонатно-магниевое (кальциевое) и хлоридно-магниевое (кальциевое) засоление [31]. Наиболее вредное влияние оказывает содовое засоление, поскольку в почве сода распадается, образуя сильную щелочь (гидроксид натрия). Все эти соли хорошо растворимы в воде, так что во влажном климате обычно вымываются из почвы атмосферными осадками и сохраняются в ней в ничтожных количествах. В сухом же и жарком климате не только не происходит промывания почвы дождем, но, наоборот, растворы солей поднимаются с восходящим током почвенной воды из глубин субстрата. Вода испаряется, а соли остаются в верхних слоях почвы. Накапливаясь, они вызывают образование солончаков и солонцов. Неумелое искусственное орошение в пустынной зоне вызывает засоление почвы. Так, в поливной зоне нашей страны насчитывается до 36% засоленных земель. По побережьям морей даже при влажном климате почва насыщена солями.
Солончаки весной увлажнены грунтовыми водами, их называют мокрыми. Концентрация солей в почвенном растворе достигает нескольких десятков процентов, причем наибольшая засоленность почвы наблюдается в сухие периоды года, когда солончаки высыхают. В это время поверхность почвы покрывается налетом соли, сверкающей на солнце. Солончаки есть и в степных зонах России, но главным образом распространены в пустынной зоне. Они встречаются как мелкими пятнами, так и большими массивами.
Солонцы отличаются от солончаков: поверхностные слои их почвы не засолены, а соли, вредные для растений, накапливаются в более глубоких горизонтах. Поверхностный слой солонца бесструктурный, сильно выщелоченный; ниже его расположен уплотненный, слой почвы, насыщенный натриевыми солями. В сухое время года эти слои растрескиваются на глыбы, столбцы. Ниже этих слоев располагается почва, засоленная растворимыми солями.
Весной плотные слои солонца долго задерживают воду на его поверхности. Летом же, когда уплотненные слои высыхают и растрескиваются, выпадающие атмосферные осадки по трещинам устремляются в глубь субстрата, не увлажняя поверхности [28].
Методы исследований.
Исследования проводились на базе лаборатории физиологии растений Московского Государственного Областного Университета в период с июня по сентябрь 2003 года. Объектом изучения являлись растения кукурузы сорта Россо. Изучение влияния 6-БАП на рост и биометрические показатели растений кукурузы при разном уровне засоления проводилось в условиях вегетационного опыта (почвенные культуры).
Семена кукурузы проращивались при температуре 20оС в термостате в течение 3 дней, а затем высаживались в сосуды Митчерлиха с почвой на 5 кг. Почва дерново-подзолистая средне-суглинистая. В каждом сосуде в среднем выращивалось по 7 растений.
Эксперимент включал 6 вариантов, разный уровень засоления создавался путём внесения раствора NaCl.
1. Контроль;
2. Обработка 6-БАП;
3. 0,1% NaCl;
4. 0,1% NaCl+6-БАП;
5. 0,2% NaCl;
6. 0,2% NaCl+6-БАП.
Опрыскивание синтетическим аналогом цитокинина 6-БАП концентрацией 4х10-5М (20мг/л) проводили в фазу кущения растений 10 июля 2003 г. В вариантах без обработки растения опрыскивались водой.
Учет роста проводился каждые 7 дней. Содержание воды, интенсивность транспирации и водоудерживающая способности фиксировались 4 раза.
Уход за растениями осуществлялся в соответствии с общепринятой агротехникой.
В ходе исследований были определены следующие показатели:
1. Высота растений;
2. Содержание воды;
3. Водоудерживающая способность;
4. Интенсивность транспирации;
5. Анатомо-морфологическая структура листа и стебля кукурузы.
Высота растений фиксировалась при помощи измерительной ленты. Содержание воды определялось по разнице между сырой и сухой массой растения, измеренной весовым методом. Анатомо-морфологическую структуру листа и стебля кукурузы анализировали под микроскопом МБР-1 с помощью окуляра-микрометра.
Водоудерживающая способность рассчитывалась по формуле:
(P1 – P3) x 100
Водоудерживающая способность = 100 - [ % ], где
Р3
Р1 – сырая масса листа;
Р3 – масса листа через 20 минут после удаления.
Интенсивность транспирации определялась методом, основанным на учёте потери воды листом за короткий промежуток времени.
Высота растений, см
17.07
%
24.07
%
31.07
%
07.08
%
14.08
%
Контроль
12,1
+ 0,7
100
15,4
+ 0,4
100
19,3
+ 0,8
100
24,1
+ 0,5
100
29,7
+ 0,73
100
0,1%
11,2
+ 0,5
92,6
14,1
+ 0,7
91,6
17,6
+ 0,57
91,2
22,3
+ 0,6
92,5
27,2
+ 0,43
91,6
0,2%
10,1
+ 0,66
83,5
11,9
+ 0,6
77,3
15,3
+ 0,68
79,3
19,4
+ 0,43
80,5
24,2
+ 0,5
81,5
Из данных таблицы следует, что снижение высоты растений находится в прямой зависимости от концентрации соли в субстрате. Схожие данные были получены в работах Йоневой, Калининой, Федяевой, Атанасовой [12, 14, 43, 51]. Причем, при увеличении концентрации соли на 0,1%, высота растений уменьшается в среднем на 8-9%. Угнетение роста наблюдается на протяжении всего опыта, что подтверждается данными графика 1, где по оси абсцисс отложены даты измерений, а по оси ординат – высота растений в сантиметрах. Контрольные растения обозначены голубым цветом, выращенные при 0,1% засолении – оранжевым; при 0,2% – зеленым.
Рисунок 1. Влияние уровня засоления на рост растений кукурузы.
б) Влияние засоления на содержание воды и водоудерживающую способность.
Как известно, увеличение концентрации соли в субстрате сказывается не только на ростовых процессах растения, но и на водном обмене. Для изучения этого вопроса определялось процентное содержание воды в листьях и водоудерживающая способность. Проведенные измерения показали, что по мере повышения уровня засоления, содержание воды в листьях растения снижается. Так, если рассмотреть эти данные в процентном соотношении на 31.07, то видно, что оводненность растений, выращенных при 0,1% засолении, на 3,9% ниже контрольных, а при 0,2% - на 6,4%.
Таблица 2. Влияние уровня засоления на оводненность листьев кукурузы.
Дата
Вариант
Содержание воды, %
17.07
%
24.07
%
31.07
%
07.08
%
Контроль
88,8
+ 4,0
100
88,8
+ 3,1
100
87,3
+ 1,6
100
78,8
+ 1,9
100
0,1%
87,5
+ 3,7
98,5
86,7
+ 3,7
97,6
83,9
+ 2,5
96,1
77,7
+ 2,2
98,6
0,2%
86,6
+ 3,5
97,5
85,9
+ 3,9
96,7
81,7
+ 3,1
93,6
73,3
+ 3,4
93,0
Снижение содержания воды в листьях растений кукурузы при 0,1% засолении составило от 1,1 до 3,4%, что подтверждается данными рисунка 2, где по оси абсцисс отложены даты измерений, а по оси ординат – содержание воды в листьях в %.
Рисунок 2. Влияние уровня засоления на содержание воды в листьях растений кукурузы.
Для растений, выращенных на почве с 0,2% засолением NaCl, эти цифры составили от 2,2 до 5,6%. В среднем оводненность растений, выращенных при 0,1 % засолении, на 2,3% ниже контрольных, а при 0,2 М - на 4,8%.
Сходная тенденция наблюдается и в опытах по определению водоудерживающей способности (табл.3), но засоление оказывает меньшее влияние на этот показатель.
Таблица 3. Влияние уровня засоления на водоудерживающую способность листьев кукурузы.
Дата
Вариант
Водоудерживающая способность, %
17.07
%
24.07
%
31.07
%
07.08
%
Контроль
97,5
+ 4,5
100
96,8
+ 3,9
100
97,4
+ 2,6
100
96,2
+ 3,1
100
0,1%
96,5
+ 4,2
99,0
96,5
+ 3,8
99,7
97,3
+ 2,6
99,9
95,1
+ 3,4
98,9
0,2%
94,8
+ 4,7
97,2
96,3
+ 3,8
99,5
95,8
+ 3,0
98,4
94,8
+ 3,4
98,5
Так при концентрации соли 0,1%, водоудерживающая способность уменьшилась от 0,1 до 1,1% по сравнению с контролем, а при 0,2% - от 0,5 до 2,7%, что можно видеть на рисунке 3. В среднем водоудерживающая способность понизилась на 0,6% при 0,1% концентрации NaCl; и на 1,6% при 0,2% засолении.
Полученные данные находят отражение в литературе [17]. В целом, можно сказать, что засоление приводит к подавлению способности к осморегуляции, то есть с увеличением концентрации соли растения теряют способность сохранять оводненность органов и это отрицательно сказывается на их солеустойчивости.
Рисунок 3. Влияние уровня засоления на водоудерживающую способность листьев кукурузы.
в) Влияние засоления на интенсивность транспирации.
Содержание воды и водоудерживающая способность – это не единственные показатели водного обмена растений. Более важным критерием является интенсивность транспирации. В таблице 4 и на рисунке 4 приведены данные по влиянию уровня засоления на этот показатель.
Проведенные измерения показали, что интенсивность транспирации растений, выращенных на фоне засоления, снижается с повышением концентрации соли в субстрате. Так, если рассмотреть результаты полученные на 17.07, видно, что интенсивность транспирации у растений, выращенных при концентрации соли 0,2 %, в 2 раза ниже, чем у контрольных; при 0,1 % – на 43,5%. Полученные данные можно объяснить тем, что в условиях засоления растения испытывают недостаток воды, что вызывает накопление АБК в листьях, приводящее к закрытию устьиц и, как следствию, снижению уровня транспирации. Схожие данные были получены Ахияровой Г.Р. и Веселовым Д.С. [3].
Таблица 4. Влияние уровня засоления на интенсивность транспирации листьев кукурузы.
Дата
Вариант
Высота растений, см
17.07
%
24.07
%
31.07
%
07.08
%
14.08
%
Контроль
12,1
+ 0,7
100
15,4
+ 0,4
100
19,3
+ 0,8
100
24,1
+ 0,5
100
29,7
+ 0,73
100
Опрыс-кивание 6-БАП
14,0
+ 0,4
115
18,5
+ 0,57
120
22,6
+ 0,68
117
27,5
+ 0,4
114
33,3
+ 0,66
112
0,1%
11,2
+ 0,5
100
14,1
+ 0,7
100
17,6
+ 0,57
100
22,3
+ 0,6
100
27,2
+ 0,43
100
0,1%
+6-БАП
13,2
+ 0,43
118
17,5
+ 0,5
124
20,8
+ 0,8
118
26,1
+ 0,7
117
31,0
+ 0,5
114
0,2%
10,1
+ 0,66
100
11,9
+ 0,6
100
15,3
+ 0,68
100
19,4
+ 0,43
100
24,2
+ 0,5
100
0,2%
+ 6-БАП
12,4
+ 0,5
123
15,5
+ 0,43
130
19,0
+ 0,5
124
22,5
+ 0,66
116
27,6
+ 0,7
114
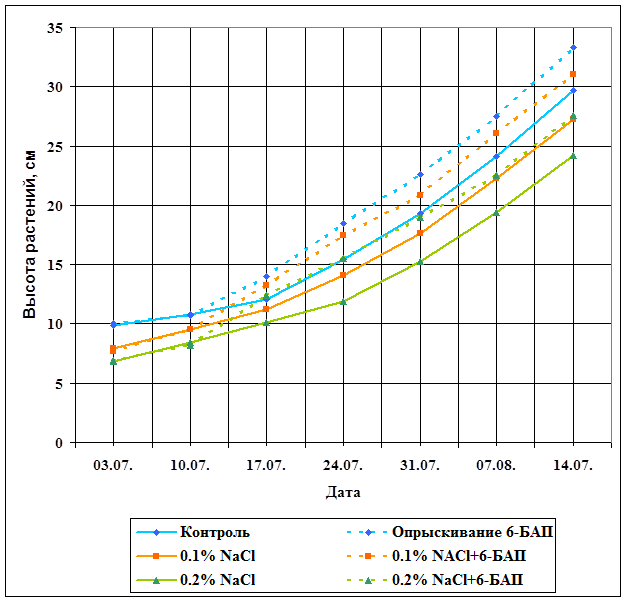
Значительная прибавка в росте наблюдается в первые 3 недели после обработки синтетическим аналогом цитокинина, и достигает своего максимума на второй неделе после опрыскивания. Затем разница в высоте растений, опрыснутых 6-БАП и не прошедших обработку, начинает становиться менее заметной.
На рис.5 представлен график ростовых процессов опытных растений. Как видно, максимальной величиной характеризуются растения, выращенные на незасоленной почве и обработанные 6-БАП. Опрыскивание синтетическим аналогом цитокинина в некоторой степени снимает угнетающие действие солей на рост растений, о чем может свидетельствовать тот факт, что растения, выращенные при 0,1% концентрации NaCl и обработанные 6-БАП, оказались в среднем на 12% выше контрольных. Вышесказанное свидетельствует о том, что 6-БАП способствует нормализации ростовых процессов, что говорит о повышении адаптивных возможностей растительного организма. Полученные данные находят отражение в литературе [14, 51].
Рисунок 5. Влияние обработки 6-БАП на рост растений кукурузы в зависимости от уровня засоления.
б) Влияние 6-БАП на содержание воды и водоудерживающую способность в зависимости от уровня засоления.
Тенденция по увеличению действия 6-БАП с возрастанием засолённости почвы прослеживалась и в опытах по определению оводнённости листьев кукурузы. Данные представлены в таблице 7.
Таблица 7. Влияние 6-БАП на содержание воды в листьях кукурузы в зависимости от уровня засоления.
Дата
Вариант
Содержание воды, %
17.07
%
24.07
%
31.07
%
07.08
%
Контроль
88,8
+ 4,0
100
88,8
+ 3,1
100
87,3
+ 1,6
100
78,8
+ 1,9
100
Опрыс-кивание 6-БАП
89,6
+ 4,2
100,9
89,4
+ 3,2
100,7
88,9
+ 2,0
101,8
80,4
+ 2,3
102
0,1%
87,5
+ 3,7
100
86,7
+ 3,7
100
83,9
+ 2,5
100
77,7
+ 2,2
100
0,1%
+6-БАП
88,7
+ 4,1
101,4
88,9
+ 4,3
102,5
87,1
+ 3,4
103,8
78,3
+ 2,4
100,8
0,2%
86,6
+ 3,5
100
85,9
+ 3,9
100
81,7
+ 3,1
100
73,3
+ 3,4
100
0,2%
+ 6-БАП
86,9
+ 3,5
100,3
85,9
+ 3,9
102,9
86,5
+ 2,7
105,8
76,0
+ 4,2
103,7
Обработка синтетическим аналогом цитокинина вызвала повышение содержания воды по сравнению с контролем. Причём у контрольных растений она составила в среднем 1,1%, выращенных на субстрате с 0,1% засолением – 2,1%; при 0,2% засолении – 3,1%.
Что касается показателей водоудерживаюшей способности, то их увеличение под действием 6-БАП оказалось незначительным и составило менее 1% (табл.8).
Таблица 8. Влияние 6-БАП на водоудерживающую способность листьев кукурузы в зависимости от уровня засоления.
Дата
Вариант
Водоудерживающая способность, %
17.07
%
24.07
%
31.07
%
07.08
%
Контроль
97,5
+ 4,5
100
96,8
+ 3,9
100
97,4
+ 2,6
100
96,2
+ 3,1
100
Опрыс-кивание 6-БАП
98,3
+ 4,7
100,8
97,0
+ 3,9
100,2
98,0
+ 2,7
100,6
96,6
+ 3,0
100,4
0,1%
96,5
+ 4,2
100
96,5
+ 3,8
100
97,3
+ 2,6
100
95,1
+ 3,4
100
0,1%
+6-БАП
97,2
+ 4,3
100,7
96,8
+ 3,9
100,3
97,6
+ 2,7
100,3
95,5
+ 3,5
100,4
0,2%
94,8
+ 4,7
100
96,3
+ 3,8
100
95,8
+ 3,0
100
94,8
+ 3,4
100
0,2%
+ 6-БАП
96
+ 4,4
101,3
96,3
+ 3,9
100
97,6
+ 3,5
101,9
95,3
+ 3,3
100,5
в) Влияние 6-БАП на интенсивность транспиации в зависимости от уровня засоления.
Данные представлены в таблице 9 и на рисунке 6.
Повышение содержания воды в листьях кукурузы после обработки 6-БАП коррелирует с уменьшением интенсивности транспирации. Так для контрольных растений, обработанных 6-БАП, снижение транспирации составило от 22 до 55,8%, по сравнению с необработанными. Для растений, произраставших при 0,1% засолении, эти цифры составляют 17,2-57,6%; при 0,2% - от 33,8 до 62%.
Таблица 9. Влияние 6-БАП на интенсивность транспирации листьев кукурузы в зависимости от уровня засоления.
Дата
Вариант
ВЫВОДЫ
1. Фактор засоления отрицательно сказывается на жизнедеятельности растений кукурузы. Засоление приводит к подавлению ростовых процессов, снижению оводнённости, водоудерживающей способности и интенсивности транспирации листьев кукурузы. В анатомо-морфологической структуре ассимиляционной поверхности наблюдаются закономерные изменения в сторону усиления ксероморфизма.
2. Величина отрицательного влияния засоления находится в прямой зависимости от концентрации соли в субстрате.
3. Обработка 6-БАП снимает угнетающее действие солей и способствует нормализации ростовых процессов, увеличению содержания воды и водоудерживающей способности, а также снижению интенсивности транспирации, что говорит о повышении адаптивных возможностей растительного организма.
4. Степень положительного влияния 6-БАП зависит от уровня засоления почвы. Максимальный эффект от обработки наблюдается у растений, выращенных при 0,2% концентрации NaCl в субстрате.
СПИСОК ЛИТЕРАТУРЫ
1. Аббасова З. И., Алиахвердиев С. Р., Зейналов Э. М., Гучейнова Н. Б. Конформационные изменения митохондрий при солевом стрессе.//Третий съезд Всероссийского общества физиологов растений: тезисы докладов, - Санкт-Петербург, - 1993., - 464 с.
2. Артемьева С.С, Солодилова О.С. Активность и изоферментный состав пероксидазы у С3- и С4- растений при солевом стрессе.//Тезисы участников 6-ой Пущинской школы-конференции молодых ученых «Биология – наука XXI века», 2002
3. Ахиярова Г.Р., Веселов Д.С. Гормональная регуляция роста и водного обмена при засолении..//Тезисы участников 6-ой Пущинской школы-конференции молодых ученых «Биология – наука XXI века», 2002
4. Балконин Ю. В., Строганов Б. П. Значение солевого обмена в солеустойчивости растений.//Проблемы солеустойчивости растений, - под ред. акад. ВАСХНИЛ Имамалиева А. И., - Ташкент, - изд-во «ФАН» Узбекской ССР, - 1989., - с. 45-64
5. Вавилов П. П. Растениеводство, - 5-е изд. - М.: Агропромиздат, - 1986
6. Васильев А. Е., Воронин Н. С., Еленевский А. Г. Ботаника: Морфология и анатомия растений. – (Учеб. Пособие для студентов пед. ин-тов по биол. и хим. спец.), - 2-е изд. Перераб. – М.: Просвещение, - 1988., - 480 с., ил.
7. Верзилов В.Ф. Регуляторы роста и их применение в растениеводстве. - М.: Наука, - 1971
8. Войников В. К., Рудиковский А. В., Побежимова Т. П., Варакина Н. Н. Физиологический стресс и стрессовые белки растений.//Второй съезд Всесоюзного общества физиологов растений: Тезисы докладов II ч., - М., - 1992., - 316 с.
9. Гамбург К.З. Регуляторы роста и рост растений. - М.: Наука, - 1964
10.Дерфлинг К. Гормоны растений. – М.: Мир, - 1985
11.Достанова Р. Х. Фенольный комплекс растений при засолении среды.//диссерт. на соискание ученой степени д. б. н. в форме научного доклада, - Новосибирск, - 1994.
12.Йонева Ж., Петров-Спиридонов А. Е. Биометрические показатели и осмотический потенциал органов растений в условиях хлоридного засоления.//Известия ТСХА, выпуск 3, - 1985, - с. 120-125
13.Кабузенко С. Н., Горшенков А. В., Володькина Л. С. Влияние хлоридного засоления и цитокинина на митотическую активность корней пшеницы и кукурузы // Физиол. и биохимия культурных растений. - 1995. - Т. 27, N1-2. - С.31-35.
14.Калинина Н. А., Драговоз И. В., Яворская В. К. Фитогормональный баланс корней кукурузы на фоне действия хлоридного засоления и 6-БАП.//Ученые записки ТНУ, Том 14 (53) № 1, 2002
15.Калинина Н.А., Кабузенко С.Н. Действие хлоридного засоления и регуляторов роста на содержание белка и активность пероксидазы в корнях кукурузы. //Ученые записки ТНУ, Том 13 (52) № 2, 2002
16.Касумов Н. А. Физиолого-биологические аспекты механизма действия солей на растительный организм. – Баку, - 1983., - 142 с.
17.Кидрей Т. А. Устойчивость С4 растений к засолению среды корнеобитания.//Вопросы экологии Волжско-Окского междуречья: Межвузовский сборник научных трудов. – Ковров: КГТА, - 1999, - с. 80-83
18.Клышев Л. К. Биохимические и молекулярные аспекты исследования солеустойчивости растений.//Проблемы солеустойчивости растений, - 1989., - 195 с.
19.Кудоярова Г. Р., Теплова И. Р., Докичева Р. А., Усманова И. Ю., Веселов С. Ю. Влияние 6-БАП на рост и содержание ауксинов в проростках пшеницы и кукурузы.//Иммуноанализ регуляторов роста в решении проблем физиологии растений, растеневодства и биотехнологии, - Материалы III конференции. – Уфа, 2000
20.Кулаева О. Н. Белки теплового шока и устойчивость растений к стрессу.//Статьи Соровского образовательного журнала. Биология, 1997
21.Кулаева О.Н. Гормональная регуляция физиологических процессов у растений на уровне синтеза РНК и белка. - М., - 1982
22.Кулаева О.Н. Цитокинины, их структура и функции. - М.: Наука, - 1973
23.Летние практические занятия по физиологии растений. – М.: Просвещение, - 1973., - 274 с.
24.Лебедев С. И. Физиология растений. – 3-е изд., перераб. и доп. – М.: Агропромиздат, 1988, - с. 519
25.Лосева А. С., Петров-Спиридонов А. Е. Устойчивость растений к неблагоприятным факторам среды. – М.: - изд-во МСХА, - 1983., - 47 с.
26.Луценко Э. К., Федюкина Е. М. Функционирование меристем и накопление ионов у растений при разных уровнях засоления.//Известия Северо-Кавказского научного центра высшей школы. Естественные науки, - 1



 В химическом отношении природные цитокинины и их синтетические заменители представляют собой производные 6-аминопурина с заместителем в аминогруппе при шестом атоме углерода пуринового кольца.
В химическом отношении природные цитокинины и их синтетические заменители представляют собой производные 6-аминопурина с заместителем в аминогруппе при шестом атоме углерода пуринового кольца.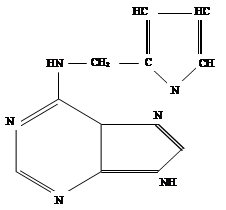
 Водоудерживающая способность = 100 - [ % ], где
Водоудерживающая способность = 100 - [ % ], где
 Дата
Дата