Ретикулярная формация получила название в связи с тем, что в ней клетки располагаются на значительном расстоянии друг от друга и разделены большим количеством волокон, идущих во все направления. Нервные клетки неоднородны: их тела и отростки имеют разную длину и толщину. Дендриты нейронов ретикулярной формации тянутся на большом протяжении и дают сравнительно небольшое количество ветвей. Аксоны сильно ветвятся. Ретикулярная формация проходит через продолговатый мозг, мост, средний мозг и заканчивается в неспецифических ядрах таламуса.
Ретикулярная формация – это комплекс диффузно расположенных нейронов спинного мозга и ствола головного мозга, имеющих обширные связи между собой, с различными нервными центрами и корой головного мозга. Впервые было описана в 1880 г. О. Дейтерсом. Затем ее подробно описали Бехтеров и Рамон-и-Кахаль. Она напоминает сеть нервных образований, представлена рассеянными клетками в спинном мозге, продолговатом мозге, варолиевом мосту, среднем (покрышке), промежуточном мозге (субсталамическое ядро, гипоталамус). Ретикулярная формация имеет обширные двусторонние связи (сама получает и посылает импульсы). Получает она импульсы по коллатералям от восходящих путей. Эти импульсы поддерживают ретикулярную формацию в активном состоянии, ее тонус (это неспецифическая афферентная система, потому что получает импульсы от разных раздражителей).
Основную роль выполняет ретикулярная формация ствола мозга. Ретикулярная формация занимает центральную часть на уровне продолговатого мозга, моста, среднего и промежуточного мозга.
Ретикулярная формация – это преимущественно неспецифическая часть ЦНС, т.е. основная масса ее нейронов и их проводников не являются центрами реализации определенных рефлексов. И тем не менее ретикулярная формация играет исключительно важную роль в осуществлении своих рефлекторных функций всеми ядрами ЦНС. Ее также называют «энергетической станцией ЦНС».
Ретикулярная формация мозга представлена сетью нейронов с многочисленными связями между собой и всеми структурами ЦНС. Она образует многочисленные афферентные пути: спиноретикулярный, церебеллоретикулярный, корково-подкорково-ретикулярный (от коры, базальных ганглиев, гипоталамуса), от структур каждого уровня ствола мозга (от среднего мозга, варолиева моста, продолговатого мозга), так и эфферентные нисходящие: ретикулоспинальные, ретикуло-корково-подкорковые, ретикуло-мозжечковые, к другим структурам ствола мозга. Таким образом, интенсивно обменивается чувствительной, так и двигательной импульсацией со всеми центрами ЦНС.
Ретикулярная формация оказывает генерализованное, тонизирующее, активирующее влияние на передние отделы головного мозга и кору больших полушарий (восходящая активирующая система ретикулярной формации), и нисходящее, контролирующее деятельность спинного мозга (нисходящая ретикуло-спинальная система).
Ретикулярная формация регулирует ряд вегетативных функций, в реализации которых принимают стартовые нейроны ретикулярной формации, запускающие процесс возбуждения внутри нейронов, отвечающих за дыхательные и сосудодвигательные функции. В ретикулярной формации продолговатого мозга расположены два ядра, одно из них отвечает за вдох, другое – за выдох – дыхательный центр. Их деятельность контролируется пневмотаксическим центром ретикулярной формации моста. Повреждение дыхательного центра (центральный механизм дыхания) приводит к остановке дыхания.
Ретикулярная формация участвует в регуляции вегетативных функций эмоциональных реакций через гипоталамус, вегетативные ядра черепных нервов и лимбическую систему.
Сосудодвигательный центр расположен в ромбовидной ямке IV желудочка, входящего в состав ретикулярной формации. Если же в эксперименте перерезать ствол мозга сразу же за продолговатым, то у животного резко падает артериальное давление, развивается коллапс. Было выявлено две группы нейронов. Нейроны одной группы усиливают свою активность при повышении артериального давления, а нейроны другой – при его снижении.
Ретикулярная формация оказывает влияние на сенсорные системы мозга: на остроту слуха, зрения, обонятельных ощущений. Ретикулярная формация обладает высокой чувствительностью к таким веществам, как адреналин и углекислый газ.
В ретикулярной формации продолговатого, среднего мозга имеются нейроны, реагирующие на болевые раздражения от мышц и внутренних органов, при этом создается ощущение тупой боли.
Ретикулярная формация регулирует вегетативные и соматические реакции, условно-рефлекторную и психическую деятельность.
Установлено, ретикулярная формации имеет большое значение в регуляции возбудимости и тонуса всех отделов ЦНС, т.е. оказывает восходящее и нисходящее возбуждающее и тормозящее влияние. Нисходящее влияние ретикулярной формации на спинной мозг может быть как активирующее, так и тормозящее.
Активирующее влияние ретикулярной формации на спинной мозг поступает по ретикулоспинальным путям. Они подавляют активность клеток Реншоу. Устранение активирующего и тормозного действия ретикулярной формации при высокой перерезке спинного мозга является одной из главных причин спинального шока.
Ретикулярная формация продолговатого мозга изменяет тонус мотонейронов мышц-разгибателей. Частично перекрещенный латеральный ретикулоспинальный тракт идет от ретикулярного гигантоклеточного ядра продолговатого мозга в спинной мозг, волокна этого тракта оканчиваются на его вставочных нейронах. В результате возбуждаются α- и γ-мотонейроны мышц-сгибателей конечностей и реципрокно с помощью клеток Реншоу тормозят α- и γ-мотонейроны мышц-разгибателей. Волокна неперекрещенного медиального тракта идут от каудальных и оральных ретикулярных ядер моста и тоже оканчиваются на интеронейронах спинного мозга. Эти волокна возбуждают α- и γ-мотонейроны мышц-разгибателей, а через клетки Реншоу тормозят α- и γ-мотонейроны мышц-сгибателей.
Ретикулярная формация изменяет активность γ-мотонейронов спинного мозга, что является причиной тонуса мышц, перерезка задних корешков приводит к исчезновению тонуса. Осуществляется контроль за положением тела в пространстве. Поддержание тонуса мышц в покое является одной из важнейших функций ретикулярной формации. Ретикулярная формация является как бы «настройщиком», поддерживающим мышцы в постоянном тонусе.
Тормозящее влияние ретикулярной формации на спинной мозг. Впервые тормозящее влияние раздражения ретикулярной формации открыто И.М. Сеченовым в опытах на лягушках (раздражение зрительных бугров кристалликами поваренной соли). Впоследствии во многих исследованиях было показано, что электрическое раздражение различных участков ретикулярной формации продолговатого мозга может тормозить и усиливать некоторые спинномозговые рефлексы. Большое значение имеет сила раздражения ретикулярной формации.
Восходящее влияние ретикулярной формации на кору больших полушарий может быть как активирующим, так и тормозным.
Активирующее влияние ретикулярной формации среднего мозга оказывает активизирующее влияние на кору больших полушарий. Если перерезать ствол мозга между верхними и нижними холмиками четверохолмия, то нарушается поступление в кору больших полушарий возбуждения по сенсорным системам – зрительной и обонятельной. На ЭЭГ регистрируются медленноволновые регулярные ритмы, характерные для сна. У животного нарушен контакт с внешним миром, не реагирует на световые и обонятельные раздражители.
Г. Мэгун и Дж. Моруцци (1949) показали, что стимуляция некоторых участков ретикулярной формации вызывает на ЭЭГ потенциалы, характерные для бодрствования, т.е. медленные ритмы сменяются высокочастотными ритмами.
Тормозное влияние ретикулярной формации на кору больших полушарий. При раздражении верхних отделов ретикулярной формации мозгового ствола животное впадает в состояние глубокого слоя (центры сна – Бремера, Чанга, Гессе). При этом на ЭЭГ возникает реакция синхронизации ЭЭГ-ритмов. Таким образом, ретикулярная формация участвует в формировании цикла «сна и бодрствования».
Ретикулярная формация ствола мозга, в отличие от неспецифических ядер таламуса, оказывает длительное и генерализованное активирующее влияние на кору больших полушарий. Она поддерживает кору в активном состоянии. Кора тоже влияет на ретикулярную формацию, но это уже волевое сознательное влияние.
Степень воздействия ретикулярной формации на кору больших полушарий головного мозга снижается под влиянием некоторых фармакологических веществ. Этим определяется снотворное и наркотическое действие барбитуратов. Во время сна ретикулярная формация осуществляет контроль за сознанием и эмоциями. Этим обусловлена, например, реакция спящей матери, которая из всех доносящихся до нее звуков чутко выделяет плач своего ребенка и мгновенно просыпается.
Ретикулярная формация стимулируется не только нервными влияниями с различных рецептивных полей, но и гуморально. Она чувствительна к гормонам, продуктам метаболизма и медикаментам. Введение гормона адреналина в кровь возбуждает ретикулярную формацию среднего мозга, а его восходящие стимулирующие влияния повышают функции коры больших полушарий. Увеличение парциального давления углекислого газа возбуждает ретикулярную формацию дыхательного центра и стимулирует дыхание.
Как показал П.К. Анохин, аминазин блокирует ретикулярную формацию. Снотворные вещества, например, производные барбитуратовой кислоты (барбамил, тиопентал натрия, гексенал и др.) угнетают через кровь ретикулярную формацию, снимая ее восходящее стимулирующее влияние на кору большого мозга, что способствует смене фаз бодрствования и сна.
Таким образом, ретикулярная формация участвует в регуляции деятельности вегетативной, соматической и психической деятельности организма.
Физиология мозжечка
Мозжечок представляет собой образование, расположенное позади больших полушарий мозга над продолговатым мозгом и мостом мозга (рис. 43).
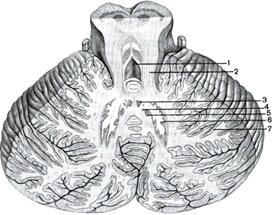
Рис. 43. Строение мозжечка:
1 – IV желудочек; 2 – верхняя мозжечковая ножка; 3 – ядро шатра; 4 – шаровидное ядро; 5 – пробковидное ядро; 6 – зубчатое ядро; 7 – кора мозжечка
Он является отделом преимущественно соматического контроля, координируя сложные и автоматизированные движения, регулирует позу, движения, дополняя и исправляя деятельность остальных двигательных центров. В осуществлении этой функции важную роль играют обширные связи мозжечка с другими соматическими отделами ЦНС и рецепторным аппаратом. В мозжечок поступает информация от рецепторов мышц, суставов, сухожилий, кожи, вестибулярной, слуховой, зрительной систем.
Выделяют три структуры мозжечка: древний мозжечок, старый мозжечок, новый мозжечок.
Древний мозжечок – состоит из клочка, узелка и нижней части червя (язычка), имеет афферентный вход от вестибулярных ядер. Это объясняет значение мозжечка в регуляции равновесия.
Старый мозжечок – состоит из центральной дольки, вершины, пирамиды, язычка червя и четырехугольной дольки полушарий. Имеет афферентные входы от спинного мозга, от проприорецепторов мышц, сухожилий, надкостницы, оболочек суставов.
Новый мозжечок – состоит из полушарий и части червя (ската, листка и бугра); получает афферентные входы от разных областей коры, зрительных и слуховых сенсорных систем, участвует в анализе зрительных и слуховых сигналов и в организации соответствующих реакций.
Белое вещество мозжечка содержит три парных ядра (шатра, промежуточные и зубчатые), это главный эфферентный выход мозжечка на моторные центры головного мозга.
Кора мозжечка состоит из трех слоев и имеет специфическое гистологическое строение (рис. 44).
Верхний, или молекулярный, слой образован параллельными волокнами разветвлений дендритов и аксонов второго и третьего слоя. В нижней части расположены корзинчатые и звездчатые клетки, обеспечивающие взаимодействие клеток Пуркинье.
Второй (средний) слой коры состоит из выстроенных в один ряд клеток Пуркинье (грушевидные клетки). Они имеют самую мощную в ЦНС дендритную систему (дендритное поле одной клетки Пуркинье может содержать до 60 тыс синапсов). Аксоны этих клеток являются единственным путем, с помощью которого кора мозжечка передает информацию в его ядра и ядра структуры большого мозга. Вся информация, приходящая в мозжечок, передается клетками Пуркинье. Они тормозят ядра мозжечка, а через них активность ретикулоруброспинальных нейронов, нейронов ядра Дейтерса, дающих начало вестибулоспинальному тракту.
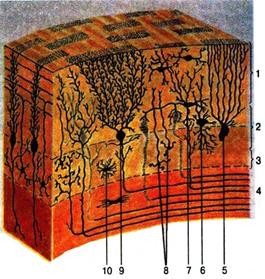
Рис. 44. Клеточные слои в коре мозжечка:
1 – молекулярный слой; 2 – слой грушевидных нейронов; 3 – зернистый слой; 4 – белое вещество; 5 – глиальная клетка; 6 – большая нервная клетка
(клетка Гольджи); 7 – корзинчатая нервная клетка; 8 – малые нейроциты;
9 – ганглиозные нервные клетки (клетки Пуркинье); 10 – астроцит
Под клетками Пуркинье лежит третий слой – гранулярный, состоящий из клеток-зерен, число которых достигает 10 млрд. Их аксоны поднимаются вверх, образуя дорожки контактов с клетками Пуркинье, здесь же расположены клетки Гольджи.
Мозжечок связан с мозговым стволом тремя парами ножек: верхние соединяют его со средним мозгом, средние – с мостом, нижние – с продолговатым мозгом. Ножки мозжечка обеспечивают прямые и обратные связи с другими отделами мозга.
Верхние ножки – гладкая эфферентная система (зубчато-красноядерно-спинномозговой путь). Через верхние ножки информация идет в таламус, варолиев мост, красное ядро, ядра стола мозга, в ретикулярную формацию и кору больших полушарий, центральную извилину.
Через средние ножки к нему поступают двигательные сигналы от кортикоспинального (пирамидного) пути. Кора мозжечка обрабатывает эту информацию, затем через ядра мозжечка и его верхние ножки направляет двигательные импульсы к красным ядрам среднего мозга, который передает их вестибулярным ядрам и далее α-мотонейронам спинного мозга.
Через задние ножки чувствительные импульсы от проприорецепторов скелетных мышц, кожи, вестибулярного аппарата проходят к мозжечку. Благодаря связям мозжечка с различными структурами ЦНС, он координирует двигательные реакции организма, вносит в них необходимые поправки, обеспечивая точность, например, при быстром переходе от сгибания конечностей к разгибанию и, наоборот (при ходьбе).
Таким образом, посредством своих трех пар ножек мозжечок подключен к экстрапирамидному пути на трех уровнях:
1) базальные ганглии (через них к коре головного мозга);
2) средний мозг – красные ядра;
3) вестибулярные ядра.
Нейроны мозжечка не имеют прямого выхода на спинальные мотонейроны, а действуют на них через корково-стволовые моторные центры.
Общей функцией мозжечка является затормаживание неправильной импульсации в экстрапирамидной системе, тем самым повышение точности тех движений, которые реализуются по этому пути.
Регуляция мышечного тонуса, позы и равновесия осуществляется преимущественно древним мозжечком и частично старым мозжечком. Мозжечок способен оценить состояние мышц, положение тела в пространстве и произвести перераспределение мышечного тонуса, изменить позу тела и сохранить равновесие. Осуществляет тонкую настройку вестибулярных рефлексов, в том числе рефлекторное поддержание вертикальной позы.
Функциональный тест: сохранение равновесия в позе
Ромберга.
Координация позы и выполняемого целенаправленного движения осуществляется старым и новым мозжечком, входящим в промежуточную зону. Осуществляет координацию позы и выполнение целенаправленного движения в пространстве, а также исправляет направление движения. Функциональный тест: пальценосовая и пяточно-коленная пробы.
Программирование целенаправленных действий осуществляется новым мозжечком. В коре нового мозжечка моторная импульсация, направляемая от высших соматических центров, преобразуется в программу движений. Это программа передается через зубчатое ядро мозжечка и вентральное латеральное ядро таламуса и поступает в кору. Обрабатывается в премоторной и моторной зонах и через пирамидную и экстрапирамидную системы осуществляется целенаправленное движение. Функциональный тест: проба на адиадохокинез.
Таким образом, мозжечок участвует в произвольной и непроизвольной регуляции функции скелетных мышц, обеспечивая целенаправленные движения, делая их плавными, точными, соразмерными.
Мозжечок выполняет вегетативные функции – участвует в согласовании вегетативного обеспечения соматической деятельности организма посредством ретикулярной формации, вегетативной нервной и эндокринной системы.
Тормозное влияние мозжечка обеспечивают соразмерность произвольных и непроизвольных движений скелетных мышц и целенаправленность. В мозжечке большое количество тормозных нейронов (клетки Гольджи, Пуркинье и т.д.). К этим тормозным клеткам через задние, средние ножки мозжечка постоянно поступает информация от коры головного мозга, проприорецепторов мышц, сухожилий, суставов, вестибулярного аппарата, ядер Дейтерса, продолговатого мозга. Вся эта информация передается им через лиановидные и моховидные нервные волокна, возбуждает клетки Пуркинье и таким образом усиливает их тормозное влияние на ядра мозжечка. Сигналы, передаваемые моховидными нервными волокнами, снижают активность клеток и уменьшают тормозное влияние коры мозжечка на его ядра. В итоге мозжечок регулирует произвольные пирамидные и непроизвольные экстрапирамидные влияния на α-мотонейроны передних рогов спинного мозга, воздействуя таким образом на перераспределение тормозных и возбуждающих импульсов, на тонус скелетных мышц.
Основные функции мозжечка:
1) контролирует значительную часть команд, поступающих в спинной мозг по нисходящим путям;
2) участвует в осуществлении статических и статокинетических рефлексов, регулирует работу всех центров, участвующих в осуществлении автоматических движений (ходьба);
3) участвует в регуляции тонуса всех центров, повышает устойчивость реакции в нервных центрах (соматических, вегетативных);
4) регулирует взаимодействие центров. Благодаря этому координирует двигательные реакции, вносит в них необходимые поправки, обеспечивая их точность.
Основные симптомы поражения мозжечка:
1) нарушение равновесия;
2) интенционное дрожание (тремор);
3) нистагм головы и глаз;
4) мышечная гипотония.
При поражении червя отмечаются нарушение равновесия, неустойчивость при стоянии и ходьбе, атаксия туловища, нарушение статики. Наблюдается падение в разные стороны, вперед и назад.
Поражение полушарий мозжечка приводит к нарушению выполнения локомоторных проб. Происходит нарушение координации в конечностях (на стороне очага) и руке. Больной падает на сторону пораженного полушария, не попадает пальцем в нос.
Влияние мозжечка на произвольные движения осуществляется благодаря наличию двусторонних связей мозжечка и коры больших полушарий. Мозжечок регулирует активность нейронов коры больших полушарий. Импульсы из мозжечка в кору идут через таламус. При удалении или повреждении мозжечка меняется механизм произвольных движений, осуществляемых при участии коры. В результате этого движения становятся размашистыми, теряется их точность – атаксия. Мозжечок регулирует устойчивость реакций в нервных центрах в точном соответствии с поставленной задачей.
Промежуточный мозг
Анатомически промежуточный мозг является продолжением мозгового слоя, находится между средним и полушариями большого мозга, включает III желудочек и образования, формирующего его стенки. В промежуточном мозге выделяют верхний отдел – эпиталамус, средний отдел – таламус, нижний отдел – гипоталамус, задний отдел – метаталамус.
Образования промежуточного мозга вместе с полосатым телом составляют анатомическую основу для врожденных рефлекторных реакций, объединяемых в понятие инстинктов.
Главными образованиями помежуточного мозга являются таламуса – зрительный бугор и гипоталамус – подбугорная область.
Таламус (зрительный бугор) – основная часть промежуточного мозга. Включает в себя собственно зрительный бугор, латеральные и медиальные коленчатые тела. К этим телам подходят ручки холмиков среднего мозга. Латеральное коленчатое тело связано с верхними холмиками – вместе с ним является подкорковым центром зрения, медиальное – с нижними холмиками – соответственно они – подкорковые центры слуха. Аксоны нейронов коленчатых тел направляются в кору больших полушарий, в корковые центры зрения и слуха.
В таламусе содержится около 40 ядер. Среди ядер таламуса выделяют следующие группы:
1) комплекс специфических, или релейных, т.е. переключательных таламических ядер, через которые проходит афферентная информация определенной модальности;
2) неспецифические таламические ядра, не связанные с проведением афферентной информации и проецирующиеся на кору больших полушарий более диффузно, чем специфические ядра;
3) ассоциативные ядра таламуса, получающие раздражения от других ядер таламуса и передающие эту информацию на ассоциативные области коры головного мозга.
Таламус является местом переключения всех чувствительных проводников, идущих от экстеро-, проприо- и интерорецепторов, поднимающихся в кору головного мозга. Таламус является высшим центром болевой чувствительности, а кора позволяет точно локализовать место болевого раздражения и формировать ощущение и отношение к боли.
Таламус – чувствительное ядро подкорки, его называют коллектором чувствительности, так как к нему сходятся аффрентные пути всех рецепторов, кроме обонятельного и слухового. В нем происходит обработка всей информации, поступающей в кору из спинного мозга и подкорковых структур. Он обладает тремя основными функциями: переключающей, интегративной, модулирующей.
Главная функция специфических ядер состоит в перекодировании, переработке информации и переключении афферентного потока импульсов в соматосенсорную зону коры больших полушарий.
Специфические ядра бывают сенсорные и несенсорные. К сенсорным ядрам относятся: вентральные задние, латеральные и медиальные коленчатые тела. Вентральные задние являются главным реле для переключения соматосенсорной импульсации, которая поступает по волокнам медиальной петли и примыкающим к ней волокнам других афферентных путей. В них переключаются тактильная, проприорецептивная, вкусовая, висцеральная, частично температурная и болевая импульсации. Импульсация из вентральных задних ядер проецируется в соматосенсорную кору постцентральной извилины, где формируются соответствующие ощущения, отражающие реальную деятельность.
Латеральное коленчатое тело является подкорковым зрительным центром, оно получает афферентные импульсы из сетчатки глаза и верхних бугров четверохолмия и посылает импульсы в затылочную зону коры больших полушарий, где располагается первичный корковый зрительный центр.
Медиальное коленчатое тело является подкорковым слуховым центром, оно получает афферентные импульсы из латеральной петли и нижних бугров четверохолмия и посылает импульсы в височную долю коры больших полушарий, где располагается первичный корковый слуховой центр.
Несенсорные релейныe ядра таламуса – переднее вентральное, переднее дорсальное и переднее медиальное ядра, переключают в кору несенсорную импульсацию, поступающую в таламус, и получают афферентную информацию от базальных ганглиев, зубчатого ядра мозжечка, красного ядра среднего мозга, которая затем проецируется в моторную и премоторную зоны коры. Эти ядра выполняют моторную функцию, которая в виде сложных двигательных программ поступает из мозжечка и базальных ганглиев.
В несенсорные ядра передней группы импульсация поступает в основном из мамиллярных тел гипоталамуса и проецируются в лимбическую кору, от нее гиппокампу и опять к гипоталамусу, образуя замкнутый круг: гипоталамус – гиппокамп – гипоталамус, формирующий эмоциональное поведение.
В вентральных ядрах переключается импульсация от базальных ганглиев, зубчатого ядра мозжечка, красного ядра среднего мозга, которая после этого проецируется в моторную и премоторную кору большого мозга, обеспечивая реализацию двигательных программ мозжечка и базальных ядер.
Специфические ядра таламуса получают также нисходящие корковые регулирующие влияния из своей проекционной зоны.
Неспецифические ядра таламуса являются ретикулярной формацией промежуточного мозга и местом переключения восходящих активирующих влияний, которые направляются от ретикулярной формации ствола в кору большого мозга. Эти ядра имеют многочисленные входы от других ядер таламуса и сенсорных систем. Нейроны неспецифических ядер таламуса посылают свои аксоны диффузно на всю кору, во все его слои. К неспецифическим ядрам поступают афферентные импульсы из моторных центров ствола мозга, ядер мозжечка, базальных ганглиев и гиппокампа, а также от коры большого мозга, особенно лобных долей. Эти ядра имеют эфферентные выходы на другие таламические ядра, кору большого мозга, а также нисходящие пути к другим структурам ствола мозга. Благодаря этим связям неспецифические ядра выступают в роли интегрирующих посредников между стволом мозга и мозжечка с новой корой, лимбической системой и базальными ядрами.
Неспецифические ядра оказывают на кору больших полушарий модулирующее влияние.
Г. Джаспер на основании электрофизиологических экспериментов пришел к выводу, что неспецифическая система таламуса принимает участие в быстрой и кратковременной активации коры. Активирует лишь те структуры, которые должны участвовать в выполнении конкретных рефлекторных актов.
Ассоциативные ядра таламуса включают в себя медиодорсальное, латеральное дорсальное ядро и ядро подушки. Волокна к этим ядрам приходят от других ядер таламуса. Эфферентные выходы от этих ядер направляются в ассоциативные поля коры. В свою очередь кора мозга посылает волокна к ассоциативным ядрам, регулируя их функцию.
Главной функцией этих ядер является интегративная функция, которая выражается в объединении деятельности таламических ядер, так и различных зон ассоциативной коры больших полушарий.
Медиодорсальное ассоциативное ядро – это важный ассоциативный центр, интегрирующий поток информации как таламических, так и внеталамических структур (центрального серого вещества ствола мозга, гипоталамуса, амигдалы, гиппокампа) и переадресовывающий ее в лобную и лимбическую кору. Участвует в формировании эмоциональной и поведенческой двигательной активности, а также запоминании информации.
Подушка получает афференты от наружного и внутреннего коленчатых тел и неспецифических ядер таламуса, эфферентные пути от нее идут в височную, затылочную и теменную кору. Подушка участвует в гностических (узнавание предметов, явлений), речевых и зрительных функциях (интеграция слова со зрительным образом).
Латеральные задние ядра связаны с подушкой, получают зрительную и слуховую импульсацию от коленчатых тел, направляющуюся соответственно в зрительную кору (затылочную область) и слуховую (височную область). Также получают соматовисцеральную импульсацию от вентрального ядра таламуса, и, переработав ее, направляются в ассоциативную теменную кору и участвуют в сложных механизмах зрительного и слухового восприятия.
Таламус обеспечивает двигательные и вегетативные реакции, связанные с сосанием, жеванием, глотанием, смехом. В таламусе переключаются, обрабатываются, проводятся все импульсы, идущие от рецепторных входов спинного мозга, ствола и мозжечка, к коре мозга. В таламусе заканчивается ряд нисходящих путей от коры, лимбической системы и базальных ганглиев.
В клинике симптомами поражения зрительных бугров являются сильные головные боли, расстройство сна, нарушение чувствительности как в сторону повышения, так и понижения, нарушение движений, их точности, возникновение насильственных, непроизвольных движений.
Обобщая вышесказанное можно считать, что таламус, как и любой отдел ЦНС выполняет две основные функции: проводниковую и рефлекторную.
Проводниковая функция осуществляется как по восходящей – проведение чувствительной импульсации в кору, так и нисходящей – регуляция рефлекторной активности практически всех нижележащих соматических и вегетативных центров.
В таламус поступает абсолютно вся афферентация центральной нервной системы. Главной функцией таламуса является интеграция всех видов чувствительности. Для анализа внешней среды недостаточно сигналов от отдельных рецепторов. Здесь происходит сопоставление информации, получаемой по различным каналам связей, подобно секретарю, «отбирают» наиболее биологически значимую в настоящий момент времени чувствительную импульсацию, которую затем переправляют в кору головного мозга с целью формирования ею рефлекторного ответа, наиболее значимого в данных условиях. Моторная функция таламуса заключается в том, что он координирует сложные безусловно рефлекторные акты, т.е. инстинкты, позволяя в данный конкретный момент времени проявляться одним и подавляя другие.
Таламус участвует в переработке сенсорных стимулов, идущих к коре больших полушарий, а также регулирует цикл бодрствование – сон.
Гипоталамус (подбугровая область) – филогенетически наиболее старая часть промежуточного мозга и входит в состав лимбической системы. Находится вентральнее таламуса и включает в себя собственно подбугорную область и ряд образований, расположенных на основании мозга (подбугорье). К нему относятся: зрительный перекрест и зрительный тракты; серый бугор, воронка и гипофиз; сосцевидные тела.
Гипоталамус имеет сложное строение (рис. 45).
Морфологически в гипоталамусе выделяют около 50 пар ядер. Ядра гипоталамуса имеют мощное кровоснабжение.
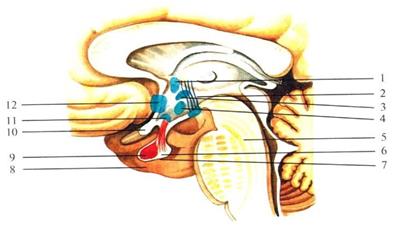
Рис. 45. Сагиттальный срез гипоталамуса:
1 – паравентрикулярное ядро; 2 – сосцевидно-таламический пучок; 3 – дорсомедиальное гипоталамическое ядро; 4 – вентромедиальное гипоталамическое ядро; 5 – мост мозга; 6 – супраоптический гипофизарный путь; 7 – нейрогипофиз; 8 – гипофиз; 9 – аденогипофиз; 10 – зрительный перекрест; 10 – супра
оптическое ядро; 11 – преоптическое ядро
Выделяют три группы ядер гипоталамуса: передняя, средняя, задняя. В преоптической области (передней гипоталамической) выделяют медиальное и латеральное преоптические ядра, паравентрикулярное и супраоптическое ядра, переднее гипоталамическое и супрахиазмальное ядра.
В медиальной части средней гипоталамической области находятся дорсомедиальное и вентромедиальное гипоталамические ядра, ядра воронки (дугообразное ядро). В латеральной части средней части гипоталамуса расположены латеральное гипоталамической, серобугорное, серобугорно-сосцевидное и перифорникальное ядра.
Задняя гипоталамическая область содержит медиальное и латеральное ядра сосцевидного тела, заднее гипоталамическое ядро.
Зрительный перекрест образуется, когда перекрещиваются волокна зрительных нервов (II пары черепных нервов), идущих из сетчатки глаз. Перекрещиваются только медиальные волокна, несущие импульсы от внутренних половин сетчатки обеих глаз. После перекреста идут зрительные тракты, которые направляются к латеральным коленчатым телам (подкорковым центрам зрения).
Серый бугор – непарный полый вырост мозга с тонкими стенками из серого вещества, в котором есть несколько групп ядер – это подкорковый центр вегетативной нервной системы (высший вегетативный центр), контролирующий работу внутренних органов, обмен веществ. Серый бугор книзу сужается (вытягивается) и образует воронку, на котором подвешен гипофиз (нижний придаток мозга). Гипофиз, также как эпифиз, является нейрогенной эндокринной железой, который вырабатывает гормоны. Его гормоны контролируют работу других эндокринных желез, осуществляют регуляцию обмена веществ, работу внутренних органов, рост и развитие организма.
Сосцевидные тела – два небольших возвышения белого цвета, внутри ядра – это подкорковые центры обоняния. Они получают информацию из проекционного центра обоняния – нейронов парагиппокампальной извилины и крючка, а также связаны с передними ядрами таламуса и верхними холмиками среднего мозга.
Капилляры гипоталамуса высокопроницаемы для крупномолекулярных белковых соединений, к которым относятся нуклеопротеиды, чем объясняется высокая чувствительность гипоталамуса к нейровирусным инфекциям, интоксикациям, гуморальным сдвигам. У человека гипоталамус окончательно созревает к 13–14 годам, когда заканчивается формирование гипоталамо-гипофизарных нейросекреторных связей.
В заднем отделе гипоталамуса расположены ядра, образованные мелкими нервными клетками, которые связаны с передней долей гипофиза особой системой кровеносных сосудов. Гипоталамусу свойственна очень важная нейросекреторная функция, в частности регуляции выделения гормонов гипофизом.
Гипоталамус имеет афферентные связи с обонятельным мозгом, базальными ганглиями, таламусом, гиппокампом, орбитальной височной и теменной корой. Эфферентные пути представлены: мамиллоталамическим, гипоталамо-таламическим, маммилло-тегментальным, гипоталамо-гипокампальным трактами. Гипоталамус посылает импульсы вегетативным центрам ствола мозга и спинного мозга.
Гипоталамус является главным подкорковым центром, регулирующим вегетативные функции. От ядер гипоталамуса начинается задний продольный пучок, который проходит по стволу головного мозга и окаймляет центральный канал спинного мозга; его волокна обеспечивают согласованную деятельность парасимпатических и симпатических ядер, тем самым участвуя в вегетативной иннервации внутренних органов. Раздражение ядер передней группы сопровождается парасимпатическими эффектами: сужение зрачка, брадикардия, снижение АД, усиление секреции и моторики ЖКТ. Супраоптическое и паравентрикулярное ядра участвуют в регуляции водного и солевого обмена за счет выработки антидиуретического гормона. Передняя группа ядер названа «трофотропной системой мозга», обеспечивающей восстановление гомеостаза и энергетических ресурсов организма.
Стимуляция задней группы активирует симпатические эффекты: расширение зрачка, тахикардию, повышение кровяного давления, торможение моторики и секреции ЖКТ. Эта часть была названа «эрготропной системой мозга».
Гипоталамус обеспечивает механизмы терморегуляции. Ядра передней группы ядер содержат нейроны, отвечающие за теплоотдачу. Задняя группа ядер отвечает за процесс теплопродукции, в результате которого температура повышается.
В пищевом центре находятся нейроны, обладающие хеморецепторной чувствительностью к некоторым веществам: глюкозе, аминокислотам, жирным и органическим веществам, гормонам (инсулин, гастрин, адреналин).
В области вентромедиальных ядер гипоталамуса и в латеральных его отделах обнаружены нейроны, избирательно чувствительные к содержанию в крови глюкозы. Одни нейроны активируются при снижении глюкозы в крови, другие, наоборот, при возрастании ее содержания. В области супраоптических и паравентрикулярных ядер гипоталамуса обнаружены нейроны, чувствительные к уровню осмотического давления крови, в области заднедорсального гипоталамуса – к норадреналину и т.д. Электрическая стимуляция указанных о





