ВОЗБУДИМЫЕ ТКАНИ - это нервная, мышечная структуры, которые способны спонтанноиливответнадействиераздражителя возбуждаться.
ВОЗБУДИМОСТЬ – способность живой ткани отвечать на действие сильного, быстрого идлительнодействующегораздражителяизменениемфизиологическихсвойстви
возникновением процесса возбуждения. ВОЗБУЖДЕНИЕ - это генерация ПД +распространениеПД+ответткани,например,сокращение,выделениесекрет.
Меройвозбудимостиявляетсяпорограздражения.ПОРОГРАЗДРАЖЕНИЯ–этота
минимальная сила раздражителя, которая впервые вызывает видимые ответные реакции.Так как порог раздражения характеризует и возбудимость, он может быть назван и порогомвозбудимости.Раздражениеменьшейинтенсивности,невызывающееответныереакции,
называютподпороговым.
РАЗДРАЖИМОСТЬ, т. е. способность ткани изменять свои физиологические свойства ипроявлятьфункциональныеотправления вответна действиераздражителей.
РАЗДРАЖИТЕЛИ – это факторы внешней или внутренней среды, действующие навозбудимыеструктуры.
2. ПРОВОДИМОСТЬ-способностьпроводитьвозбуждение,т.е.проводитьПД
3. СОКРАТИМОСТЬ-способностьразвиватьсилуилинапряжениепривозбуждении
4. ЛАБИЛЬНОСТЬ - или функциональная подвижность - способность ткани воспроизводитьопределенное число волн возбуждения в единицу времени в точном соответствии с ритмомнаносимыхраздражений.Этосвойствохарактеризует скоростьвозникновения
возбуждения.
5. СПОСОБНОСТЬВЫДЕЛЯТЬСЕКРЕТ(секреторнаяактивность),медиатор
6. РЕФРАКТЕРНОСТЬ – временное снижение возбудимости одновременно с возникшим вткани возбуждением. Рефрактерность бывает абсолютной (нет ответа ни на какойраздражитель) и относительной (возбудимость восстанавливается, и ткань отвечает наподпороговыйилисверхпороговыйраздражитель);
Согласно современным представлениям, биологические мембраны образуют наружнуюоболочкувсехживотныхклеток иформируютмногочисленныевнутриклеточные
органеллы.
МЕМБРАНЫ БИОЛОГИЧЕСКИЕ - функционально активные поверхностные структурытолщиной в несколько молекулярных слоев, ограничивающие цитоплазму и большинствоорганелл клетки, а также образующие единую внутриклеточную систему канальцев,складок,замкнутыхобластей.
Биологическиемембраныимеютсявовсехклетках.Мембранныеструктурыклетки
представлены поверхностной (клеточной, или плазматической) и внутриклеточными(субклеточными)мембранами.Названиевнутриклеточных(субклеточных)мембран
обычно зависит от названия ограничиваемых или образуемых ими структур. Так, различаютмитохондриальные, ядерные, лизосомные мембраны, мембраны пластинчатого комплексааппарата Гольджи, эндоплазматического ретикулума, саркоплазматического ретикулума идр.Толщинабиологическихмембран —7—10 нм, ноихобщая площадьоченьвелика.
Одним из важных свойств живых клеток является их электрическая возбудимость, т.е.способностьвозбуждатьсявответ надействиеэлектрического тока.Высокая
чувствительность возбудимых тканей к действию слабого электрического тока впервыебыла продемонстрирована Гальвани в опытах на нервно-мышечном препарате задних лапоклягушки.
Электрические явления, которые возникают в возбудимых тканях, обусловленыэлектрическимисвойствамиклеточныхмембран.
Наиболеехарактернымструктурнымпризнакомявляетсято,чтомембранывсегда
образуют замкнутые пространства, и такая микроструктурная организация мембранпозволяетим выполнятьважнейшие функции.
СТРОЕНИЕИФУНКЦИИКЛЕТОЧНЫХМЕМБРАН.
1. БАРЬЕРНАЯфункциявыражаетсявтом,чтомембранаприпомощисоответствующих
механизмов участвует в создании концентрационных градиентов, препятствуя свободнойдиффузии. К ним относятся механизмы создания потенциала покоя, генерация потенциаладействия.
2. РЕГУЛЯТОРНАЯфункция клеточной мембраны заключается в тонкой регуляциивнутриклеточногосодержимогоивнутриклеточныхреакцийзасчетрецепции
внеклеточных биологически активных веществ, что приводит к изменению активностиферментныхсистем мембраны.
3. ПРЕОБРАЗОВАНИЕ ВНЕШНИХ СТИМУЛОВ НЕЭЛЕКТРИЧЕСКОЙ ПРИРОДЫ ВЭЛЕКТРИЧЕСКИЕСИГНАЛЫ(ВРЕЦЕПТОРАХ).
4. МЕТАБОЛИЧЕСКИЕ ФУНКЦИИ мембран определяются двумя факторами: во-первых, связьюбольшого числа ферментов с мембранами, во-вторых, способностью мембран физическиразделятьклеткуна отдельныеотсеки,отграничивая другот другаметаболические
процессы, протекающие в них. Метаболические системы не остаются при этом полностьюизолированными.
5. МЕЖКЛЕТОЧНЫЕВЗАИМОДЕЙСТВИЯ.Определяютвзаимодействиеклеткисокружающей
средой и формирование многоклеточного организма как единого целого. Молекулярно-мембранныеаспектымежклеточныхвзаимодействийкасаютсяпреждевсегоиммунных
реакций, гормонального контроля роста и метаболизма, закономерностей эмбриональногоразвития..
ХИМИЧЕСКИЙ АНАЛИЗ показал, что мембраны в основном состоят из липидов и белков, иуглеводов, количество которых неодинаково у разных типов клеток. В настоящее времяможноговоритьонесколькихвидахмоделейклеточноймембраны,средикоторых
наибольшеераспространениеполучилажидкостно-мозаичнаямодель.
Согласноэтоймодели,мембранапредставленабислоемфосфолипидныхмолекул,
ориентированных таким образом, что гидрофобные (не взаимодействуют с водой) концымолекул находятся внутри бислоя, а гидрофильные направлены в водную фазу. Такаяструктураидеальноподходитдляобразованияразделадвухфаз:вне-ивнутриклеточной.
В фосфолипидном бислое находятся белки. Эти белки выполняют различные функции, втомчислерецепторную,ферментативную,являютсяпереносчикамиионовимолекул.
ЭЛЕКТРИЧЕСКИЕХАРАКТЕРИСТИКИМЕМБРАН.Особаяморфологияклеточныхмембран
определяет их электрические характеристики, среди которых наиболее важными являютсяемкостьипроводимость.
Через фосфолипидный бислой могут диффундировать различные вещества, причем степеньпроницаемости, т. е. способность клеточной мембраны пропускать эти вещества, зависит отразности концентраций диффундирующего вещества по обе стороны мембраны, егорастворимостивлипидахисвойствклеточноймембраны.Увеличениепроводимости
свидетельствуетобувеличенииколичестваионов,проходящихчерезмембрану.
Видытранспортныхструктурмембраны:
Ионные каналы- это специальные поры (дырочки) в мембране, образованные канальнымибелками, позволяющие ионам проходить через мембрану в обоих направлениях: как внутрь,такинаружу.
ПотенциалзависимыеЛигандзависимыеМеханочувствительныеКаналы«утечки»
2. Транслоказы, - специальные мембранные белки, облегчающие переход вещества черезмембрану за счёт своего временного связывания с диффундирующим веществом. Нетребуютэнергии,работаютвобоихнаправленияхвзависимостиотконцентрации
переносимоговещества.
3. Транспортёры - белковые структуры, насильно протаскивающие определённыевеществасквозьклеточную мембранувопределённомнаправлениисзатратами
энергии. Ионные насосы - это транспортёры ионов. По способу использования энергии длясвоейработы транспортёры можно разделитьна "симпортные"и
"антипортные". Симпортные транспортёры используют совместный транспорт в одномнаправлении двух веществ: одно из них должно иметь большую потенциальную энергиюдлядвижениячерезмембрану.Например, симпортвклеткуспомощьюионов натрия
глюкозы,или симпорт ионов кальция с помощью ионов натрия. Антипортныетранспортёры (обменники) используют встречный транспорт двух веществ с разнойпотенциальной энергией диффузии. Так работает, например, натрий-калиевый ионныйнасос.
Итак,переносвеществчерезклеточнуюмембранупроисходитразличнымипутями.
Механизмы транспорта веществ через мембрануПассивныйтранспорт.
Если вещество движется через мембрану из области с высокой концентрацией в сторонунизкойконцентрации(т.е.поградиентуконцентрацииэтого вещества)беззатраты
клеткой энергии, то такой транспорт называется пассивным, или диффузией. Различают дватипадиффузии:простуюиоблегченную.
Простаядиффузия.
Характернадлянебольшихнейтральныхмолекул(H2O,CO2,O2),атакжегидрофобных
низкомолекулярных органических веществ. Эти молекулы могут проходить без какого-либовзаимодействия с мембранными белками через поры или каналы мембраны до тех пор, покабудетсохраняться градиентконцентрации.
Облегченнаядиффузия.
Характернадлягидрофильныхмолекул,которыепереносятсячерезмембранутакжепо
градиенту концентрации, но с помощью специальных мембранных белков - переносчиков.Для облегченной диффузии, в отличие от простой, характерна высокая избирательность,так как белок переносчик имеет центр связывания комплементарный транспортируемомувеществу,ипереноссопровождаетсяконформационнымиизменениямибелка.Одиниз
возможных механизмов облегченной диффузии может быть следующим: транспортныйбелок (транслоказа) связывает вещество, затем сближается с противоположной стороноймембраны, освобождает это вещество, принимает исходную конформацию и вновь готоввыполнятьтранспортную функцию. Малоизвестноотом,какосуществляется
передвижение самого белка. Другой возможный механизм переноса предполагает участиенескольких белков-переносчиков. В этом случае первоначально связанное соединение самопереходит от одного белка к другому, последовательно связываясь то с одним, то с другимбелком,пока не окажется напротивоположнойстороне мембраны.
Активныйтранспорт.
Имеетместовтомслучае,когдаперенос осуществляетсяпротивградиентаконцентрации.
Такой перенос требует затраты энергии клеткой. Активный транспорт служит длянакоплениявеществвнутриклетки.Источникомэнергиичасто являетсяАТФ.Для
активного транспорта кроме источника энергии необходимо участие мембранных белков.Одна из активных транспортных систем в клетке животных отвечает за перенос ионов Na+ иK+черезклеточную мембрану.ЭтасистеманазываетсяNa+-K+-насос.Онаотвечаетза
поддержаниесостававнутриклеточнойсреды,вкоторойконцентрацияК+выше,чемNa+.
8.  Потенциал покоя, ионные механизмы его формирования. Методыобнаружения иизмеренияпотенциалапокоя.
Потенциал покоя, ионные механизмы его формирования. Методыобнаружения иизмеренияпотенциалапокоя.
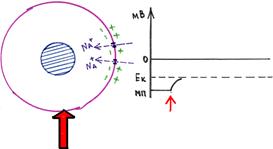
 Мембранныйпотенциалпокоя(МПС)-эторазностьпотенциаловмеждувнешнейи
Мембранныйпотенциалпокоя(МПС)-эторазностьпотенциаловмеждувнешнейи
внутренней сторонами мембраны в условиях, когда клетка не возбуждена. Цитоплазмаклетки заряжена отрицательно по отношению к внеклеточной жидкости неравномернымраспределениеманионовикатионовпообесторонымембраны.Разностьпотенциалов
(напряжение) для различных клеток имеет значения от -50 до -200 мВ (минус означает, чтовнутриклеткаболееотрицательнозаряженная,чемснаружи).Мембранныйпотенциал
покоя возникает на мембранах всех клеток и возбуждающих (нервов, мышц, секреторныхклеток)иневозбуждающих. Потенциалпокоя(ПП)необходимдляподдержания
возбудимости таких клеток, как мышечные и нервные. Также влияет на транспорт всехзаряженных частиц в любом типе клеток: способствует пассивному транспорту анионов изклеткиикатионоввклетку.Образованияиподдержаниямембранногопотенциала
обеспечивают различные типы ионных насосов (в частности натрий-калиевый насос) иионныхканалов(калиевые,натриевые,хлорныеионныеканалы).Квозникновению
мембранного потенциала покоя приводят два фактора: во-первых, концентрации различныхионовотличаются внешнеивнутриклетки, во-вторых,мембранаявляется
полупроницаемой: одни ионы могут через нее проникать, другие - нет. Оба эти явлениязависятотналичиявмембранеспециальныхбелков:концентрационные градиенты
создают ионные насосы, а проницаемость мембраны для ионов обеспечивают ионныеканалы. Важнейшую роль в формировании мембранного потенциала играют ионы калия,натрияихлора. Концентрацииэтихионовотличаются подве сторонымембраны.
Засчетэтихдвухфакторовсоздаютсяусловиядлядвиженияионов.Этодвижение
осуществляется без затрат энергии путем пассивного транспорта – диффузией в результатеразности концент-рации ионов. Ионы K выходят из клетки и увеличивают положительныйзаряд на наружной поверхности мембраны, ионы Cl пассивно переходят внутрь клетки, чтоприводит к увеличению положительного заряда на наружной поверхности клетки. Ионы Naнакапливаются на наружной поверхности мембраны и увеличивают ее положительныйзаряд.Органическиесоединенияостаютсявнутриклетки.Врезультатетакогодвижения
наружная поверхность мембраны заряжается положительно, а внутренняя – отрицательно.Внутренняяповерхностьмембраныможетнебытьабсолютноотрицательнозаряженной,
но она всегда заряжена отрицательно по отношению к внешней. Такое состояние клеточноймембраны называется состоянием поляризации. Движение ионов продолжается до тех пор,поканеуравновесится разностьпотенциалов намембране, т.е. ненаступит
электрохимическоеравновесие.Моментравновесиязависитотдвухсил:
1) силыдиффузии;
2) силы электростатического взаимодействия.Значениеэлектрохимическогоравновесия:
1) поддержаниеионнойасимметрии;
2) поддержаниевеличинымембранногопотенциаланапостоянномуровне.
| Ион
| Концентрациявнутриклетки
| Концентрацияснаружиклетки
|
| Na+
| 12-15мМ
| 145-150 мМ
|
| K+
| 150-155мМ
| 4-5,5мМ
|
| Cl-
| 9 мМ
| 125мМ
|
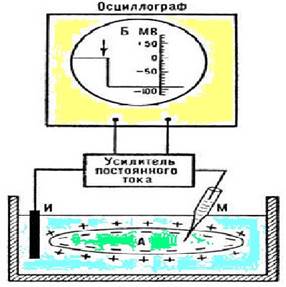
Измерение потенциала покоя: Электроды, которые применяются для измерениямембранныхпотенциалов,
представляют собой тонкие стеклянные капилляры с открытым концом, заполненныерастворомкакоголибоэлектролитаиприсоединенныечерезхлорсеребряныйили
каломельныйэлектродкоднойизклемммилливольтметрасвысокимвходным
сопротивлением. К второй клемме милливольтметра подсоединяется такой же электрод(электрод сравнения), погруженный в наружный раствор. Фактически измеренный ППсоставляетот-90 мВдо-30 мВвразныхклетках.
Послетого,какмикроэлектродпопадаетвклетку,вокругнегочерезнекоторое
время начинает образовываться цитоплазматический чехол с более высоким показателемпреломлениясвета(поэтомуэтотпроцесс легконаблюдатьвполемикроскопа).Этот
чехолначинаетформироватьсяотместапроколаклеткиипостепенноохватываетвесь
микроэлектрод, который он в конце концов и покрывает. Как только формирование чехлазаканчивается, разность потенциалов между цитоплазмой и наружным раствором резкоснижается(на30--40мВ). Внекоторыхслучаях, особеннокогдамикроэлектрод
вводится глубоко в клетку ( на 100 мкм), цитоплазматический чехол может и необразовыватьсявтечение несколькихчасов.
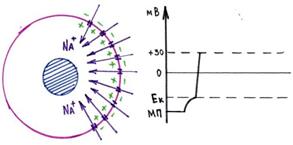
9. Потенциал действия нервного волокна, ионные механизмы егоразвития.Фазывозбудимости.
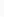 В нервных волокнах сигналы передаются с помощью потенциалов действия, которыепредставляютсобойбыстрыеизменения мембранногопотенциала,быстро
В нервных волокнах сигналы передаются с помощью потенциалов действия, которыепредставляютсобойбыстрыеизменения мембранногопотенциала,быстро
распространяющиесявдольмембранынервноговолокна.Каждыйпотенциалдействия
начинается со стремительного сдвига потенциала покоя от нормального отрицательногозначениядоположительнойвеличины,затемонпочтитакжебыстровозвращаетсяк
отрицательному потенциалу. При проведении нервного сигнала потенциал действиядвижетсявдольнервного волокнавплотьдо его окончания.
Механизмы:
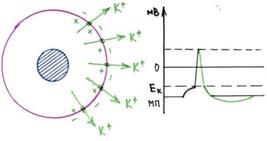
Стадияпокоя.Этастадияпредставленамембраннымпотенциаломпокоя,который
предшествует потенциалу действия. Мембрана во время этой стадии поляризована в связи сналичиемотрицательного мембранного потенциала, равного-90 мВ.
Фаза деполяризации. В это время мембрана внезапно становится высокопроницаемой дляионов натрия, позволяя огромному числу положительно заряженных ионов натриядиффундироватьвнутрьаксона. Нормальноеполяризованноесостояние в-90мВ
немедленно нейтрализуется поступающими внутрь положительно заряженными ионаминатрия, в результате потенциал стремительно нарастает в положительном направлении.Этот процесс называют деполяризацией, В крупных нервных волокнах значительныйизбытоквходящихвнутрьположительныхионовнатрияобычноприводитктому, что
мембранный потенциал «проскакивает» за пределы нулевого уровня, становясь слегкаположительным.Внекоторыхболеемелкихволокнах,какивбольшинственейронов
центральной нервной системы, потенциал достигает нулевого уровня, не «перескакивая»его.
Фаза реполяризации. В течение нескольких долей миллисекунды после резкого повышенияпроницаемости мембраны для ионов натрия, натриевые каналы начинают закрываться, акалиевые—открываться. Врезультате быстраядиффузия ионовкалиянаружу
восстанавливает нормальный отрицательный мембранный потенциал покоя. Этот процессназываютреполяризациеймембраны.
Гиперполяризация мембраны до уровня большего (более отрицательного), чем потенциалпокоя,обусловленаоченьвысокойкалиевойпроницаемостью вфазуреполяризации.
10. Современные представления о процессе возбуждения.Потенциал действия, его фазы развития, ионные механизмыформирования,характеристикафазвозбудимости.
Возбужде́ниевфизиологии—ответтканинараздражение,проявляющийсяпомимо
неспецифических реакций (генерация потенциала действия, метаболическиеизменения) ввыполненииспецифическойдля этойтканифункции;возбудимыми
являютсянервная(проведениевозбуждения),мышечная(сокращение)
и железистая(секреция) ткани. Возбудимость — свойство клеток отвечать на раздражениевозбуждением.
При возбуждении живая система переходит из состояния относительного физиологическогопокояк состоянию физиологическойактивности. Восновевозбуждениялежат
сложные физико-химические процессы. Мерой возбуждения является сила раздражителя,котораявызывает возбуждение.
Возбудимые ткани обладают высокой чувствительностью к действиюслабого электрического тока(электрическая возбудимость), что впервыепродемонстрировалЛ.Гальвани.
ВОЗБУЖДЕНИЕ
свойство живых организмов - активный ответ возбудимой ткани на раздражение. Основнаяфункциядлянервнойсистемы.Клетки,образующиеее,обладаютсвойством проведения
возбужденияизучастка, гдеоновозникло, вдругиеучасткиинасоседниеклетки:
Благодаряэтомунервныеклеткиспособныпередаватьсигналыотоднихструктур
организмакдругим,Темсамымвозбуждениесталоносителеминформацииосвойствах
поступающих извне раздражений и, вместе с торможением, - регулятором активности всехоргановисистеморганизма.Процесс возбуждения возникаетлишьприопределенной
интенсивности внешнего стимула, превышающей абсолютный порог возбуждения (-> порогвосприятия),свойственныйданномуоргану.Воснове возбуждениялежат физико-
химические процессы, с коими связаны биоэлектрические явления, .сопровождающиевозбуждениеирегистрируемыекакнаклеточномуровне,такисповерхноститела
животного и человека. Способы его распространения зависят от формы системы нервной: всамой примитивной - диффузной - системе нервной активность проводится равномерно вразличных направлениях, постепенно затухая. В ходе эволюции наряду с усложнениемсистемынервнойсовершенствовалисьиспособыпередачивозбуждения, прикоихэтот
процесс распространяется до конца пути без какого-либо ослабления, что позволяетвозбуждению выполнять регулирующую функцию в целостном организме. Процессывозбужденияиторможенияобразуетосновудеятельностинервнойвысшей.Их
индивидуальные особенности определяют тип последней (-> деятельность нервнаявысшая:тип),аихдинамика накладываетпечатьнавсе акты поведения.
Стадияпокоя.Этастадияпредставленамембраннымпотенциаломпокоя,который
предшествует потенциалу действия. Мембрана во время этой стадии поляризована в связи сналичиемотрицательного мембранногопотенциала, равного-90 мВ.

11. Анализ графиков одиночного сокращения, потенциаладействия, фаз возбудимости мышечного волокна. Механизмэлектромеханическогосопряжениявскелетноймышце

Механическийответ отдельного мышечного волокна на одиночный потенциал действияназываетсяодиночнымсокращением.Началомышечногонапряжениязапаздываетна
несколько миллисекунд по отношению к потенциалу действия. В течение этого латентногопериодапроходятвсе этапы электромеханическогосопряжения.
Интервалотначаларазвития напряжениядомоментаегомаксимума -этовремя
сокращения . Оно различно для разных типов волокон скелетных мышц. Время сокращениябыстрыхволоконне превышает10мс, тогдакакдляболеемедленныхволоконононе
меньше 100 мс. Длительность сокращения определяется тем, как долго цитоплазматическаяконцентрацияСа2+остается повышенной,обеспечивая продолжениециклической
активности поперечных мостиков. Время сокращения обусловлено активностью Са2+-АТФазысаркоплазматического ретикулума, которая в быстрых волокнах выше, чем вмедленных.
Электромеханическоесопряжениеволоконскелетныхмышц
Электромеханическоесопряжение-этопоследовательностьпроцессов,врезультате
которых потенциал действия плазматической мембраны мышечного волокна приводит кзапускуциклапоперечныхмостиков.Плазматическая мембранаскелетныхмышц
электрически возбудима и способна генерировать распространяющийся потенциалдействияпосредствоммеханизма,аналогичноготому,которыйдействуетвнервных
клетках. Потенциал действия в волокне скелетной мышцы длится 1-2 мс и заканчиваетсяраньше,чемпоявятся какие-либопризнакимеханическойактивности.Начавшаяся
механическая активность может продолжаться более 100 мс. Электрическая активностьплазматическоймембранынеоказываетпрямоговлияниянасократительныебелки,а
вызывает повышение цитоплазматической концентрации ионов Са2+, которые продолжаютактивироватьсократительныйаппаратипослепрекращенияэлектрическогопроцесса.
В состоянии покоя в мышечном волокне концентрация свободного ионизированного Са2+ вцитоплазмевокругтолстыхитонких филаментовоченьнизка,около одной
десятимиллионной доли моля/л. При такой низкой концентрации ионы Са2+ занимаюточень небольшое количество участков связывания на молекулах тропонина, поэтомутропомиозин блокирует активность поперечных мостиков. После потенциала действияконцентрацияионовСа2+ вцитоплазме быстровозрастает, ионисвязываются
с тропонином, устраняя блокирующий эффект тропомиозинаи инициируя циклпоперечныхмостиков. Источникомпоступления Са2+вцитоплазму
являетсясаркоплазматическийретикулуммышечноговолокна.
Саркоплазматический ретикулум мышц гомологичен эндоплазматическомуретикулумудругихклеток.Онрасполагаетсявокругкаждоймиофибриллынаподобие
"рваного рукава", сегментами которого окружены A-дискии I-диски( рис. 30.15). Концевыечастикаждогосегментарасширяютсяввидетакназываемых латеральныхцистерн,
соединенныхдругсдругомсериейболеетонкихтрубок.Влатеральныхцистернах
депонируетсяСа2+;послевозбужденияплазматическоймембраныонвысвобождается.
Отдельную систему составляют поперечные трубочки (T-трубочки), которыепересекаютмышечноеволокнонаграницеA-дисковиI-дисков, проходят
между латеральными цистернамидвух смежныхсаркомерови выходят на поверхностьволокна,составляяединоецелоес плазматическоймембраной.ПросветТ-трубочки
заполненвнеклеточнойжидкостью,окружающеймышечноеволокно.Еемембрана,каки
плазматическая, способна к проведению потенциала действия. Возникнув в плазматическоймембране,потенциалдействиябыстро распространяетсяпоповерхностиволокнаи
мембране Т-трубочек в глубь клетки. Достигнув области Т-трубочек, прилегающих клатеральным цистернам, потенциал действия активирует потенциалзависимые "воротные"белкиихмембраны,физическиилихимическисопряженныескальциевымиканалами
мембраны латеральных цистерн. Таким образом, деполяризация мембраны Т-трубочек.обусловленнаяпотенциаломдействия,приводиткоткрываниюкальциевыхканалов
мембраны латеральных цистерн, содержащих Са2+ в высокой концентрации, и ионы Са2+выходятвцитоплазму.ПовышениецитоплазматическогоуровняСа2+обычнобывает
достаточнымдляактивациивсехпоперечныхмостиковмышечноговолокна.
Процесс сокращения продолжается, пока ионы Са2+ связаны с тропонином, т.е. до тех пор,покаихконцентрациявцитоплазменевернетсякисходномунизкомузначению.
Мембранасаркоплазматического ретикулумасодержит Са2+-АТФазу- интегральный белок,осуществляющийактивныйтранспорт Са2+изцитоплазмыобратновполость
саркоплазматического ретикулума. Са2+ высвобождается из ретикулума в результатераспространенияпотенциаладействияпоТ-трубочкам;дляеговозвращениявретикулум
нужно гораздо больше времени, чем для выхода. Поэтому повышенная концентрация Са2+ вцитоплазмесохраняетсявтечениенекотороговремениисокращениемышечного
волокнапродолжаетсяпослезавершенияпотенциаладействия.
12. Действие постоянного тока на живые ткани. Характеристикапонятий:катэлектротон,анэлектротон,катодическаядепрессия.
Прохождении постоянного тока через живую ткань, является электролитом, вызывает в нейрядфизиологическихизменений, связанныхсперемещением анионовикатионов.
Постоянныйток
В основе биологического действия постоянного гальванического тока лежат процессыэлектролиза,изменения концентрацииионов вклеткахитканяхиполяризационные
процессы. Они обусловливают раздражение нервных рецепторов и возникновениерефлекторныхреакцийместного иобщегохарактера.
В развитии ответных реакций существенную роль играют сила тока, длительностьвоздействия, полярность активного электрода, а также исходное функциональноесостояниеорганов исистем организма.
Припрохождениитокапонервуменяетсявозбудимостьпоследнего.У
катода возникает повышенная возбудимость к раздражителям, у анода — пониженная.Электротон- это есть изменение величины мембранного потенциала и возбудимости подэлектродами при действии постоянного тока на ткань. Различают два вида электротона –катэлектротонианэлектротон.
Катэлектротон-этоуменьшениевеличинымембранногопотенциалаиувеличение
возбудимости под катодом при действии постоянного тока на ткань. Анэлектротон- этоувеличениемембранного потенциала иуменьшениивозбудимостиподанодомпри
действиипостоянноготоканаткань.
Катодическая депрессия - снижение возбудимости ткани под катодом при длительномдействиина нее постоянного электрического тока.
13. Закон частоты раздражения. Понятие лабильности, методыизмерениялабильностивозбудимыхтканей.
Прежде всего, к показателям возбудимости относятся сила и частота раздражителя (первыйзакон раздражения). Исследования свидетельствуют, что возбудимость, сила и частотараздражителя находятся в обратной зависимости, т. е. чем возбудимее ткань, тем меньшийпо силе и частоте раздражитель нужно применить, и, чем меньше возбудимость, тембольшийпосилеичастотетребуетсяраздражитель.Возбудимостьпринятооцениватьпо
порогу силы и частоте раздражителя, которая также находится в обратных соотношениях свозбудимостью.
Лабильностьвфизиологии—функциональнаяподвижность,скоростьпротекания
элементарных циклов возбуждения в нервной и мышечной тканях. Понятие «лабильность»введено русским физиологом Н. Е. Введенским(1886), который считал мерой лабильностинаибольшуючастотураздраженияткани,воспроизводимуюеюбезпреобразованияритма.
Лабильность отражает время, в течение которого тканьвосстанавливает работоспособностьпослеочередного циклавозбуждения.
Лабильность можно измерить, раздражая ткань электрическим током различной частоты.Момент, когда ткань произойдёт преобразование ритма (ткань перестанет воспроизводитьзаданныйритмбезизменений)ибудетлабильностьюданнойткани.Единицыеёизмерения
- количество воспроизводимых импульсов за единицу времени [имп./сек.(мин.), и т. д. ].Проводимость можно охарактеризовать расстоянием, преодолённым импульсом за единицувремени,то естьскоростью распространения импульса.
14. Закон длительности раздражения возбудимых тканей.Характеристикаметодики хронаксиметрии
Закондлительностираздражения
Дляминимальноговозбуждениятребуетсяопределеннаякритическая
пороговая длительность действия раздражителя. Увеличение длительности внешнеговоздействиязапределыпорогаведетк нарастанию возбуждениядо максимальной
величины. Дальнейшее увеличение длительности действия раздражителя не ведет кнарастанию возбуждения.
Эти зависимости сформулированы в законе длительности раздражения: чем длительнеераздражение,темсильнеедо известныхпределовответнаяреакцияживойсистемы.
В основе метода хронаксиметрии лежит оценка возбудимости, исходящая из пороговоговременипротекания определенногоэлектрического раздражителя.
Реакция возбудимой ткани на определенные промежутки времени действия электрическогораздражителяявляетсяболеетонкойичувствительной,чемреакциянаразличную
интенсивностьтока.Так,методомхронаксиметриичастоулавливаютсяизменения,
недоступные классическому методу. Далее, изменения хронаксии в некоторых случаяхпозволяют судить об изменениях в центральных иннервационных влияниях, идущих извысшихотделов нервнойсистемы («субординация»).
Поскольку исследование хронаксии требует подачи импульсов электрического тока весьмамалой длительности (доли миллисекунды), аппаратура для исследования сложнее, чем яридругих методах исследования электровозбудимости. Наиболее распространенным являетсяконденсаторный хронаксиметр, в котором короткие импульсы тока получаются за счетразряда в цепи больного конденсаторов различной емкости, заряженных предварительнодоопределенного потенциала.
15. Закон силы раздражения возбудимых тканей. Сравнительнаяхарактеристикалокальногоответаи потенциаладействия.
Закон силы: чем больше сила раздражителя, тем больше величина ответной реакции. Всоответствии с этим законом функционируют сложные структуры, например, скелетнаямышца.Амплитудаеесокращенийотминимальных(пороговых)величинпостепенно
увеличивается с увеличением силы раздражителя до субмаксимальных и максимальныхзначений. Это обусловлено тем, что скелетная мышца состоит из множества мышечныхволокон,имеющихразличную возбудимость.Поэтомунапороговыераздражители
 отвечают только те мышечные волокна, которые имеют самую высокую возбудимость,амплитуда мышечного сокращения при этом минимальна. С увеличением силыраздражителявреакциювовлекаетсявсебольшееибольшееколичествомышечных
отвечают только те мышечные волокна, которые имеют самую высокую возбудимость,амплитуда мышечного сокращения при этом минимальна. С увеличением силыраздражителявреакциювовлекаетсявсебольшееибольшееколичествомышечных
волокониамплитудасокращениямышцывсевремяувеличивается.Когдавреакцию
 вовлечены все мышечные волокна, составляющие данную мышцу, дальнейшее увеличениесилыраздражителя неприводит кувеличениюамплитудысокращения.
вовлечены все мышечные волокна, составляющие данную мышцу, дальнейшее увеличениесилыраздражителя неприводит кувеличениюамплитудысокращения.
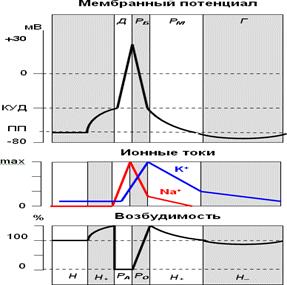
Сравнительнаяхарактеристикалокальногоответаипотенциаладействия.
| Свойство
| Локальныйответ
| Потенциалдействия
|
| Распространение.
| Распространяется на 1-2мм от места раздраженияс затуханием
| Распространяется без затуханияна большие расстояния по всейдлиненервного волокна
|
| Зависимость величиныотсилы стимула
| Возрастает с увеличениемсилы стимула, т.е. подчиняетсязакону«силы»
| Независит (подчиняетсязакону
«всёилиничего»)
|
| Явлениесуммации
| Суммируется - возрастает приповторныхчастых
подпороговыхраздражениях
| Несуммируется
|
| Амплитуда
| 10-40 мВ
| 80-130 мВ
|
| Возбудимость ткани привозникновении
потенциала
| Увеличивается
| Уменьшается вплоть до полнойневозбудимости
(рефрактерность)
|
16. Динамика электромеханического сопряжения в мышечномволокне.Современнаятеориямышечногосокращенияи
Расслабления.
Электромеханическоесопряжение-этопоследовательностьпроцессов,врезультате
которых потенциал действия плазматической мембраны мышечного волокна приводит кзапускуциклапоперечныхмостиков.Плазматическая мембранаскелетныхмышц
электрически возбудима и способна генерировать распространяющийся потенциалдействияпосредствоммеханизма,аналогичноготому,которыйдействуетвнервных
клетках. Потенциал действия в волокне скелетной мышцы длится 1-2 мс и заканчиваетсяраньше,чемпоявятсякакие-либопризнакимеханическойактивности.Начавшаяся
механическая активность может продолжаться более 100 мс. Электрическая активностьплазматическоймембранынеоказываетпрямоговлияниянасократительныебелки,а
вызывает повышение цитоплазматической концентрации ионов Са2+, которые продолжаютактивироватьсократительныйаппаратипослепрекращенияэлектрическогопроцесса.
В состоянии покоя в мышечном волокне концентрация свободного ионизированного Са2+ вцитоплазмевокругтолстыхитонких филаментовоченьнизка,около одной
десятимиллионной доли моля/л. При такой низкой концентрации ионы Са2+ занимаюточень небольшое количество участков связывания на молекулах тропонина, поэтомутропомиозин блокирует активность поперечных мостиков. После потенциала действияконцентрацияионовСа2+ вцитоплазме быстровозрастает, ионисвязываются
с тропонином, устраняя блокирующий эффект тропомиозинаи инициируя циклпоперечныхмостиков. Источникомпоступления Са2+вцитоплазму
являетсясаркоплазматическийретикулуммышечноговолокна.
Саркоплазматический ретикулум мышц гомологичен эндоплазматическомуретикулумудругих клеток. Он располагается вокруг каждой миофибриллынаподобие"рваногорукава",сегментамикоторогоокружены A-дискииI-диски.Концевыечасти
каждого сегмента расширяются в виде так называемых латеральных цистерн, соединенныхдругс другомсериейболеетонкихтрубок.ВлатеральныхцистернахдепонируетсяСа2+;
послевозбужденияплазматическоймембраныонвысвобождается.
Отдельную систему составляют поперечные трубочки (T-трубочки), которыепересекаютмышечноеволокнонаграницеA-дисковиI-дисков,проходят
между латеральными цистернамидвух смежныхсаркомерови выходят на поверхностьволокна,составляяединоецелоес плазматическоймембраной.ПросветТ-трубочки
заполненвнеклеточнойжидкостью,окружающеймышечноеволокно.Еемембрана,каки
плазматическая, способна к проведению потенциала действия. Возникнув в плазматическоймембране,потенциалдействиябыстро распространяетсяпоповерхностиволокнаи
мембране Т-трубочек в глубь клетки. Достигнув области Т-трубочек, прилегающих клатеральным цистернам, потенциал действия активирует потенциалзависимые "воротные"белкиихмембраны, физическиилихимическисопряженныес кальциевыми каналами
мембраны латеральных цистерн. Таким образом, деполяризация мембраны Т-трубочек.обусловленнаяпотенциаломдействия,приводиткоткрываниюкальциевыхканалов
мембраны латеральных цистерн, соде