

Кормораздатчик мобильный электрифицированный: схема и процесс работы устройства...

Механическое удерживание земляных масс: Механическое удерживание земляных масс на склоне обеспечивают контрфорсными сооружениями различных конструкций...
Кормораздатчик мобильный электрифицированный: схема и процесс работы устройства...
Механическое удерживание земляных масс: Механическое удерживание земляных масс на склоне обеспечивают контрфорсными сооружениями различных конструкций...
Топ:
Характеристика АТП и сварочно-жестяницкого участка: Транспорт в настоящее время является одной из важнейших отраслей народного хозяйства...
Устройство и оснащение процедурного кабинета: Решающая роль в обеспечении правильного лечения пациентов отводится процедурной медсестре...
Теоретическая значимость работы: Описание теоретической значимости (ценности) результатов исследования должно присутствовать во введении...
Интересное:
Аура как энергетическое поле: многослойную ауру человека можно представить себе подобным...
Средства для ингаляционного наркоза: Наркоз наступает в результате вдыхания (ингаляции) средств, которое осуществляют или с помощью маски...
Принципы управления денежными потоками: одним из методов контроля за состоянием денежной наличности является...
Дисциплины:
|
из
5.00
|
Заказать работу |
|
|
|
|
Изотоническое сокращение - это сокращение мышцы без нагрузки. Его можно получить в том случае, если мышцу закрепить с одной стороны, и раздражать её. В этом случае длина мышцы будет изменяться, а тонус окажется постоянным.
И при чисто изометрическом, и при чисто изотоническом сокращении работа совершаться не будет (нет перемещения груза). Поэтому в нормальных условиях скелетные мышцы сокращаются в смешанном (или ауксотоническом) режиме, где есть элементы и изометрического, и изотонического сокращения.
14. Закон силы и закон «все или ничего» в ответах возбудимых структур на раздражение.
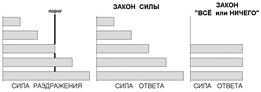
Закон силы: чем больше сила раздражителя, тем больше величина ответной реакции. В соответствии с этим законом функционирует скелетная мышца. Амплитуда ее сокращений постепенно увеличивается с увеличением силы раздражителя вплоть до достижения максимальных значений. Это обусловлено тем, что скелетная мышца состоит из множества мышечных волокон, имеющих различную возбудимость. На пороговые раздражители отвечают только волокна, имеющие самую высокую возбудимость, амплитуда мышечного сокращения при этом минимальна. Увеличение силы раздражителя приводит к постепенному вовлечению волокон, имеющих меньшую возбудимость, поэтому амплитуда сокращения мышцы усиливается. Когда в реакции участвуют все мышечные волокна данной мышцы, дальнейшее повышение силы раздражителя не приводит к увеличению амплитуды сокращения.
Закон «все или ничего»: подпороговые раздражители не вызывают ответной реакции («ничего»), на пороговые раздражители возникает максимальная ответная реакция («все»). По закону «все или ничего» сокращаются сердечная мышца и одиночное мышечное волокно. Закон «все или ничего» не абсолютен. Вопервых, на раздражители подпороговой силы не возникает видимой ответной реакции, но в ткани происходят изменения мембранного потенциала покоя в виде возникновения местного возбуждения (локального ответа). Во-вторых, сердечная мышца, растянутая кровью, реагирует по закону «все или ничего», но амплитуда ее сокращения будет больше по сравнению с таковой при сокращении нерастянутой сердечной мышцы.
|
|
«ВСЁ ИЛИ НИЧЕГО» закон — эмпирически установленное соотношение между силой действующего раздражителя и величиной ответной реакции возбудимой структуры. Согласно этому закону возбудимая ткань дает максимальную ответную реакцию («все») при любой силе надпорогового раздражения (включая и пороговое) или не дает никакого ответа («ничего») при подпороговом раздражении (см. Возбудимость).
Впервые подобные отношения между силой раздражения и эффектом наблюдал Боудич (H. Bowditch, 1871). Постепенно увеличивая силу одиночных раздражений, он выявил, что при подпороговой силе раздражения мышца сердца не сокращается, а при пороговой силе раздражения сокращение сердца имеет максимальную амплитуду. При дальнейшем увеличении силы раздражения амплитуда сокращения не увеличивается.
Исходя из предположения о хим. природе процесса возбуждения и его взрывном характере, Готч (F. Gotch, 1902) пришел к выводу о применимости закона «все или ничего» к нервной ткани. Сравнивая токи действия нерва при слабых и сильных раздражениях, он обнаружил, что длительность и форма токов остаются постоянными. Этот феномен он объяснил независимостью нервного импульса от природы и силы вызвавшего его раздражителя (в случае если раздражитель достиг порогового значения). В дальнейшем эта закономерность была экспериментально установлена и на одиночных поперечнополосатых мышечных волокнах.
Проявление закона «все или ничего» в возбудимых тканях физиологи связывали с качественной характеристикой возникновения процесса возбуждения, предполагая, что «ничего» означает полное отсутствие возбуждения на подпороговое раздражение. При достижении силы действующего раздражителя пороговой величины возбудимая ткань реагирует с полным использованием всех своих потенциальных возможностей («все»), по типу взрыва пороха (см. Триггерные механизмы).
|
|
Закон «все или ничего» долгое время рассматривался как общий принцип реагирования возбудимых тканей. Однако экспериментально установлена относительность этого закона. Оказалось, что мышца сердца не при всех условиях отвечает одинаково на раздражители разной силы: «все» изменяется в зависимости от температуры, исходного растяжения мышцы, степени ее утомления.
Исследованиями возбудимых структур с применением тонких электрофизиологических методов исследования (см. Электрофизиология) также установлена и относительная независимость эффекта от силы действующего раздражителя. В одиночном нервном волокне подпороговое раздражение вызывает местное, нераспространяющееся изменение потенциала; следовательно, нельзя говорить, что раздражение не дает «ничего». Кроме того, уровень «все» в свою очередь также изменяется с изменением функционального состояния нервного волокна.
15. ЭЭГ. Формирование суммарного электрического ответа нейронов.
Большое значение для понимания природы электрических колебаний головного мозга имели исследования нейрофизиологии отдельных нейронов с помощью метода микроэлектродов, выявившие те структурные субъединицы и механизмы, из которых слагается суммарная ЭЭГ (Костюк П.Г., Шаповалов А.И., 1964, Eccles J., 1964).
ЭЭГ представляет собой сложный колебательный электрический процесс, который может быть зарегистрирован при расположении электродов на мозге или на поверхности скальпа, и является результатом электрической суммации и фильтрации элементарных процессов, протекающих в нейронах головного мозга.
Многочисленные исследования показывают, что электрические потенциалы отдельных нейронов головного мозга связаны тесной и достаточно точной количественной зависимостью с информационными процессами. Для того чтобы нейрон генерировал потенциал действия, передающий сообщение другим нейронам или эффекторным органам, необходимо, чтобы собственное его возбуждение достигло определенной пороговой величины.
|
|
Уровень возбуждения нейрона определяется суммой возбуждающих и тормозных воздействий, оказываемых на него в данный момент через синапсы. Если сумма возбуждающих воздействий больше суммы тормозных на величину, превышающую пороговый уровень, нейрон генерирует нервный импульс, распространяющийся затем по аксону. Описанным тормозным и возбуждающим процессам в нейроне и его отростках соответствуют определенной формы электрические потенциалы.
Мембрана -- оболочка нейрона -- обладает электрическим сопротивлением. За счет энергии обмена веществ концентрация положительных ионов в экстраклеточной жидкости поддерживается на более высоком уровне, чем внутри нейрона. В результате существует разность потенциалов, которую можно измерить, введя один микроэлектрод внутрь клетки, а второй расположив экстраклеточно. Эта разность потенциалов называется потенциалом покоя нервной клетки и составляет около 60-70 мВ, причем внутренняя среда заряжена отрицательно относительно экстраклеточного пространства. Наличие разности потенциалов между внутриклеточной и внеклеточной средой носит название поляризации мембраны нейрона.
Увеличение разности потенциалов называется соответственно гиперполяризацией, а уменьшение -- деполяризацией. Наличие потенциала покоя является необходимым условием нормального функционирования нейрона и генерирования им электрической активности. При прекращении обмена веществ или снижении его ниже допустимого уровня различия концентраций заряженных ионов по обе стороны мембраны сглаживаются, с чем связано прекращение электрической активности в случае клинической или биологической смерти мозга. Потенциал покоя является тем исходным уровнем, на котором происходят изменения, связанные с процессами возбуждения и торможения, -- спайковая импульсная активность и градуальные более медленные изменения потенциала. Спайковая активность (от англ. spike-- острие) характерна для тел и аксонов нервных клеток и связана с бездекрементной передачей возбуждения от одной нервной клетки к другой, от рецепторов к центральным отделам нервной системы или от центральной нервной системы к исполнительным органам. Спайковые потенциалы возникают в момент достижения мембраной нейрона некоторого критического уровня деполяризации, при котором наступает электрический пробой мембраны и начинается самоподдерживающийся процесс распространения возбуждения в нервном волокне.
|
|
При внутриклеточной регистрации спайк имеет вид высокоамплитудного, короткого, быстрого положительного пика.
Характерными особенностями спайков являются их высокая амплитуда (порядка 50-125 мВ), небольшая длительность (порядка 1-2 мс), приуроченность их возникновения к достаточно строго ограниченному электрическому состоянию мембраны нейрона (критический уровень деполяризации) и относительная стабильность амплитуды спайка для данногонейрона (закон все или ничего).
Градуальные электрические реакции присущи в основном дендритам в соме нейрона и представляют собой постсинаптические потенциалы (ПСП), возникающие в ответ на приход к нейрону спайковых потенциалов по афферентным путям от других нервных клеток. В зависимости от активности возбуждающих или тормозящих синапсов соответственно различают возбуждающие постсинаптические потенциалы (ВПСП) и тормозные постсинаптические потенциалы (ТПСП).
16. Роль синаптических и спайковых потенциалов в генерации ЭЭГ.
17. Частотный спектр ЭЭГ. Связь частотной структуры ЭЭГ с функциональным состоянием мозга.
Выделяют 5 главных ритмов, имеющих различные частотный диапазон, амплитуду и функциональное значение.
1. Дельта-ритм (0,5-4 Гц). Возникает при естественном и наркотическом сне, а так же наблюдается при регистрации ЭЭГ от участков коры, граничащих с областью, пораженных опухолью. У здорового взрослого человека практически отсутствует.
2. Тэта-ритм (5-7 Гц). Наиболее выражен в гиппокампе. Связан с поисковым поведением, усиливается при эмоциональном напряжении. Его иногда так и называют стресс-ритм. Сопровождается переживанием как положительных, так и отрицательных эмоций.
3. Альфа-ритм (8-13 Гц). Основной человеческий ритм, наблюдающийся в состоянии бодрствования, медитации и длительной монотонной деятельности. Наиболее выражен в затылочных областях головного мозга. Обычно эти колебания испытывают определенную модуляцию, так называемое «веретено альфа-ритма» – постепенное увеличение, а затем уменьшение амплитуды колебания альфа-ритма. Длительность веретен доля секунды-несколько секунд. «Десинхронизация» альфа-ритма – реакция пробуждения на раздражитель.
4. Мю-ритм — по частотно-амплитудным характеристикам сходен с альфа-ритмом, но преобладает в передних отделах коры больших полушарий.
|
|
5. Бета-ритм (15-35 Гц) – локализуется в предцентральной и фронтальной коре. Наблюдается при умственной активности у взрослых, причем значимое усиление высокочастотной активности наблюдается при умственной деятельности, включающей элементы новизны, в то время как стереотипные, повторяющиеся умственные операции сопровождаются ее снижением. Переход к состоянию напряжения сопровождается появлением бета-активности.
6. Гамма-ритм (выше 35 Гц). В предцентральной, теменной, височной зонах коры. Наблюдается при решении задач, требующих максимального сосредоточения.
Клинический и статический методы изучения ЭЭГ. С момента возникновения выделились и продолжают существовать как относительно самостоятельные два подхода к анализу ЭЭГ: визуальный (клинический) и статистический.
Визуальной (клинический) анализ ЭЭГ используется, как правило, в диагностических целях. Электрофизиолог, опираясь на определенные способы такого анализа ЭЭГ, решает следуюшие вопросы: соответствует ли ЭЭГ общепринятым стандартам нормы; если нет, то какова степень отклонения от нормы, обнаруживаются ли у пациента признаки очагового поражения мозга и какова локализация очага поражения. Клинический анализ ЭЭГ всегда строго индивидуален и носит преимущественно качественный характер. Несмотря на то, что существуют общепринятые в клинике приемы описания ЭЭГ, клиническая интерпретация ЭЭГ в большей степени зависит от опыта электрофизиолога, его умения "читать" электроэнцефалограмму, выделяя в ней скрытые и нередко очень вариативные патологические признаки.
18. Методики регистрации ЭЭГ.
Регистрация ЭЭГ представляет собой непрерывную запись величин разности потенциалов между двумя точками мозга. Отведение потенциалов производят с помощью специальных контактных электродов, приложенных к поверхности кожи на голове. Электроды должны обладать малым переходным (электрод-кожа) сопротивлением, минимальной поляризацией и антикоррозийными свойствами. Для уменьшения сопротивления кожи, контактирующей с электродом, ее обрабатывают жирорастворяющими веществами (например, эфиром). На голове электроды закрепляют с помощью специальных эластичных шлемов. К электродам присоединяют проводники, обладающие изоляцией и служащие для подключения электродов на вход электроэнцефалографа.
Во избежание артефактов регистрацию проводят в специальной экранированной камере, предохраняющей объект от воздействия внешних электрических и магнитных полей. Испытуемого усаживают в удобное кресло. Одевают на него шлем-сетку и определяют точки на голове, где должны быть расположены электроды. На этих участках раздвигают волосы, протирают кожу для обезжиривания эфиром и на кусочки марли, смоченные физиологическим раствором, накладывают электроды, прижимая их эластичными шнурами шлема. Парные активные электроды должны располагаться на лобных, височных, теменных и затылочных областях полушарий.
Индифферентный электрод, проложенный смоченным в физиологическом растворе кусочком марли, располагается на мочке уха, которая также обезжиривается эфиром.
Электрод заземления через кусочек марли помещается на обезжиренное запястье испытуемого.
Испытуемого просят расслабиться и закрыть глаза. Начать запись с регистрации калибровочного сигнала. Далее ведут запись ЭЭГ испытуемого. Спустя несколько минут наблюдают появление на ЭЭГ α -ритма. Просят испытуемого открыть глаза и наблюдают депрессию α -ритма.
После этого просят испытуемого снова закрыть глаза и расслабиться. Как только восстановится исходный α -ритм испытуемому предлагают умственную нагрузку. Например, произвести арифметические действия разного уровня сложности (умножение, вычитание, сложение, деление чисел). При этом продолжают записывать ЭЭГ, отмечая при этом угнетение α -ритма и появление β -волн, особенно в лобном отведении.
После восстановления исходного α -ритма предъявите испытуемому эмоционально значимую для него информацию (слова, текст). При записи ЭЭГ отметьте опять-таки активацию (десинхронизацию) ЭЭГ.
Подсчитайте частоту и амплитуду зафиксированных ритмов ЭЭГ как в покое, так и при умственном и эмоциональном напряжении.
Сделайте соответствующие выводы. Вклейте в тетрадь образцы записей ЭЭГ.
Вегетативные рефлексы
Спинальные вегетативные рефлексы — это замыкающиеся на уровне спинного мозга регуляторные реакции, осуществляющиеся при участии вегетативной нервной системы. В норме эти рефлексы находятся под жестким контролем со стороны вышележащих отделов центральной нервной системы. Перерыв спинного мозга приводит к развитию спинального шока — временному исчезновению соматических и вегетативных рефлексов. Чем выше уровень организации животного, тем больше длительность спинального шока: от нескольких минут у лягушки до нескольких месяцев у человека. Со временем явления спинального шока исчезают благодаря тому, что спинной мозг, отделенный от головного, начинает выполнять ряд регуляторных функций: восстанавливается регуляция сосудистого тонуса, рефлексы опорожнения мочевого пузыря и толстого кишечника, половые рефлексы.
Регулировать вегетативные функции за счет влияния на активность преганглионарных симпатических и парасимпатических нейронов могут практически все отделы центральной нервной системы, вплоть до коры больших полушарий. Наиболее тесно вегетативная нервная система связана со структурами, расположенными в стволе мозга и в гипоталамусе.
Методика проведения электроэнцефалографии Техника проведения
В обычной практике ЭЭГ отводят с помощью электродов, расположенных на интактных покровах головы. Электрические потенциалы усиливают и регистрируют. В электроэнцефалографах предусмотрено 16-24 и более идентичных усилительно-регистрирующих блоков (каналов), позволяющих одномоментно записывать электрическую активность от соответствующего количества пар электродов, установленных на голове пациента. Современные электроэнцефалографы создают на базе компьютеров. Усиленные потенциалы преобразуют в цифровую форму; непрерывная регистрация ЭЭГ отображается на мониторе и одновременно записывается на диск. После обработки ЭЭГ может быть распечатана на бумаге.
Электроды, отводящие потенциалы, представляют собой металлические пластины или стержни различной формы с диаметром контактной поверхности 0,5-1 см. Электрические потенциалы подаются на входную коробку электроэнцефалографа, имеющую 20-40 и более пронумерованных контактных гнёзд, с помощью которых к аппарату можно подсоединить соответствующее количество электродов. В современных электроэнцефалографах входная коробка объединяет коммутатор электродов, усилитель и аналогоцифровой преобразователь ЭЭГ. Из входной коробки преобразованный сигнал ЭЭГ подают в компьютер, с помощью которого производят управление функциями прибора, регистрацию и обработку ЭЭГ.
ЭЭГ регистрирует разность потенциалов между двумя точками головы. Соответственно на каждый канал электроэнцефалографа подают напряжения, отведённые двумя электродами: одно на «вход 1», другое на «вход 2» канала усиления. Многоконтактный коммутатор отведений ЭЭГ позволяет коммутировать электроды по каждому каналу в нужной комбинации. Установив, например, на каком-либо канале соответствие затылочного электрода гнезду входной коробки «1», а височного - гнезду коробки «5», получают тем самым возможность регистрировать по этому каналу разность потенциалов между соответствующими электродами. Перед началом работы исследователь набирает с помощью соответствующих программ, несколько схем отведений., которые и используют при анализе полученных записей. Для задания полосы пропускания усилителя используют аналоговые и цифровые фильтры высокой и низкой частоты. Стандартная полоса пропускания при записи ЭЭГ - 0,5-70 Гц.
|
|
|

Поперечные профили набережных и береговой полосы: На городских территориях берегоукрепление проектируют с учетом технических и экономических требований, но особое значение придают эстетическим...

Эмиссия газов от очистных сооружений канализации: В последние годы внимание мирового сообщества сосредоточено на экологических проблемах...

Таксономические единицы (категории) растений: Каждая система классификации состоит из определённых соподчиненных друг другу...

Автоматическое растормаживание колес: Тормозные устройства колес предназначены для уменьшения длины пробега и улучшения маневрирования ВС при...
© cyberpedia.su 2017-2024 - Не является автором материалов. Исключительное право сохранено за автором текста.
Если вы не хотите, чтобы данный материал был у нас на сайте, перейдите по ссылке: Нарушение авторских прав. Мы поможем в написании вашей работы!