Симпатического и парасимпатического отделов вегетативнойнервнойсистемы.Характеристикаадренергическихи
Холинергическихрецепторов.
Влияниесимпатическогоотдела:
На сердце -- повышает частоту и силу сокращений сердца.Наартерии-- сужает артерии.
На кишечник -- угнетает перистальтику кишечника и выработку пищеварительныхферментов.
Наслюнныежелезы--угнетаетслюноотделение.
Намочевойпузырь--расслабляетмочевойпузырь.
На бронхи и дыхание -- расширяет бронхи и бронхиолы, усиливает вентиляцию лёгких.Назрачок -- расширяетзрачки.
Влияниепарасимпатическогоотдела:
На сердце -- уменьшает частоту и силу сокращений сердца.Наартерии-- расслабляетартерии.
На кишечник -- усиливает перистальтику кишечника и стимулирует выработкупищеварительныхферментов.
На слюнные железы -- стимулирует слюноотделение.Намочевойпузырь--сокращаетмочевойпузырь.
На бронхи и дыхание -- сужает бронхи и бронхиолы, уменьшает вентиляцию лёгкихНазрачок --сужает зрачки
Холинергическими рецепторами называют рецепторы специфичные для ацетилхолина.Ацетилхолин — это вещество, которое в полной мере отвечает требованиям, позволяющимсчитать его химическим посредником передачи возбуждения: локализация ацетилхолина иучаствующеговегосинтезеферментавпресинаптическойтерминалиисоответствующей
нервной клетке, его высвобождение при возбуждении; выделение при стимуляции нервныхокончаний; наличие в постсинаптической мембране высокочувствительных химическихструктур(рецепторов),атакжеферментативныхсистемдля инактивациивещества.
Возбуждающее действие: глубокие слои коры мозга позвоночных, новая кора, ядраталамуса, ствол мозга, нервно-мышечные синапсы, нейроны вегетативных ганглиев,мотонейроныспинного мозга
Тормозящее действие: полиморфные нейроны глубоких слоев коры (М-холинорецепторы),стволмозга, хвостатоеядро, блуждающийнервсердца позвоночных
Рецепторы чувствительные к адреналину и норадреналину называют адренергическими.Адренорецепторы делят на α-адренорецепторы и β-адренорецепторы. Действие веществасчитаетсяа-адренергическим,еслиприеговоспроизведенииэффективностьубываетв
последовательности норадреналин-адреналин-изопротеренол, а также избирательноблокируется B-адреноблокаторами. Если же воспроизведение действия будет происходитьв обратной последовательности: изопротеренол-адреналин-норадреналин — и этот эффектпредотвращаетсяспецифическимиблокаторами,товэтомслучае имеетместо В-
адренергическийэффект.
В большинстве висцеральных органов, реагирующих на катехоламины, находятся оба видаадренорецепторов и результаты их возбуждения бывают, как правило, противоположными.Например, в гладкой мышце артерий скелетных мышц содержатся а- и В-адренорецепторы.Возбуждениеа-адренорецепторовприводитксужениюартериол, авозбуждение B-
адренорецепторов — к их расширению. В кишке также имеются оба вида адренорецепторов,однаковоздействиенатеидругиевызываетторможениеактивностигладкоймышцы.В
обычныхжизненныхусловияхреакцияоргананаадреналининорадреналин,
выделяющиеся нервными окончаниями или приносимые кровью, зависит от преобладанияа-илиВ-адренергического действия.
31. Методыизученияфункцийцентральнойнервнойсистемы.
СуществуютследующиеметодыисследованияфункцийЦНС:
1. Метод перерезок ствола мозга на различных уровнях. Например, между продолговатым испинныммозгом.
2. Метод экстирпации (удаления) или разрушения участков мозга. Например, удалениемозжечка.
3. Методраздраженияразличныхотделовицентровмозга.
4. Анатомо-клинический метод. Клинические наблюдения за изменениями функций ЦНС припораженииеекаких-либоотделовспоследующимипатологоанатомическимисследованием.
5. Электрофизиологическиеметоды:
· Электроэнцефалография – регистрация биопотенциалов мозга с поверхности кожи черепа.МетодикаразработанаивнедренавклиникуГ. Бергером.
· Регистрация биопотенциалов различных нервных центров: используется вместе состереотаксической техникой при которой электроды с помощью микроманипулятороввводятвстрого определенное ядро.
· Метод вызванных потенциалов, регистрация электрической активности участков мозгаприэлектрическомраздражениипериферическихрецепторовилидругихучастков.
6. Методвнутримозговоговведениявеществспомощью микроинофореза.
7. Хронорефлексометрия–определениевременирефлексов.
8. Методмоделирования.
32. Электрофизиологические методы исследования ЦНС(электроэнцефалография,вызванныепотенциалыит.д.)
Кэлектрофизиологическимметодамисследованияотносят:
* ЭРГ; Электроретинография (ЭРГ) — это регистрация электрического потенциала,возникающего в ответ на вспышку света. Этот ответ является результатомтранерстинальногопотокаионов,возникающегокакреакциянасветовойстимул.
* электроокулографию (ЭОГ); В основе этого метода лежит использование собственныхэлектрическихсвойствглазногояблока.Посвоейфизическойприродеоноявляется
диполем, в котором роговица относительно сетчатки электроположительна. Электрическаяось глазного яблока примерно совпадает с оптической осью и, следовательно, можетслужитьиндикатором направления взора.
* зрительные вызванные корковые потенциалы (или зрительные вызванные потенциалы(ЗВП)).
Электроэнцефалограмма – это криваярегистрациисуммарного потенциала постоянноменяющейся биоэлектрической активности значительной группы нервных клеток. В этусуммувходятсинаптическиепотенциалы иотчастипотенциалы действиянейронови
нервныхволокон.
Методвызванныхпотенциалов
Специфическая активность, связанная со стимулом, называется вызванным потенциалом. Учеловека–эторегистрацияколебанияэлектрическойактивности,возникающегонаЭЭГ
при однократном раздражении периферических рецепторов (зрительных, слуховых,тактильных). У животных раздражают также афферентные пути и центры переключенияафферентной импульсации.Амплитуда их обычно невелика, поэтому для эффективноговыделения вызванных потенциалов применяют прием компьютерного суммирования иусредненияучастковЭЭГ,котороезаписалосьприповторномпредъявлениистимула.
33. Ретикулярная формация ствола мозга, функциональнаяхарактеристика ее нисходящих влияний на рефлекторнуюдеятельностьспинного мозга.
Ретикулярная формация ствола мозга рассматривается как один из важных интегративныхаппаратовмозга.
К собственно интегративных функций ретикулярной формации относятся:контрольнадсостояниямиснаибодрствования
мышечный(фазныйитонический)контроль
обработка информационных сигналов окружающей и внутренней среды организма,которыепоступают по разным каналам
Нисходящее влияние. При раздражении ретикулярной формации заднего мозга (особенногигантоклеточной ядра продолговатого мозга и ретикулярного ядра моста, где принимаютначало ретикулоспинальному пути), возникает торможение всех спинальных двигательныхцентров(сгибательныхиразгибательных). Этоторможениеоченьглубокоеи
продолжительное. Такое положение в естественных условиях может наблюдаться приглубокомсне.
Наряду с диффузными тормозящими влияниями, при раздражении определенных участковретикулярной формации выявляется диффузное влияние, которое облегчает деятельностьспинальнойдвигательнойсистемы.
Ретикулярная формация играет важную роль в регуляции деятельности мышечных веретен,изменяячастотуразрядов,поступающиегамма-эфферентнымиволокнамикмышцам.
Такимобразоммодулируетсяобратнаяимпульсациявних.
34. Характеристика восходящих влияний ретикулярной формациинакоруголовногомозга.Участиеретикулярнойформациивформировании
целостнойдеятельностиорганизма.
Восходящий влияние. Исследования Н. W. Megoun, G. Moruzzi (1949) показали, чтораздражение ретикулярной формации (заднего, среднего и промежуточного мозга)сказывается на деятельности высших отделов головного мозга, в частности коры большогомозга, обеспечивая переход ее в активное состояние. Это положение подтверждаетсяданнымимногочисленнымиэкспериментальнымиисследованиямииклиническими
наблюдениями. Так, если животное находится в состоянии сна, то прямое раздражениеретикулярной формации (особенно варолиева моста) через введенные в эти структурыэлектроды вызывает поведенческую реакцию пробуждения животного. При этом на ЭЭГвозникает характерное изображение - изменение альфа-ритма бета-ритмом, т.е.фиксируетсяреакциядесинхронизацииилиактивизации.Указаннаяреакцияне
ограничивается определенным участком коры большого мозга, а охватывает большие еемассивы, т.е. носит генерализованный характер. При разрушении ретикулярной формациииливыключенииее восходящихсвязейскоройбольшого мозгаживотное впадает в
сноподобное состояние, не реагирует на световые и обонятельные раздражители,фактически не вступает в контакт с внешним миром. То есть конечный мозг прекращаетактивнофункционировать.
Таким образом, ретикулярная формация ствола головного мозга выполняет функциивосходящей активирующей системы мозга, которая поддерживает на высоком уровневозбудимостьнейроновкоры большого мозга.
35. Функции спинного мозга. Функциональная характеристикаспинальныхрефлексовипроводящихпутейспинногомозга.
Спинной мозг — наиболее древнее образование центральной нервной системы; он впервыепоявляетсяуланцетника.
Характерной чертой организации спинного мозга является периодичность его структуры вформесегментов, имеющихвходы ввиде заднихкорешков,клеточнуюмассунейронов
(серое вещество) и выходы в виде передних корешков. Морфологических границ междусегментамиспинного мозганесуществует, поэтомуделение насегменты является
функциональным и определяется зоной распределения в нем волокон заднего корешка изонойклеток,которыеобразуютвыходпереднихкорешков. Каждыйсегментчерезсвои
корешки иннервирует три метамера тела и получает информацию также от трех метамеровтела.Витоге перекрытиякаждыйметамертелаиннервируетсятремясегментамии
передаетсигналывтрисегментаспинногомозга.
Спинной мозг человека имеет два утолщения: шейное и поясничное — в них содержитсябольшеечислонейронов, чем востальныхегоучастках.
Спинному мозгу присущи две функции - рефлекторная и проводниковая. Как рефлекторныйцентрспинноймозгспособеносуществлятьсложные двигательныеивегетативные
рефлексы. Афферентными (чувствительными) путями он связан с рецепторами, аэфферентными-соскелетноймускулатуройивсемивнутреннимиорганами.
Проводниковая функция спинного мозга осуществляется с помощью нисходящих ивосходящихпроводящихпутей.
Афферентная информация поступает в спинной мозг через задние корешки,эфферентнаяимпульсация и регуляция функций различных органов и тканей организма осуществляетсячерезпередниекорешки(законБелла –Мажанди).
Каждыйкорешокпредставляетсобоймножествонервныхволокон.
Всеафферентные входывспинноймозгнесут информациюот трехгруппрецепторов:
1) откожныхрецепторов(болевых,температурных,прикосновения,давления,вибрации);
2) от проприорецепторов (мышечных - мышечных веретен, сухожильных – рецепторовГольджи,надкостницыиоболочексуставов);
3) от рецепторов внутренних органов – висцерорецепторов (механо- и хеморецепторов).Медиатором первичных афферентных нейронов, локализующихся в спинальных ганглиях,является,по-видимому, субстанция Р.
Значение афферентной импульсации, поступающей в спинной мозг, заключается вследующем:
1) участие в координационной деятельности ЦНС по управлению скелетной мускулатурой.При выключении афферентной импульсации от рабочего органа управление им становитсянесовершенным.
2) участиевпроцессахрегуляциифункцийвнутреннихорганов.
3) поддержание тонуса ЦНС; при выключении афферентной импульсации наступаетуменьшениесуммарнойтоническойактивностиЦНС.
4) несет информацию об изменениях окружающей среды. Основные проводящие путиспинногомозга приведены втаблице 1.
Основныепроводящиепутиспинногомозга
| Восходящие(чувствительные)пути
| Физиологическоезначение
|
| Клиновидный пучок (Бурдаха) проходит взаднихстолбах,импуьсация поступаетвкору
| Осознаваемаяпроприорецептивная
импульсацияотнижнейчаституловищаиног
|
| Тонкий пучок (Голля), проходит в заднихстолбах,импульсацияпоступаетвкору
| Осознаваемаяпроприорецептивная
импульсация от верхней части туловища ирук
|
| Заднийспинно-мозжечковый(Флексига)
| Не осознаваемая проприорецептивнаяимпульсация
|
| Переднийспинно-мозжечковый(Говерса)
| Тоже
|
| Латеральныйспиноталамический
| Болеваяитемпературнаячувствительность
|
| Переднийспиноталамический
| Тактильная чувствительность,прикосновение,давление
|
| Нисходящие(двигательные)пути
| Физиологическоезначение
|
| Латеральный кортикоспинальный(пирамидный)
| Импульсыкскелетныммышцам
|
| Переднийкортикоспинальный(пирамидный)
| Тоже
|
| Руброспинальный (Монакова) проходит вбоковыхстолбах
| Импульсы, поддерживающие тонусскелетныхмышц
|
| Ретикулоспинальный, проходит в переднихстолбах
| Импульсы,поддерживающиетонус
скелетныхмышцспомощьювозбуждающихитормозящих влияний на ά- и γ-мотонейроны,атакже регулирующиесостояние
спинальныхвегетативныхцентров
|
| Вестибулоспинальный,проходитвпередних
| Импульсы,обеспечивающиеподдержание
|
| столбах
| позыиравновесиятела
|
| Тектоспинальный, проходит в переднихстолбах
| Импульсы, обеспечивающие осуществлениезрительныхислуховыхдвигательных
рефлексов(рефлексовчетверохолмия)
|
III.Рефлексыспинногомозга
Рефлекторные реакции спинного мозга зависят от места, силы раздражения, площадираздражаемойрефлексогеннойзоны,скоростипроведенияпоафферентными
эфферентным волокнам и, наконец, от влияния головного мозга. Сила и длительностьрефлексовспинногомозгаувеличивается приповторениираздражения(суммация).
Собственная рефлекторная деятельность спинного мозга осуществляется сегментарнымирефлекторнымидугами.
Сегментарная рефлекторная дуга состоит из рецептивного поля, из которого импульсацияпочувствительномуволокнунейронаспинальногоганглия, азатемпоаксонуэтогоже
нейрона через задний корешок входит в спинной мозг, далее аксон может идти прямо кмотонейронупереднегорога, аксонкоторого подходит к мышце.Такобразуется
моносинаптическая рефлекторная дуга, которая имеет один синапс между афферентнымнейроном спинального ганглия и мотонейроном переднего рога. Эти рефлекторные дугиобразуются в таких рефлексах, которые возникают только при раздражении рецепторованнулоспиральныхокончаниймышечныхверетен.
Другие спинальные рефлексы реализуются с участием интернейронов заднего рога илипромежуточнойобластиспинногомозга.Витогевозникаютполисинаптические
рефлекторныедуги.
экстрафузальные волокна четырехглавой мышцы бедра. Реализация этого рефлекса былабы невозможна, если бы одновременно с сокращением мышц-разгибателей нерасслаблялисьмышцы-сгибатели.Рефлекснарастяжениесвойственвсеммышцам,ноу
мышц-разгибателей, они хорошо выражены и легко вызываются.àмотонейроны переднихроговтогожесегментаàзадние рога IIIпоясничного сегментаàзадние корешки à
спинальный ганглий àМиотатические рефлексы — рефлексы на растяжение мышцы.Быстрое растяжение мышцы, всего на несколько миллиметров механическим ударом по еесухожилию приводитксокращению всеймышцыидвигательнойреакции.Например,
легкий удар по сухожилию надколенной чашечки вызывает сокращение мышц бедра иразгибание голени. Дуга этого рефлекса следующая: мышечные рецепторы четырехглавоймышцы бедра
Рефлексы с рецепторов кожи носят характер, зависящий от силы раздражения, видараздражаемого рецептора, но чаще всего конечная реакция выглядит в виде усилениясокращениямышц-сгибателей.
Висцеромоторные рефлексы возникают при стимуляции афферентных нервов внутреннихорганов и характеризуются появлением двигательных реакций мышц грудной клетки ибрюшнойстенки,мышцразгибателейспины.
Рефлексы автономной нервной системы имеют свои пути. Они начинаются от различныхрецепторов, входят в спинной мозг через задние корешки, задние рога, далее в боковыерога,нейроныкоторыхчерезпереднийкорешокпосылаютаксоныненепосредственнок
органам, а к ганглию симпатического или парасимпатического отдела автономной нервнойсистемы.
Автономные (вегетативные) рефлексы обеспечивают реакцию внутренних органов,сосудистой системы на раздражение висцеральных, мышечных, кожных рецепторов. Этирефлексыотличаютсябольшимлатентнымпериодом(ЛП)двумяфазамиреакции:первая
— ранняя—возникаетсЛП7—9мсиреализуетсяограниченнымчисломсегментов,вторая
— поздняя — возникает с большим ЛП — до 21 мс и вовлекает в реакцию практически всесегментыспинного мозга.Позднийкомпонентвегетативного рефлексаобусловлен
вовлечениемвнеговегетативныхцентровголовногомозга.
Сложной формой рефлекторной деятельности спинного мозга является рефлекс,реализующий произвольное движение. В основе реализации произвольного движениялежитγ-афферентнаярефлекторная система.Внеевходятпирамиднаякора,
экстрапирамидная система, α- и γ-мотонейроны спинного мозга, экстра- и интрафузальныеволокнамышечного веретена.
При травмах у человека в ряде случаев происходит полное или половинное пересечениеспинного мозга. При половинном латеральном повреждении спинного мозга развиваетсясиндром Броун-Секара. Он проявляется в том, что на стороне поражения спинного мозга(нижеместапоражения)развивается параличдвигательнойсистемы вследствие
повреждения пирамидных путей. На противоположной поражению стороне движениясохраняются.
Насторонепоражения(нижеместапоражения)нарушаетсяпроприоцептивная
чувствительность. Это обусловлено тем, что восходящие пути глубокой чувствительностиидутпо своейсторонеспинного мозгадо продолговатогомозга, где происходитих
перекрест.
Напротивоположнойсторонетуловища(относительноповрежденияспинногомозга)
нарушается болевая чувствительность, так как проводящие пути болевой чувствительностикожиидутотспинальногоганглия взаднийрогспинного мозга,гдепереключаются на
новыйнейрон,аксонкоторогопереходитнапротивоположнуюсторону.Витогеесли
повреждена левая половина спинного мозга, то исчезает болевая чувствительность правойполовины туловища ниже повреждения. Полную перерезку спинного мозга в экспериментахнаживотныхпроизводятдля исследования влияниявышележащихотделовЦНС на
нижележащие.
Спинальные рефлексы целесообразно объединить по органам (эффекторам рефлекса): а)рефлексы конечностей; б)брюшные; в) яичковый; г) анальный.Рассмотрим каждую группуэтихрефлексов.Рефлексыконечностей.Даннаягруппарефлексовнаиболееобширная,в
Клиническойпрактикеихисследуютнаиболеечасто.Однакоотдельныерефлексы
описываютбезвзаимнойсвязи.Есливкачествеобъединяющегопризнакарефлексов
конечностей принять характер ответной реакции, то все их можно объединить в четырегруппы:1)сгибательные;2)разгибательные;3)ритмические;4)познотонические.
Сгибательные рефлексы делятся на фазные и тонические. Фазные рефлексы — этооднократноесгибаниеконечностиприоднократномраздражениикожиили
проприорецепторов. Одновременно с возбуждением мотонейронов мышц-сгибателейпроисходитреципрокноеторможениемотонейроновмышцразгибателей.Рефлексы,
возникающие с рецепторов кожи, являются полисинаптическими, они имеют защитноезначение. Например, погружение в слабый раствор серной кислоты лапки спинальнойлягушки,подвешеннойнакрючке,илищипоккожиконечностипинцетомвызывают
отдергивание конечности вследствие сгибания ее в коленном суставе, а при более сильномраздражении — и в тазобедренном суставе. Рефлексы, возникающие с проприорецепторов,могутбытьмоносинаптическимииполисинаптическими,напримершейные
познотонические рефлексы. Фазные рефлексы с проприорецепторов участвуют вформировании акта ходьбы. По степени выраженности фазных сгибательных иразгибательныхрефлексовопределяютсостояниевозбудимостиЦНСивозможныеее
нарушения. Тонические сгибательные, а также разгибательные рефлексы возникают придлительном растяжении мышц, их главное назначение — поддержание позы. Тоническоесокращение скелетных мышц является фоновым для осуществления всех двигательныхактов,осуществляемыхс помощьюфазическихсокращениймышц.
Разгибателъныерефлексы,какисгибательные,бываютфазнымиитоническими,
возникают с проприорецепторов мышц-разгибателей, являются моносинаптическими.Одновременно со сгибательным рефлексом возникает перекрестный разгибательныйрефлекс другой конечности.Фазные рефлексы возникают в ответ на однократноераздражение мышечных рецепторов, например при ударе по сухожилию четырехглавоймышцыниженадколеннойчашечки.Приэтомвозникаетколенныйразгибательный
рефлекс вследствие сокращения четырехглавой мышцы (мотонейроны мышц-сгибателей вовремяразгибательногорефлексатормозятся—постсинаптическоереципрокное
торможениеспомощьювставочныхтормозныхклетокРеншоу).Рефлекторнаядуга
коленногорефлексазамыкаетсявовтором—четвертомпоясничныхсегментах(L2—L4).
Фазные разгибательные рефлексы участвуют, как и сгибательные, в формировании актаходьбы.
Тонические разгибательные рефлексы представляют собой длительное сокращение мышц-разгибателей при длительном растяжении их сухожилий. Их роль — поддержание позы. Вположениистоятоническоесокращениемышц-разгибателейпредотвращаетсгибание
нижнихконечностейиобеспечиваетсохранениевертикальнойестественнойпозы.
Тоническое сокращение мышц спины удерживает туловище в вертикальном положении,обеспечивая осанку человека. Тонические рефлексы на растяжение мышц (сгибателей иразгибателей)называют также миотатическими.
Рефлексы позы — перераспределение мышечного тонуса, возникающее при измененииположениятелаилиотдельныхегочастей.Рефлексыпозыосуществляютсясучастием
различных отделов ЦНС. На уровне спинного мозга замыкаются шейные позные рефлексы,наличиекоторыхустановилголландскийфизиологР.Магнус вспециальныхопытахна
кошке. Имеется две группы этих рефлексов — возникающие при наклоне и при поворотахголовы.Первая группарефлексов возникаетпринаклонеголовы вниз(кпереди):
увеличивается тонус мышц-сгибателей передних конечностей и тонус мышц разгибателейзадних конечностей, в результате чего передние конечности сгибаются, а задниеразгибаются. При наклоне головы вверх (кзади) возникают противоположные реакции —передние конечности разгибаются вследствие увеличения тонуса их мышц-разгибателей, азадние конечности сгибаются вследствие повышения тонуса их мышц-сгибателей. Этирефлексывозникаютспроприорецепторовмышцшеиифасций,покрывающихшейный
отдел позвоночника. В условиях естественного поведения они увеличивают животномушансыдостатьпищу, находящуюсявыше илиниже головы.
Втораягруппашейныхпозныхрефлексоввозникаетстехжерецепторов,нотолькопри
поворотах головы вправо или влево. При этом повышается тонус мышц-разгибателей обеихконечностейнастороне,кудаповернутаголова,иповышаетсятонус мышц-сгибателейна
противоположнойстороне.Рефлекс направленнасохранениепозы,котораяможетбыть
нарушена вследствие изменения положения центра тяжести после поворота головы. Центртяжести смещается в сторону поворота головы — именно на этой стороне повышаетсятонус мышц-разгибателейобеихконечностей.Подобныерефлексынаблюдаютсяиу
человека.
Ритмические рефлексы — многократное повторное сгибание и разгибание конечностей.Примерами этих рефлексов могут быть рефлексы потирания у лягушки, чесательный ишагательныйрефлексыусобаки. Рефлекс потираниязаключается втом,чтопосле
смазывания раствором кислоты кожи бедра спинальная лягушка (лягушка, у которойудален головной мозг) многократно потирает этот участок, т. е. пытается освободиться отраздражителя. Слабое раздражение кожи туловища у собаки вызывает почесывание этогоучастказаднейконечностью.Чесательныйрефлекс—аналогрефлексапотиранияу
лягушки. Шагательный рефлекс наблюдают у спинальной собаки, подвешенной с помощьюлямоквстанке.
Брюшные рефлексы (верхний, средний и нижний) проявляются при штриховомраздражениикожиживота;выражаютсявсокращениисоответствующихучастков
мускулатуры стенки живота. Это защитные рефлексы. Для вызова верхнего брюшногорефлекса раздражение наносят параллельно нижним ребрам непосредственно под ними,дугарефлексазамыкается науровне грудныхсегментовспинногомозга(Th8—Th9).
Средний брюшной рефлекс вызывают раздражением на уровне пупка (горизонтально), дугарефлекса замыкается на уровне Th9—Th10. Для получения нижнего брюшного рефлексараздражение наносят параллельно паховой складке (рядом с ней), дуга рефлексазамыкаетсяна уровнеTh11—Th12.
Кремастерный (яичковый рефлекс) заключается в сокращении m. cremaster и подниманиимошонки в ответ на штриховое раздражение верхней внутренней поверхности кожи бедра(кожныйрефлекс),этотакжезащитныйрефлекс;егодугазамыкаетсянауровнеL1—L2.
Анальный рефлекс выражается в сокращении наружного сфинктера прямой кишки в ответна штриховое раздражение или укол кожи вблизи заднего прохода, дуга рефлексазамыкаетсяна уровнеS2 —S5.
в)функцииальфа- игамма–мотонейроновспинногомозга
Аксонмотонейронасвоимитерминаламииннервируетсотнимышечныхволокон,образуя
мотонейронную единицу. Чем меньше мышечных волокон иннервирует один аксон (т. е. чемменьше количественно мотонейронная единица), тем более дифференцированные, точныедвижениявыполняет мышца.
Несколько мотонейронов могут иннервировать одну мышцу, в этом случае они образуюттакназываемыймотонейронныйпул.Возбудимостьмотонейроноводногопуларазлична,
Спинальныйшок
Перерезкаилитравмаспинногомозгавызываетявление,получившееназвание
спинальногошока.Спинальныйшоквыражаетсяврезкомпадениивозбудимостии
угнетении деятельности всех рефлекторных центров спинного мозга, расположенных нижеместа перерезки. Во время спинального шока раздражители, которые обычно вызывалирефлексы,оказываютсянедейственными. Втоже времядеятельностьцентров,
расположенных выше перерезки, сохраняется. После перерезки исчезают не толькоскелетно-моторныерефлексы, но и вегетативные. Снижается кровяное давление,отсутствуютсосудистыерефлексы, актыдефекацииимочеиспускания.
Продолжительностьшокаразличнауживотных,стоящихнаразличныхступенях
эволюционной лестницы. У лягушки шок продолжается 3-5 минут, у собаки – 7-10 дней, уобезьяны–больше1месяца,учеловека –4-5месяцев.Когдашок проходит,рефлексы
восстанавливаются.Причинойспинальногошокаявляетсявыключение
вышерасположенныхотделовголовногомозга,оказывающихнаспинноймозг
активирующее влияние, в котором большая роль принадлежит ретикулярной формациистволамозга.
У человека прослежена последовательность восстановления рефлекторной деятельности ивыявлено несколько стадий при полной параплегии (при разрывах спинного мозга науровнеTh2 до Th12):
1) полнаяарефлексия.Длительностьэтойстадии4-6недель;
2) период, в котором наблюдаются небольшие рефлекторные движения пальцев ног иособеннобольшогопальца.Длительностьэтойстадиисоставляетот2-хнедельдо
несколькихмесяцев;
3) стадияпостепенногоусилениясгибательныхрефлексов;
4) стадия, в которой наблюдаются сгибательные рефлексы, а иногда мощныеразгибательные (спинальное «стояние»). На этой стадии все рефлексы спинного мозгаусиливаются(гиперрефлексия).
36. Спинальные механизмы регуляции тонуса скелетных мышц.Характеристика М- и Н- ответов при стимуляции большеберцовогонерва человека.
Тонус мышц исследуют после поперечной перерезки спинного мозга. Сразу после перерезкив эксперименте или повреждения спинного мозга у человека в случае травмы наблюдаютсямышечнаяатонияиотсутствиерефлексов (спинальныйшок). Главнойпричиной
спинального шока является выключение влияния на спинной мозг вышележащих отделовЦНС(повторная перерезка спинного мозганиже первойповторношокневызывает).
Спинальныйшокулягушек длитсянесколькоминут,усобак—несколькодней,учеловека
— около2мес.Послеисчезновенияспинальногошокатонус мышц,иннервируемых
посредством сегментов спинного мозга, которые находятся ниже перерезки (повреждения),резкоповышается.Сгибательныеиразгибательныерефлексынижнихконечностейу
человекавслучаеповрежденияспинногомозгаусиливаются.
Гипертонусимеетрефлекторнуюприроду,онразвиваетсявследствиеафферентной
импульсации от мышечных рецепторов. Деафферентация, например, нижней конечности улягушкиведеткисчезновениюунеетонуса (опытБронжеста).Имеетсядвафактора,
обеспечивающихафферентнуюимпульсациюотмышечныхрецепторовпозадним
корешкам спинного мозга, которая ведет к возбуждению а-мотонейронов спинного мозга исокращению скелетных мышц при повреждении спинного мозга у человека: 1) спонтаннаяактивность мышечных рецепторов; 2) спонтанная активность γ-мотонейронов, котораяреализуется следующим образом. Возбуждение γ-мотонейронов вызывает возбуждение исокращение иннервируемых ими интрафузальных мышечных волокон, в результате чегоувеличивается натяжение мышечного рецептора, поскольку концы его зафиксированы наскелетноймышце.Вследствиенатяжениямышечноговеретена раздражаютсяи
возбуждаются мышечные рецепторы, обеспечивающие импульсацию к α-мотонейронам ивозбуждение их. В свою очередь α-мотонейроны посылают импульсы к скелетной мышце ивызываютеепостоянное (тоническое) сокращение.
Н-рефлекс-моносинаптическийрефлекс,которыйвызываютвлабо-раторнойи
неврологической практике путем электростимуляции афферен-тных волокон мышечныхнервов(большеберцовогонерваподколеном).ОценкуН-рефлексапроводятсяпоанализу
М- и Н- ответов мышцы. Н – ответ уменьшается при повышении интенсивности стимуляциизасчет встречиего с антидромнымвозбуждением альфа-мотонейрона.
Антидромное возбуждение нейрона- возникает в ответ не раздражение аксона напериферииисвязаносдвустороннимпроведениемвозбужденияпонервномуволокну.
М - ответ- нарастает при увеличении силы раздражения за счет непрямогораздражениямышцысвовлечениемвпроцессвозбужденияновыхдвигательныхединиц
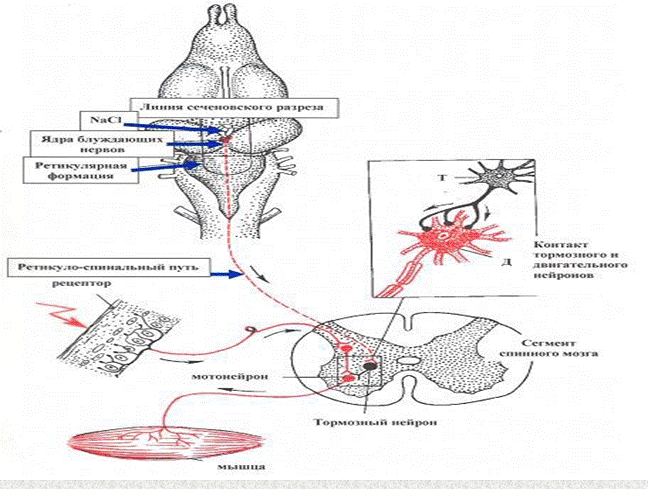
37. Опыт И.М.Сеченова (центральное торможение). Значение этоготорможения в координации рефлекторной деятельности спинногомозга.
В сознании Сеченова, приступившего в Париже к опытам над головным мозгом,синтезировалисьтрипланафизиологическихисследований:
а) изучение мозга путем перерезок его различных частей с последующим физико-химическимраздражением,
б)изучениекожно-мышечныхрефлексов,
в)изучениевнутриорганическихтормозныхвлиянийнамышечнуюдеятельность.
Эти направления подготовили связанный с именем Сеченова физиологический синтез. Ониявилисьестественнонаучнойпредпосылкойсозданнойимсхемыработыголовногомозга.
Всеэкспериментыонпроводилнаклассическомобъектеисследованийфизиологовтого
времени — лягушках. С ювелирной точностью производил разрезы мозга, а затем подолгунаблюдал, какие изменения происходили под влиянием каждого разреза в отражательныхдвижениях животных. Полученные факты говорили сами за себя: угнетение отраженнойдеятельности возникает лишь после разрезов мозга непосредственно перед зрительнымибуграмиивнихсамих.
Явление торможения в нервных центрах было впервые открыто И. М. Сеченовым в 1862 г.Значениеэтогопроцесса былорассмотрено имвкниге«Рефлексыголовного мозга».
Опуская лапку лягушки в кислоту и одновременно раздражая некоторые участки головногомозга(например,накладываякристалликповареннойсолинаобластьпромежуточного
мозга), И. М. Сеченов наблюдал резкую задержку и даже полное отсутствие «кислотного»рефлекса спинного мозга (отдергивания лапки). Отсюда он сделал заключение, что однинервныецентрымогутсущественноизменятьрефлекторную деятельностьвдругих
центрах,вчастностивышележащиенервныецентрымогуттормозитьдеятельность
нижележащих. Описанный опыт вошел в историю физиологии под названием «Сеченовскоготорможения».
Тормозные процессы — необходимый компонент в координации нервной деятельности. Во-первых, процесс торможения ограничивает иррадиацию возбуждения, чем способствует егоконцентрациивнеобходимыхучасткахнервнойсистемы.Во-вторых,возникаяводних
нервных центрах параллельно с возбуждением других нервных центров, процессторможения тем самым выключает деятельность ненужных в данный момент органов,осуществляя координационную функцию. В-третьих, развитие торможения в нервныхцентрахпредохраняетихотчрезмерногоперенапряженияприработе,т.е. играет
охранительнуюроль.
По месту возникновения различают пресинаптическое торможение и постсинаптическое.Постсинаптическое торможение. Оно возникает в постсинаптической мембране нейрона врезультате действия тормозного медиатора и связано с наличием в центральной нервнойсистемеспециальныхтормозныхнейронов.Этоособыйтипвставочныхнейронов,у
которых окончания аксонов выделяют тормозной медиатор. Природа тормозногомедиаторавнастоящеевремяточнонеустановлена.Нервныеимпульсы,подходяк
тормознымнейронам,вызываютвнихтакойжепроцессвозбуждения,какивДругих
нервных клетках. В ответ по аксону тормозной клетки распространяется обычный ПД, но вотличие от других нейронов окончания аксона при этом выделяют не возбуждающий, атормозноймедиатор.Подвлияниемэтогомедиаторавозникаеткратковременная
гиперполяризация постсинаптической мембраны следующего нейрона и регистрируетсятормознойпостсинаптическийпотенциал(ТПСП).Врезультатетормозныеклеткине
возбуждают, а тормозят те нейроны, на которых оканчиваются их аксоны. Такой видторможения называют прямым, так как оно возникает сразу, без предварительноговозбуждения.
Специальные тормозные нейроны. — это клетки Рэншоу в спинном мозгу и корзинчатыеклетки в промежуточном мозгу. Клеткам Рэншоу принадлежит важная роль в координациидеятельностиспинного мозга.Большое значение,например, этиклеткиимеют при
регуляции деятельности мышц-антагонистов. Они обеспечивают развитие торможения вмотонейронах мышц-антагонистов (рис. 52, В), что облегчает осуществление сокращенияэтихмышц.КлеткиРэншоуучаствуютврегуляцииуровняактивностиотдельных
мотонейронов, ограничивая (тормозя) чрезмерное их возбуждениеСХЕМАОПЫТАИ.М. СЕЧЕНОВА.
Основнымипринципамикоординацииявляются:
1. Принципсопряженноготорможенияилиреципрокности
Принципсопряженноготорможенияилиреципрокностибазируетсянаописанномвыше
одноименном виде торможения и означает возникновение при возбуждении одного центраодновременного торможения другого центра для обеспечения цепных рефлексов и сложнойфункции.Примером реципрокностиможет бытьрегуляцияспинным мозгом
противоположныхпофункциональномуназначениюмышцконечностей.Так,при






