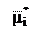Классические и новые разделы физиологии растений. Их предмет и задачи. Важность физиологии и биотехнологии растений для общества и устойчивого развития человечества.
Основные классические фундаментальные разделы ФЗР: - фотосинтез - дыхание - водобмен- минеральное питание - рост и развитие - физиология стресса
В последние годы появились новые фундаментальные разделы ФЗР, такие как
- сигнальная трансдукция и передача информации - внутриклеточный транспорт - иммунитет растений - геномика (раздел молекулярной генетики, посвящённый изучению генома и генов живых организмов.) - протеомика (изучает белки и их взаимодействия в живых организмах.
Идентификация, характеристика и количественный учет ВСЕХ белков метаболического пути, органеллы, клетки, ткани, органа и всего организма.)
- феномика
- метаболомика (это «систематическое изучение уникальных химических „отпечатков пальцев“ специфичных для процессов, протекающих в живых клетках» — конкретнее, изучение их низкомолекулярных метаболических профилей. Метаболом представляет собой совокупность всех метаболитов, являющихся конечным продуктом обмена веществ в клетке, ткани, органе или организме. В то время как данные об экспрессии мРНК генов и данные протеомного анализа не раскрывают полностью всего того, что может происходить в клетке, метаболические профили могут дать мгновенный снимок физиологических процессов в клетке. Одна из задач системной биологии и функциональной геномики — интегрирование данных протеомики, транскриптомики и метаболической информации для получения более целостного представления о живых организмах. Метаболомика — это научное изучение химических процессов, в которые вовлечены метаболиты.)
- иономика
Новые прикладные отрасли ФЗР: - молекулярная селекция - генетическая модификация
- молекулярные основы урожайности - устойчивое развитие и устойчивое сельское хозяйство
- получение биотоплива – агрофеномика - молекулярное сельское хозяйство - биоинформатика растений - системная биология растений и т.д.
Сущность проблемы использования генетически-модифицированных растений. Преимущества ГМ-растений. Предмет и задачи феномики растений.
По данным Всемирного института питания здоровое питание и доступность свежей растительной пищи имеют большее значение для здоровья человека, чем все другие факторы, такие как качество и доступность жилья, лекарства, социальное положение и физкультура.
Только 20-25% населения планеты имеет постоянный доступ к здоровой пище.
Домыслы о вреде ГМО можно воспринимать только как лженауку. На сегодняшний день не известно ни одного ГМО продукта, который бы статистически-достоверного вызывал токсичность или аллергию у человека или животных.
Современные подходы к селекции основываются на новом разделе ФЗР — феномике.
Генетически трансформировать растение — не проблема. Методы исключительно просты сейчас.
Но отобрать линии с требуемыми физиологическими параметрами, например, с устойчивостью к определенным стресс-факторам, является большой проблемой, так как требуется протестировать тысячи растений в стандартных условиях.
Находящиеся на различных стадиях испытания трансгенные культуры, обладающие признаками устойчивости к экологически более безопасным гербицидам, болезням и вредителям, толерантные к засухе, засолению, холоду, с повышенным содержанием белков витаминов, микроэлементов, полезных жирных кислот и углеводов, несомненно привлекут покупателей, и никакие запугивания дезинформированных защитников окружающей среды не смогут остановить прогресс.
Сравнение циклического и нециклического фотофосфорилирования.
| Процесс
| Нециклическое
| Циклическое
|
| Путь электронов
| Нециклический
| Циклический
|
| Первый донор (источник) электронов
| Вода
| ФС I (Р700)
|
| Последний акцептор (место назначения) электронов
| НАДФ+
| ФС I (Р700)
|
| Продукты
| Основные:
АТФ, НАДФН
Побочные: О2
| Основные:
АТФ
|
| Участвующие ФС
| I и II
| I
|
Образование ацетил-СоА, его участие в метаболизме. Общая схема и суммарное уравнение цикла Кребса (трикарбоновых кислот).
В аэробных условиях главный путь преобразования ПВК – окислительное декарбоксилирование при помощи пируватдегидрогеназного мультиферментного комплекса, составной частью которого является коэнзим А (СоА или СоА-SH). В результате образуется ацетил-СоА. Окисление пирувата и образование ацетил-СоА (активированной уксусной кислоты) – очень сложный процесс, рассматриваемый подробно в курсе биохимии. Ограничимся суммарным выражением этого процесса:СН3СОСООН + НАД+ + СоА-SH →СН3СО ~ S-СоА + НАДН + Н+ + СО2
В молекуле ацетил-СоА (СН3СО ~ S-СоА) тиоловая группа участвует в образовании эфирной связи уксусной кислоты. Тиоэфирная связь относится к высокоэнергетическим связям.Рассмотренный путь образования ацетил-СоА является главным, но не единственным. Ацетил-СоА может образовываться при окислении аминокислот и жирных кислот. Это значит у высших растений дыхательным субстратом могут быть все основные запасенные вещества: углеводы, белки, жиры. В анаэробных условиях пируват вступает в реакции брожжения. При брожжении, в основном спиртовом, пируват декарбоксилируется с образованием уксусного альдегида при участии карбоксилазы, а затем восстанавливается до этилового спирта ферментом алкагольдегидрогеназой:СН3СОСООН → СН3СОН + СО2
СН3СОН + НАДН + Н+ → СН3СН2ОН + НАД+.
Ни одна из реакций не сопровождается ни образованием АТФ, ни восстановлением коферментов. Образовавшийся во время гликолиза НАДН растрачивается на восстановление уксусного альдегида.В последние годы выявлено молочно-кислое брожжение. В этом случае из одной молекулы гексозы образуются две молекулы молочной кислоты: С6Н2О6 → 2СН3СНОН СООН. Таким образом, основная функция брожжения заключается в регенерации НАД+ и обеспечения гликолитических процессов окисленной формой кофермента в аэробных условиях. В аэробных условиях образовавшийся ацетил-СоА вступает в цикл Кребса. В цикле Кребса после реакций отнятия и присоединения воды, декарбоксилирования и дегидрирования ацетильный остаток, поступивший в цикл в виде ацетил-СоА, полностью расщепляется. Суммарная реакция записывается в следующем виде: СН3СО ~ S-СоА + 3Н2О + АДФ + Н3РО4 → НS-СоА + 2СО2 + 4[Н2] + АТФ
Цикл Кребса проходит одинаково у животных и растений. Это является еще одним доказательством единства происхождения. Цикл происходит в строме митохондрий. Рассмотрим его подробней: Первая реакция цикла – перенос ацетильного остатка от ацетил-СоА на щавелево-уксусную кислоту (ЩУК) с образованием лимонной кислоты (цитрат)
В ходе реакции, катализируемой цитратсинтазой, растрачивается макроэргическая связь ацетил-СоА, т. е. та энергия, какая была занесена в процессе окисления пирувата перед началом цикла. Это значит, как и гликолиз, цикл Кребса начинается не с запасания энергии в клетке, а с расходования. Подчеркнем, что цепь преобразований, образующих этот цикл и направленных, в конечном счете, на разрушение углеродного состава рядя кислот, начинается с их увеличения: двухуглеродный фрагмент (уксусная кислота) присоединяется к четырехугольному фрагменту ЩУК с образованием шестиуглеродной трикарбоновой кислоты цитрата, которая может запасаться в клетках в больших количествах.
Таким образом, цикл Кребса – процесс каталитический и начинается не с катаболизма (разрушения), а с синтеза цитрата. Цитратсинтетаза, катализирующая эту реакцию, относится к регуляторным ферментам: она ингибируется НАДН и АТФ. НАДН – конечный продукт, в форме которого запасается энергия, освобождаемая в процессе дыхания. Чем активней цитратсинтетаза, тем быстрей пойдут и другие реакции цикла, быстрей пойдет дегидрирование веществ с образованием НАДН. Однако увеличение количества последнего вызывает ингибирование фермента, и цикл затормозится. Это пример реакции по принципу обратной связи. Следующая серия реакций – преобразование цитрата в активную изолимонную кислоту (изоцитрат). Она протекает при участии воды и по сути сводится до внутримолекулярного преобразования лимонной кислоты. Промежуточным продуктом этого преобразования является цис-аконитовая кислота. Катализируются обе реакции аконитатгидротазой. Затем изоцитрат дегидрируется с участием изоцитратдегидрогеназы, коферментом которой является НАД+. В результате окисления образуется щавелево-янтарная кислота (оксалосукцинат). Последняя кислота декарбоксилируется. Отсоединяющийся СО2 принадлежит ацетильному остатку, вступившему в цикл в виде ацетил-СоА. В результате декарбоксилирования образуется очень активная α-кетоглутаровая кислота (кетоглутарат). α-Кетоглутарат, в свою очередь, подвергается тем же изменением, которые происходят перед началом цикла с пируватом: одновременное окисление и декарбоксилирование.В реакции принимает участие α-кетоглутарат дегидрогеназный комплекс: α-кетоглутарат + НАД+ + СоА–SН →сукцинил-S-СоА + СО2 + НАДН + Н+ → сукцинил–S–СОА + АДФ + Н3РО4 → янтарная кислота + АТФ + СоА–SН
Освободившийся СО2 является другой частицей, которая отщепляется от ацетильного остатка. Образовавшийся в результате этих сложных преобразованием янтарная кислота (сукцинат) вновь дегидрируется, и образуется фумаровая кислота (фумарат). Реакция происходит с помощью сукцинатдегидрогеназы. Фумарат после присоединения молекулы воды легко преобразуется в яблочную кислоту (малат). В реакции принимает участие фумаратгидротаза. Яблочная кислота, окисляясь, преобразуется в ЩУК при участии НАД+ – специфической малатдегидрогеназы.Напомним, что ЩУК конечный продукт цикла Кребса – образуется и при фотосинтезе С4-растений (цикл Хетча – Слека) при карбоксилировании ФЕП на свету, и в темноте у растений типа САМ.Таким образом, цикл Кребса заканчивается и может начинаться сначала. Одно условие – подача новых молекул ацетил-СоА. Главное значение цикл Кребса состоит в запасании энергии, которая освобождается в результате разрушения пирувата, в макроэргических связях АТФ. Поставляя в клетку АТФ, цикл Кребса может являться регулятором других процессов, идущих с затратой энергии, таких как транспорт воды и солей, синтез и транспорт органических веществ. Чем быстрей проходит преобразование веществ в цикле, тем больше может синтезироваться АТФ, тем быстрей пойдут указанные процессы.
Физиологическая значимость процессов минерального питания, коэффициент накопления, органогены, определение макро- и микроэлементов, их представители. Первичные и вторичные макроэлементы. Тройное правило Арнона.
Растения могут поглощать и перемещать неорганические и органические вещества. Раньше считали, что для роста растений (фотосинтезирующих автотрофов) необходимы только минеральные элементы. Однако в последние годы показано, что многие водоросли требуют витамины В12, а также веществ, содержащих кобальт и т. д.
Для наземных растений основным источником воды и минеральных веществ служит почва, не менее важна и атмосфера, которая обеспечивает фотосинтезирующие растения СО2, и, в свою очередь, сама атмосфера поглощает О2 и воду, выделяемую при транспирации.
В системе растение-почва-атмосфера происходит значительный круговорот веществ, особенно воды и СО2.
Концентрация минеральных веществ в самих растениях в большинстве случаев мало связана с их концентрацией в среде. Например, соотношение K+/Na+ в растениях обычно составляет 5–20, тогда как в среде, как правило, больше Na+. С другой стороны, элементы, содержащиеся в окружающей среде в низких концентрациях, могут накапливаться в растениях в значительном количестве.
Эта способность растений характеризуется так называемым коэффициентом накопления:
| |
| |  |
Еще раз отметим, что ни присутствие, ни концентрация минеральных элементов в самих растениях не является критерием необходимости. Хорошо зарекомендовало себя в этом отношении (относительно необходимости) так называемое тройное правило Арнона, которое было предложено еще в 1939 году. Элемент признается необходимым в случае, когда: 1. Растение без него не может закончить свой жизненный цикл; 2. Другой элемент не может заменить функцию изучаемого элемента; 3. Элемент непосредственно включен в метаболизм растения.
Питательные элементы, которые используются растениями из субстрата в больших количествах (содержание их в золе 10–1–10–2 %), называют макроэлементами, а необходимые в значительно меньшем количестве (содержание 10–3 % и ниже) – микроэлементами.
Вода и СО2 рассматриваются отдельно, но в этом контексте необходимо считать их макроэлементами. К макроэлементам относятся K, Ca, Mg, N, P, S (Н2О и СО2), для галофитов следует добавить Na и Cl.
Микроэлементы: Mn, Zn, Cu, Cl, B, Mo, сюда же относится и Fe, который располагается на границе макро- и микроэлементов.
Водорослям необходим один или несколько из следующих элементов: Со, Si, I, V. Некоторым высшим растениям необходимы селен, кремний.
Кроме необходимых элементов для роста, в оптимальных пределах должны быть и другие факторы: рН, температура, обеспечение О2, градиент водного потенциала, а для зеленых растений – освещенность соответствующей интенсивности и протяженности.
Пассивный транспорт – перемещение веществ путем диффузии по градиенту электрохимического потенциала без затраты энергии (простая и, в какой-то мере, облегченная диффузия).
Активный транспорт – перемещение веществ против градиента электрохимического потенциала с затратой метаболической энергии, как правило в форме АТФ или редокс-цепей.
Для того чтобы понять механизмы трансмембранного переноса элементов минерального питания, остановимся на рассмотрении некоторых физико-химических закономерностях, определяющих движение ионов в растворе и мембране. Начнем с процессов пассивного транспорта (в частности вспомним законы диффузии).
Согласно первому закону Фика, поток (Ф) прямо пропорционален коэффициенту диффузии D и градиенту концентрации dC/dх в точке х в данный момент времени.
Ф = – D
Знак минус в правой части уравнения означает, что, если градиент положителен, т. е. концентрация при увеличении х возрастает, диффузионный поток направлен в противоположную сторону.
Однако при описании диффузии на большие расстояния в непрерывной системе уравнения (5.1) необходимо определить зависимость концентрации не только от расстояния, но и от времени. Поэтому целесообразно преобразовать первый закон Фика в дифференциальные уравнения в частных производных, которые обычно называют вторым законом Фика. В одномерном случае этот закон аналитически выражается следующим образом:
=
D

т. е. скорость изменения концентрации пропорциональна второй производной от концентрации на координате х. Из этого закона мы получаем очень важное соотношение:
х =
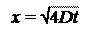
движение ионов через мембрану обусловлено градиентом электрохимического потенциала.
Обычно мембрана на внутренней стороне отрицательно заряжена по отношению к наружной поверхности. Это приводит к преимущественному поступлению катионов по сравнению с анионами. В этом случае физической движущей силой будет электрохимический потенциал  , в величину которого вносят свой вклад химический и электрический потенциалы:
, в величину которого вносят свой вклад химический и электрический потенциалы:
µi
| µi = µi + RT ln Ci + ZiFφ,
|
где – электрохимический потенциал i-иона в стандартных условиях; Zi – валентность i-иона; F – число Фарадея; φ – электрический потенциал.
Δ µi =

В реальных условиях описания процесса диффузии ионов в клетку нас интересует разность электрохимических потенциалов (Δφ) по обеим сторонам мембраны, т. е.. Используя уравнение (5.9) можно записать:

| (5.10)
|
µi = μi
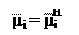
В случае равновесного состояния, когда электрохимический потенциал i-иона одинаков по обе стороны мембраны, т. е., получаем:
Примеры структуры субъединиц катионных каналов, их молекулярно-биологических и физиологических свойств.
В настоящее время на мембранах растительных клеток идентифицированы несколько видов катионных каналов. Это Са2+-канал и K+-каналы. В последние годы имеются доказательства о наличии на плазмалемме клеток высших растений Na+ -каналов. Характеристики кальциевого канала получены как на модифицированных искусственных бислойных мембранах, так и на нативной плазмалемме. Отличительной их особенностью по отношению к другим катионным каналам является их малый вклад в общую проводимость мембраны в обычном физиологическом состоянии. Са2+-канал идентифицирован в нестационарных условиях, т. е. при возбуждении клетки и возникновении потенциала действия (ПД). Все кальциевые каналы делят на две группы: потенциалзависимые и
рецепторуправляемые (хемозависимые). В зависимости от проводимости, времени жизни в открытом состоянии, скорости активации и инактивации потенциалзависимые Са2+-каналы делятся на L -, T -, N - и P -типы. Эффективными блокаторами кальциевых токов через мембрану служат ионы Ni2+, Cd2+, Co2+, Mn2+ и La3+. К блокаторам Са2+-каналов органической природы относятся три типа липофильных азотосодержащих соединений; в частности, верапамил, нифедипин и дильтиазем. Чувствительность к лигандам зависит от их строения, от типа канала и величины мембранного потенциала. Деполяризация мембраны, как правило, вызывает повышение степени связывания с каналом дигидро-
пиридинов. Различие в действии органических лигандов проявляется в случае Са2+-агонистов.
Фитогормоны модулируют работу Са2+-каналов, при этом внешний лиганд действует через G-белки. Кальциевые каналы, активируемые инозитол-1,4,5-трифосфатом, обнаружены в тонопласте. Са2+-каналы функционируют практически во всех мембранных структурах растительных клеток. Например, увеличение концентрации ионов в цитоплазме замыкающих клеток устьиц Vicia faba и протопластах Pisum sativum при действии гормонов связано с работой Са2+-каналов плазмалеммы. Из клеток колеоптилей удалось выделить мембранные белки, имеющие сайты связывания с ингибитором Са2+-каналов верапамилом. При встраивании этих белков в липидный бислой они проявляли свойства Са2+-каналов. Выделенные из клеток корней пшеницы ивстроенные в липидный бислой Са2+-каналы находились в закрытом состоянии при потенциалах отрицательнее –130 мВ. Каналы блокировались верапамилом (10–6 М) с внешней стороны мембраны. Са2+-каналы мембран растительных клеток характеризуются большим временем нахождения в открытом (от нескольких миллисекунд до секунд) и закрытом (до 100 с) состояниях по сравнению с животными клетками. Анализ зависимости транспорта ионов кальция через мембраны
плазматических визикул клеток колеоптилей кукурузы показал, что наиболее интенсивный поток их наблюдается в диапазоне потенциала от –50 до –70 мВ. Аналогично наибольшую активность Са2+-каналы плазмалеммы клеток водоросли Nitellopsis obtusa проявляли при потенциале фиксации –50 мВ.
Повышенный интерес к кальциевым каналам связан с особой ролью Са2+ как вторичного посредника, медиатора многих функционально важных процессов в клетке. Проводимость Са2+-канала для разных видов одновалентных ионов характеризуется следующим рядом: Rb+ > K+ > Cs+ > Na+ > Li+, т. е. проводимость растет с увеличением размера иона; выпадает из этого ряда только Cs+. Неполное блокирование этакриновой кислотой Сl−-каналов не позволяет детально проанализировать ряд селективности плазмалеммы для двухвалентных катионов, который имеет вид: Cа2+ > Mg2+ > Sr2+ > Ba2+. На реконструированном одиночном Са2+-канале ряд селективности для двухвалентных катионов несколько отличен от такового на плазмалемме клеток харовой водоросли: (Ba2+ > Sr2+ > Са2+ > Mg2+). В опытах с перфузированными клетками с удаленным тонопластом
показано, что для функционирования потенциалзависимых кальциевых
каналов необходимо присутствие внутри клетки Mg2+ и АТФ. Более того, проведенные эксперименты по действию ингибитора цАМФ фосфодиэстеразы (теофилина), действие самого цАМФ, а также влияние на активацию каналов ингибитора кальмодулина (хлорпромазина) и активатора аденилатциклазы (форсколина) подтверждают существование цАМФ –зависимого регулирования функционального состояния Са2+-каналов растительной клетки. Что касается калиевых каналов, то к настоящему времени имеется уже достаточно экспериментальных данных, описывающих их транспортные свойства: механизм движения в канале, потенциалзависимость, селективность, блокирование и др. Коротко остановимся на каждом из перечисленных показателей полученных, как правило, в электрофизиологическом эксперименте. При помещении клетки в экспериментальный раствор, содержащий 5·10–3 М ионов K+, потенциал снижался от –60 до –80 мВ (–70 ± 5), чтоот величины напряжения на плазмалемме и времени выделялись три состояния: максимум выходящего тока, стационарное состояние после завершения инактивации при деполяризации и стационарное состояние по окончании уменьшения тока (деактивация) при гиперполяризации. Можно предположить, что каждое состояние обусловлено функционированием разных типов каналов. В частности, при деполяризации плазмалеммы ток возрастал при постоянном напряжении. Это происходит вследствие открывания (активации) каналов. Последующий спад тока с выходом на стационарный уровень – результат закрывания (инактивации) каналов. Причем в максимуме тока проводимость реализуется каналами обоих типов, а во время стационарного режима – преимущественно одного. Условно эти типы каналов были обозначены Д-каналы, открывающиеся при деполяризации, Г-каналы – при гиперполяризации; в последнее время их стали называть наружу- и внутрь- выпрямляющими соответственно. Смещение потенциала нулевого тока для обоих семейств кривых на десятикратное варьирование концентраций ионов K+ в диапазоне 0,5–10 мМ составляло 50–55 мВ, т. е. как при де-, так и при гиперполяризации мембраны ее проводимость оставалась почти чисто калиевой.
Впоследствии на основании анализа процессов активации – деактивации Д-каналов были идентифицированы два их типа – ДI и ДII.
Определения роста, развития и онтогенеза растений. Этапы онтогенеза.
Рост растений – сложный процесс, в его основе лежат такие фундаментальные явления как ритмичность, полярность, дифференциация, раздражимость, корреляция. Все эти процессы являются общими для онтогенеза всех живых организмов.
Онтогенез – индивидуальное развитие организма от зиготы (или вегетативного зачатка) до природной смерти.
Благодаря активной деятельности меристем и фотосинтетической активности листьев зеленое растение приобретает ряд тех своеобразных черт, которые характеризуют его рост. Последний в процессе онтогенеза растения наблюдается на протяжении основных этапов его жизненного цикла.
Этапность онтогенеза – это морфологическое и функциональное разветвление онтогенеза, которое проявляется в изменении характера роста, дифференциации и функциональной активности организма. Таких этапов выделяют пять: эмбриональный, ювенильный, спелость, размножение и старость. Эмбриональный этап у растений, размножающихся семенами – это период формирования зародыша и семян от оплодотворения яйцеклетки до начала прорастания семян; для вегетативного размножающихся растений – период формирования почек в органах вегетативного размножения от возникновения почки до начала ее прорастания. В это время ростовые процессы находятся в скрытой фазе или фазе подготовки. Происходит синтез основных метаболитов, ядра, образование ростовых гормонов. Ювенильный этап – период заложения, роста и развития вегетативных органов от прорастания семян до появления способности к образованию репродуктивных органов. В начале ювенильного периода растения не переходят к образованию репродуктивных органов даже в оптимальных условиях; потом они постепенно приобретают способность к репродукции, т. е. это этап доминирования ростовых процессов. В это же время растение и его органы активно увеличиваются в размерах, происходит новообразование отдельных элементов растения (клеток, тканей, органов). Многие авторы отмечают взаимодействие фитогормонов в период интенсивного роста растительного организма.
Спелость (зрелость) – период цветения в семенных или репродукции в вегетативно размножаемых растениях, от появления первичных зачатков репродуктивных органов до формирования бутонов, цветков, клубней, луковиц и других органов, а также образование новых зародышей. В этот период процесс роста сопряжен с элементами генеративного развития. Совместно с ростом вегетативных частей растения происходит детерминация генеративных органов – приобретение клеткой, тканью, органом или организмом состояния готовности к реализации определенных наследственных свойств. Детерминация развития характеризуется готовностью к развитию по определенному типу. Другими словами, растение переходит от этапа вегетативного размножения к этапу генеративного, т. е. к этапу спелости. Следующий этап – размножение – это период плодоношения, роста, развития и поспевания плодов и семян в растениях, размножающихся семенами; или поспевание клубней, луковиц и др. органов у вегетативно размножающихся растений. Во время этого этапа доминируют процессы роста генеративных органов, увеличение размеров семян или других органов. В этот период совместно с гормонами выявляются и доминируют природные ингибиторы. Последний этап – старость – период от полного прекращения плодоношения до природного отмирания растений. Рост в этом случае происходит очень редко (поросль из глубоко спящих почек, жирующие побеги). Замедляют старение факторы, стимулирующие синтез РНК и белков, и в первую очередь это цитокинины, а у некоторых растений также гиббереллины. Необходимо отметить, что этапы онтогенеза – это неизолированные периоды развития, а взаимопереходящие одна в другую фазы, в основе которых лежат медленно идущие возрастные изменения, т. е. изменения организма и его отдельных частей, обусловленные возрастом, и идущие на протяжении всей жизни растений. Возрастные изменения клеток, тканей и органов зависят от присущего им роста, общего возраста организма, а также от характера взаимосвязей с другими частями растения. Отметим, что в онтогенезе наблюдаются этапы, как интенсивного роста, так и его замедление (покой).
Этилен
Продуцируется в ответ на стресс и повреждение.
 Вызывает опадание листьев, обеспечивает адаптацию метаболизма к стрессу, созревание фруктов.
Вызывает опадание листьев, обеспечивает адаптацию метаболизма к стрессу, созревание фруктов.
Уменьшение уровня ауксина и повышение уровня этилена приводит к образованию зоны отделения в основании черешка и опадению листьев.(СН2=СН) – гормон старения (гормональный фактор газоподобного типа). Давно известно, что одно гнилое яблоко в бочке вызывает порчу всех остальных. Как оказалось, в гнилом яблоке вырабатыва-ется летучее вещество – этилен, вызывающее разрушительное действие в здоровых плодах.
Впервые физиологический эффект этилена на растения был описан Д. А. Нелюбовым в 1901 г. Он выявил, что в этиолированных проростках (гороха) этилен вызывает тройную реакцию стебля: ингибирование растяжения, утолщение и горизонтальную ориентацию. В 20-х гг. было показано, что этилен способен ускорять спелость плодов и регулировать процесс старения у растений. Тот факт, что действие этилена можно снять повышенной концентрацией СО2 в окружающей среде, лежит в основе практического приема хранения яблок и других плодов. Этилен вызывает образование апикального изгиба во многих этиолированных проростках; действие света на выпрямление изгиба связано с тем, что свет ингибирует образование этилена. Этилен может также влиять на геотропизм и другие опосредованные ауксином реакции (например, подавление роста боковых почек). Этилен тормозит полярный транспорт ауксина, усиливает процессы старения, опадения листьев и плодов, устраняет апикальное доминирование, а также ускоряет поспевание плодов. Образуется этилен из серосодержащей аминокислоты метионина, который превращается в S-аденозилметионин. Затем с помощью фермента АЦК-синтазы синтезируются 1-амино-циклопропан-1-карбоновая кислота (АЦК) и 5/-метилтиоаденазин. Последний этап синтеза этилена происходит в присутствии кислорода и катализируется ферментом АЦК-оксидазой. Постоянное пополнение L-метионина происходит в результате превращения 5/-метилтиоаденазина через последовательное образование 5/-метилтиорибозы, 5/-метилрибозо-1-фосфат и α-кето-γ-метилтиомасляной кислоты (цикл Янга)
Абсцизовая кислота (АБК) – природный гормональный ингибитор роста терпеноидной природы. Абсцизовая кислота (AБК) замедляет рост
Два основных эффекта:- Поддерживает покой семян - Закрывает устьица при засухе
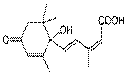 АБК. Изопреноидный путь синтеза в пластидах. Название АБК получила в связи с ее способностью вызывать опадение черешков, листьев, завязей и плодов (от английского abscission – отнятие, ампутация).В 1955 г. из старых листьев фасоли и некоторых древесных растений выделили вещества, которые ускоряли опадение листьев. Была вы-сказана догадка, что опадение листьев регулируется не только эндогенными ауксинам и этиленам, но и другими веществами, которые образуются в спелых листьях. Скоро (1961 г.) подобное вещество было найдено и в молодых плодах хлопчатника. Последующие исследования показали, что веществом, ускоряющим опадение, во всех случаях была АБК. Это соединение стимулировало опадение листьев у березы, клена и др. Молекулярная структура АБК была установлена в 1963 г. На основе ее структуры сделано предположение, что ее синтез, как и синтез гиббереллинов, идет по пути образования изопреноидов. Изопентенилпирофосфат является тем структурным элементом, из которого образуется АБК. Пути биосинтеза, распада и связывания АБК, выявленные с помощью мутантов, выглядят следующим образом.Катаболизм АБК идет по пути коньюгации и образования глюкозидов, а также по пути деградации с образованием соответствующих кислот (фазеевой, оксиабсцизовой и др. кислот).АБК синтезируется главным образом в листьях, а также в корневом чехлике. Перемещение АБК в растениях наблюдается как в базипетальном, так и в акропетальном направлениях в составе ксилемного и флоэмного соков. Как мы уже в начале отметили, в большинстве случаев, АБК тормозит рост растений. Этот гормон в ряде случаев выступает как антагонист ИУК, цитокинина и гиббереллинов. АБК ингибирует прорастание семян и рост почек, содействует опадению листьев, которое связано с их старением. АБК ускоряет распад нуклеиновых кислот, белков и хлорофилла. В некоторых случаях АБК является активатором: она стимулирует развитие партенокарпии у розы, удлинение гипокотеля огурца, образование корней у черенков фасоли. Активные формы кислорода (АФК) - Супероксидный анион •O2- - Перекись водорода H2O2- Синглетный кислород 1O2 - Гидроксильный радикал •OH
АБК. Изопреноидный путь синтеза в пластидах. Название АБК получила в связи с ее способностью вызывать опадение черешков, листьев, завязей и плодов (от английского abscission – отнятие, ампутация).В 1955 г. из старых листьев фасоли и некоторых древесных растений выделили вещества, которые ускоряли опадение листьев. Была вы-сказана догадка, что опадение листьев регулируется не только эндогенными ауксинам и этиленам, но и другими веществами, которые образуются в спелых листьях. Скоро (1961 г.) подобное вещество было найдено и в молодых плодах хлопчатника. Последующие исследования показали, что веществом, ускоряющим опадение, во всех случаях была АБК. Это соединение стимулировало опадение листьев у березы, клена и др. Молекулярная структура АБК была установлена в 1963 г. На основе ее структуры сделано предположение, что ее синтез, как и синтез гиббереллинов, идет по пути образования изопреноидов. Изопентенилпирофосфат является тем структурным элементом, из которого образуется АБК. Пути биосинтеза, распада и связывания АБК, выявленные с помощью мутантов, выглядят следующим образом.Катаболизм АБК идет по пути коньюгации и образования глюкозидов, а также по пути деградации с образованием соответствующих кислот (фазеевой, оксиабсцизовой и др. кислот).АБК синтезируется главным образом в листьях, а также в корневом чехлике. Перемещение АБК в растениях наблюдается как в базипетальном, так и в акропетальном направлениях в составе ксилемного и флоэмного соков. Как мы уже в начале отметили, в большинстве случаев, АБК тормозит рост растений. Этот гормон в ряде случаев выступает как антагонист ИУК, цитокинина и гиббереллинов. АБК ингибирует прорастание семян и рост почек, содействует опадению листьев, которое связано с их старением. АБК ускоряет распад нуклеиновых кислот, белков и хлорофилла. В некоторых случаях АБК является активатором: она стимулирует развитие партенокарпии у розы, удлинение гипокотеля огурца, образование корней у черенков фасоли. Активные формы кислорода (АФК) - Супероксидный анион •O2- - Перекись водорода H2O2- Синглетный кислород 1O2 - Гидроксильный радикал •OH
Классические и новые разделы физиологии растений. Их предмет и задачи. Важность физиологии и биотехнологии растений для общества и устойчивого развития человечества.
Основные классические фундаментальные разделы ФЗР: - фотосинтез - дыхание - водобмен- минеральное питание - рост и развитие - физиология стресса
В последние годы появились новые фундаментальные разделы ФЗР, такие как
- сигнальная трансдукция и передача информации - внутриклеточный транспорт - иммунитет растений - геномика (раздел молекулярной генетики, посвящённый изучению генома и генов живых организмов.) - протеомика (изучает белки и их взаимодействия в живых организмах.
Идентификация, характеристика и количественный учет ВСЕХ белков метаболического пути, органеллы, клетки, ткани, органа и всего организма.)
- феномика
- метаболомика (это «систематическое изучение уникальных химических „отпечатков пальцев“ специфичных для процессов, протекающих в живых клетках» — конкретнее, изучение их низкомолекулярных метаболических профилей. Метаболом представляет собой совокупность всех метаболитов, являющихся конечным продуктом обмена веществ в клетке, ткани, органе или организме. В то время как данные об экспрессии мРНК генов и данные протеомного анализа не раскрывают полностью всего того, что может происходить в клетке, метаболические профили могут дать мгновенный снимок физиологических процессов в клетке. Одна из задач системной биологии и функциональной геномики — интегрирование данных протеомики, транскриптомики и метаболической информации для получения более целостного представления о живых организмах. Метаболомика — это научное изучение химических процессов, в которые вовлечены метаболиты.)
- иономика
Новые прикладные отрасли ФЗР: - молекулярная селекция - генетическая модификация
- молекулярные основы урожайности - устойчивое развитие и устойчивое сельское хозяйство
- получение биотоплива – агрофеномика - молекулярное сельское хозяйство - биоинформатика растений - системная биология растений и т.д.