· трансляция не обеспечивает абсолютной точности воспроизведения аминокислотной последовательности: по ходу полипептида включены “неправильные” аминокислоты; следствие – дефектный фолдинг и функциональная несостоятельность;
· белковые молекулы повреждаются в силу действия агрессивных факторов
· старые белковые молекулы уничтожаются и замещаются скорости синтеза белка
Способы детекции функционально “несостоятельных белков”:
o убиквитиновый; убиквитин – небольшой белок; в ходе атф-зависимого процесса его концевая карбоксильная группа связывается с аминогруппой боковой цепи остатков лизина белка-мишени (метка);
o предположительно, белки для деградации метятся шаперонами;
o помеченные белки разрушаются в основном в лизосомах;
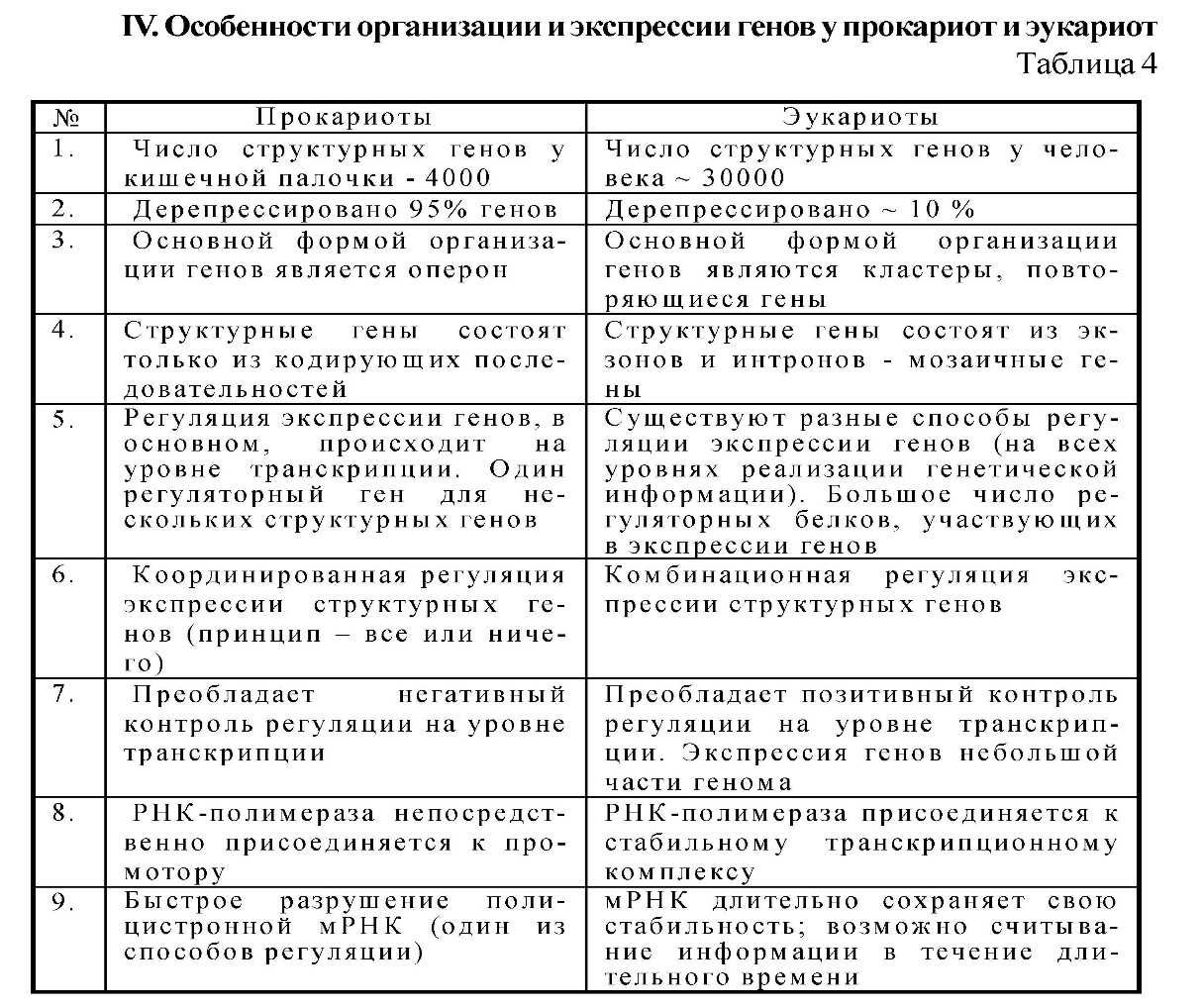
15) Особенности экспрессии генетической информации у про- и эукариот. Взаимосвязь между геном и признаком.
Регуляция экспрессии генов у прокариот. Группа генов, расположенных в ДНК, управляется одним геном-опероном, образую оперон. Оперон – участок ДНК, на котором считывается иРНК, определяет синтез белка-фермента, ген-регулятор – содержит информацию для синтеза.
Регуляция экспрессии генов у эукариот. В клетке прокариота наследственный материал биосинтеза белка пространственно не разобщены, транскрипция и трансляция происходят почти одновременно. Транскриптон – единица транскрипции, состоящей из неинформационной и информационной зоны.
Долгое время ген рассматривали как минимальную часть наследственного материала (генома), обеспечивающую развитие определенного признака у организмов данного вида. Однако каким образом функционирует ген, оставалось неясным. В 1945 г. Дж. Бидлом и Э. Татумом была сформулирована гипотеза, которую можно выразить формулой «Один ген — один фермент». Согласно этой гипотезе, каждая стадия метаболического процесса, приводящая к образованию в организме (клетке) какого-то продукта, катализируется белком-ферментом, за синтез которого отвечает один ген.
Позднее было показано, что многие белки имеют четвертичную структуру, в образовании которой принимают участие разные пептидные цепи. Поэтому формула, отражающая связь между геном и признаком, была несколько преобразована: «Один ген — один полипептид».
Открытия экзон-интронной организации эукариотических генов и возможности альтернативного сплайсинга показали, что одна и та же нуклеотидная последовательность первичного транскрипта может обеспечить синтез нескольких полипептидных цепей с разными функциями или их модифицированных аналогов.
16) Мутационная генотипическая изменчивость. Генные мутации. Характеристика мутаций ядерных генов. Причины и механизмы возникновения генных мутаций. Гены и здоровье человека.
Основу мутационной генотипической изменчивости составляют мутации, происходящие на генном, хромосомном и геномном уровнях;
Мутация – это изменение организации (химической, морфологической), количества и/или интенсивности функционирования генетических структур;
Основные свойства мутаций (гуго де фриз, 1901-1903 г.г.):
· они возникают внезапно;
· они передаются из поколения в поколение (особей или клеток),
· они (генные мутации) наследуются по доминантному или рецессивному типу;
· они не имеют направленности;
· мутирует любой локус, не вызывая видимых, вызывая незначительные или существенные изменения признаков (от простых – полипептиды до сложных);
Одна и та же мутация возникает многократно, то есть может повторяться;
Генные мутации – в строгом смысле это изменения нуклеотидных последовательностей ДНК, воспроизводимые в циклах репликации и проявляющиеся в фенотипе потомков в виде измененных вариантов признака. Это изменение в последовательности нуклеотидов: выпадение, вставка, замена и т.п. Причины – нарушения при удвоении (репликации) ДНК. Примеры: серповидноклеточная анемия, фенилкетонурия.
Генные мутации являются истинными, поскольку только они связаны с изменением биоинформации, записанной и сохраняемой в мутировавшем участке днк. Результатом генных мутаций является образование новой биологической (генетической) информации;
Типы точковых мутаций
Точковые мутации можно разделить на несколько типов в зависимости от характера молекулярного изменения в гене. Здесь мы кратко опишем четыре типа таких мутаций (Wallace, 1981*)
1. Missense-мутация. В одном из триплетов происходит замена одного основания (например, ЦТТ→ГТТ), в результате чего измененный триплет кодирует аминокислоту, отличную от той, которую кодировал прежний триплет.
2. Мутация со сдвигом рамки. Если в последовательность ДНК включается новое основание или пара оснований, то все лежащие за ними триплеты изменяются, что влечет за собой изменение синтезируемого полипептида. Возьмем, например, последовательность АТТ—ТАГ—ЦГА, перед которой включилось основание Т. В результате получится новая последовательность ТАТ—ТТА—ГЦГ—А… К такому же результату приведёт утрата одного из имеющихся оснований.
3. Nonsense-мутация. В результате замены одного основания возникает новый триплет, представляющий собой терминирующий кодон. В генетическом коде имеется три таких триплета. При такой замене синтез полипептидной цепи прекращается в новой (т. е. другой) точке, и соответственно эта цепь отличается своим свойствам от полипептида, который синтез прежде.
4. Синонимическая missence-мутация. Генетический код обладает значительной избыточностью: два или несколько его триплетов кодируют одну и ту же аминокислоту. Поэтому можно ожидать, что в некоторых случаях при замене оснований один триплет заменяется другим — синонимическим, кодирующим ту же аминокислоту. В этом случае, вследствие избыточности кода мы имеем дело с молекулярным изменением в пределах данного гена, которое не вызывает фенотипического эффекта. Такие синонимические мутации, вероятно, довольно обычны.
Генные мутации происходят в области экзонов, интронов, промоторов, 5΄ и 3΄ фланкирующих последовательностях ДНК, в генах энхансеров, факторов транскрипции, сервисных, конценсусных и регуляторных сайтах, в эухроматиновых и гетерохроматиновых участках, другими словами в любых по функционально-генетической характеристике нуклеотидных последовательностях ДНК;
Согласно представлениям классической генетики частота генных мутаций у всех живых форм в среднем укладывается в интервал 10-5 – 10-7 изменений на локус (в другой редакции на гамету) за поколение.
Различают мутации спонтанные (самопроизвольные, происходящие без видимой причины) и индуцированные, вызываемые воздействием физических (температура, ионизирующие излучения), химических (песитициды и инсектициды, некоторые антибиотики, цитостатики), биологических (чужеродные белки сывороток и вакцин, вирусы полиомиелита, вич, простого герпеса), недостаточное белковое питание и других факторов - мутагенов; мутагены – эндогенные (афк, стероидные гормоны, гистамин) и экзогенные (некоторые лекарства, бытовая химия); мутагены подразделяют по их природе на физические (радиоционный мутагенез), химические (химический мутагенез), биологические (биологический мутагенез) и др. (промышленные, бытовые);
Самостоятельная проблема генетики – направленный мутагенез;
Гены и здоровье
Болезни людей с доминирующей или выраженной наследственной составляющей:
Генные/моногенные болезни (гб/мгб) числом порядка 4,5 тыс., диагностируются у 3 детей из 1000, частота в популяциях – в среднем по миру 3%, в разных регионах россии – 4,2-6,5%;
При мгб известно первичное звено поражения фенотипа:
болезни ионных каналов, коллагенопатии, накопления, обмена веществ, гемоглобинопатии и т.п.;
Мгб характеризуются определенным типом наследования;
Мультифакториальные заболевания (мфз): частота – 1,5%, на их долю приходится 94-96% хронической неинфекционной патологии; при мфз наблюдается выраженная генетическая предрасположенность, но необходим “разрешающий” фактор;
Онкогенные болезни (огб): 90% огб относится к категории мфз, 10% - к категории мгб;
Врожденные пороки развития (впр);
Клинические проявления мгб многообразны; основа патогенеза – отсутствие определенного продукта, повышенное образование продукта, образование измененного продукта; важно - генные мутации происходят не только в структурных генах; генокопирование и фенкопирование;
17) Хромосомы – структурные компоненты ядра. Структура и функции хромосом, их динамика в клеточном и митотическом цикле. Хромосомы и здоровье человека.
Хромосомы – нуклеопротеидные структуры в ядрах эукариот, которые являются единицами морфологической организации генетического материала и обеспечивают его точное распределение при делении клетки. Число хромосом в клетках каждого биологического вида постоянно.
Строение.
1)Центромера (первичная перетяжка) это место соединения двух хроматид; к центромере присоединяются нити веретена деления.
По сторонам от центромеры лежат плечи хромосомы. В зависимости от места расположения центромеры хромосомы делят на:
o акроцентрические (палочковидные хромосомы с очень коротким, почти незаметным вторым плечом);
o субметацентрические (неравноплечие, напоминающие по форме букву L);
o метацентрические (V-образные хромосомы, равноплечие).
2)Вторичная перетяжка – ядрышковый организатор, содержит гены рРНК, имеется у одной – двух хромосом в геноме. Теломеры – концевые участки хромосом, содержащие до 10 тысяч пар нуклеотидов с повторяющейся последовательностью. Хромосомы состоят в основном из ДНК и белков, которые образуют нуклеопротеиновый комплекс—хроматин, получивший свое название за способность окрашиваться основными красителями.
Белки составляют значительную часть вещества хромосом. На их долю приходится около 65% массы этих структур. Все хромосомные белки разделяются на две группы: гистоны и негистоновые белки. Помимо ДНК и белков в составе хромосом обнаруживаются также РНК, липиды, полисахариды, ионы металлов.
Функция хромосом заключается:
- В хранении наследственной информации. Хромосомы являются носителями генетической информации.
- В передаче наследственной информации. Наследственная информация передается путем репликации молекулы ДНК.
- В реализации наследственной информации. Благодаря воспроизводству того или иного типа и-РНК и соответственно того или иного типа белка осуществляется контроль над всеми процессами жизнедеятельности клетки и всего организма.
Хромосомы сохраняют свою структурную целостность на протяжении всего митотического (клеточного) цикла; Структура хромосом меняется путем изменения степени компактизации (уплотнения) ее материала по длине;
Регуляторная роль компонентов хромосом заключается в «запрещении» или «разрешении» считывания информации с молекулы ДНК.
В первой половине митоза хромосомы состоят из двух хроматид, соединенных между собой в области первичной перетяжки (центромеры) особым образом организованного участка хромосомы, общего для обеих сестринских хроматид. Во второй половине митоза происходит отделение хроматид друг от друга. Из них образуются однонитчатые дочерние хромосомы, распределяющиеся между дочерними клетками.
Гетерохроматин — участки хроматина, находящиеся в течение клеточного цикла в конденсированном (компактном) состоянии. Особенностью гетерохроматиновой ДНК является крайне низкая транскрибируемость.
ЭУХРОМАТИН - вещество хромосомы, сохраняющее деспирализованное (диффузное) состояние в покоящемся ядре и спирализующееся при делении клеток. Содержит большинство структурных генов организма.
Описано порядка 1000 хромосомных синдромов (хромосомных болезней): в перечень включены синдромы, обусловленные изменением числа отдельных хромосом – моносомии, трисомии;
Согласно современным данным – не менее 50% спонтанных (самопроизвольных) абортов или выкидышей обусловлено хромосомными мутациями; частота хромосомных аномалий среди новорожденных – 0,7%, среди мертворожденных – 5%; хромосомные аномалии у 2-х - 4-х недельных абортусов – 60 -70%, у абортусов 1-го триместра беременности – 50%, у абортусов 2-го триместра – 25-30%; у плодов, погибших после 20 недель беременности, – 7%;
Признаки, указывающие на возможность хромосомной аномалии: отставание ребенка в физическом и психическом развитии, карликовость, черепно-лицевой дисморфизм, пороки развития внутренних органов;
Вероятность рождения ребенка с хромосомной аномалией выше, если возраст женщины 35 лет и более; хромосомные мутации наследуются ребенком чаще от матери; если в семье есть ребенок с хромосомной мутацией, то вероятность 2-го больного ребенка выше;
Синдром «кошачьего крика» Делеция короткого плеча хромосомы 5 - 5p-
Синдром Патау 1:6000 Ж:М 1:1 Трисомия 13 47(+13).95% умирает в первый год
Синдром Эдвардса 1:7000 Ж>М 4:1 Трисомия 18 47 (+18). Продолжительность жизни 2-3 месяца
18) Хромосомные мутации – инструмент комбинативной генотипической изменчивости: изменение дозы генов, перераспределение генов между хромосомами, изменения положения генов в хромосоме. Типы хромосомных мутаций.
Хромосомные мутации, в отличие от генных, не дают новой биологической (генетической) информации; они – инструмент комбинативной генотипической изменчивости. Заключаются в сокращении или увеличении числа определённых генов (изменение дозы генов), перераспределении генов между хромосомами, изменении положения генов в хромосоме;
Структурной предпосылкой хромосомной мутации является нарушение ее целостности - разрыв;
Комбинативная наследственная изменчивость возникает в результате обмена гомологичными участками гомологичных хромосом в процессе мейоза, а также как следствие независимого расхождения хромосом при мейозе и случайного их сочетания при скрещивании. Изменчивость может быть обусловлена не только мутациями, но и сочетаниями отдельных генов и хромосом, новая комбинация которых при размножении приводит к изменению определенных признаков и свойств организма. Такой тип изменчивости называют комбинативной наследственной изменчивостью. Новые комбинации генов возникают:
1) при кроссинговере, во время профазы первого мейотического деления;
2) во время независимого расхождения гомологичных хромосом в анафазе первого мейотического деления;
3) во время независимого расхождения дочерних хромосом в анафазе второго мейотического деления
4) при слиянии разных половых клеток. Сочетание в зиготе рекомбинированных генов может привести к объединению признаков разных пород и сортов.
Различают два основных типа хромосомных мутаций: численные хромосомные мутации и структурные хромосомные мутации.
В свою очередь численные мутации делятся на ан-эуплоидии, когда мутации выражаются в утрате или появлении дополнительной одной либо нескольких хромосом, и полиплоидии, когда увеличивается число гаплоидных наборов хромосом. Потерю одной из хромосом называют моносомией, а возникновение дополнительного гомолога у любой пары хромосом — трисомией.
Структурные хромосомные мутации представлены транслокациями (реципрокными и робертсоновскими), делециями, инсерциями, инверсиями (парацентрическими и перицентрическими), кольцами и изохромосомами. -

Внутрихромосомные аберрации — аберрации в пределах одной хромосомы. К ним относятся делеции, инверсии и дупликации.
Делеция — утрата одного из участков хромосомы (внутреннего или терминального), что может стать причиной нарушения эмбриогенеза и формирования множественных аномалий развития (например, делеция в регионе короткого плеча хромосомы 5, обозначаемая как 5р-, приводит к недоразвитию гортани, ВПР сердца, отставанию умственного развития). Этот симптомокомплекс обозначен как синдром кошачьего крика, поскольку у больных детей из-за аномалии гортани плач напоминает кошачье мяуканье.
Инверсия — встраивание фрагмента хромосомы на прежнее место после поворота на 180°. В результате нарушается порядок расположения генов.
Дупликация — удвоение (или умножение) какого-либо участка хромосомы (например, трисомия по короткому плечу хромосомы 9 приводит к появлению множественных ВПР, включая микроцефалию, задержку физического, психического и интеллектуального развития).
Межхромосомные аберрации — обмен фрагментами между негомологичными хромосомами. Они получили название транслокаций. Различают три варианта транслокаций: реципрокные (обмен фрагментами двух хромосом), нереципрокные (перенос фрагмента одной хромосомы на другую), робертсоновские (соединение двух акроцентрических хромосом в районе их центромер с потерей коротких плеч, в результате образуется одна метацентрическая хромосома вместо двух акроцентрических).
Изохромосомные аберрации — образование одинаковых, но зеркальных фрагментов двух разных хромосом, содержащих одни и те же наборы генов. Это происходит в результате поперечного разрыва хроматид через центромеры (отсюда другое название — центрическое соединение).
19) Митотический (пролиферативный) цикл клетки. Фазы митотического цикла. Главные механизмы пролиферативного цикла. Регуляция митоза. Амитоз. Эндомитоз, политения, их значения.
Клеточный цикл – это период существования клетки от момента ее образования путем деления материнской клетки до собственного деления или смерти.
Обязательным компонентом клеточного цикла является митотический цикл – комплекс взаимосвязанных и детерминированных хронологических событий, происходящих в процессе подготовки клетки к делению и на протяжении самого деления. Кроме того, в жизненный цикл включается период выполнения клеткой многоклеточного организма специальных функций, а также периоды покоя. В периоды покоя ближайшая судьба клетки неопределенна: она может либо начать подготовку к митозу, либо стать на путь специализации.
Главные события митотического цикла заключаются в редупликации (самоудвоении) наследственного материала материнской клетки и в равномерном распределении этого материала между дочерними клетками. По двум главным событиям митотического цикла в нем выделяют репродуктивную (интерфаза) и разделительную (митоз) фазы.
В митотическом цикле выделены 4 периода – митоз (М), а также пресинтетический (G1), синтетический (S) и постсинтетический (G2) периоды интерфазы. Продолжительность митотического цикла для большинства клеток составляет от 10 до 50 ч. Длительность цикла регулируется путем изменения продолжительности всех его периодов.
Митотический цикл включает следующие части:
1. Интерфаза. Занимает 90% времени и включает три периода:
G1 – пресинтетический период (постмитотический) – активный синтез всех белков, построение структур, рост клетки;
S– синтетический – синтез ДНК, продолжается синтез белков, накопление АТФ и активная работа клетки;
G2 - постсинтетический, премитотический – снижается активность клетки, синтез сократительных белков, удвоение пластид, митохондрий и клеточного центра, набухает ядро и к концу клетка выключается из работы и таким образом интерфаза закончена.
2. Митоз – разделительная фаза (10% времени). В митозе можно выделить 4 фазы:
a) В профазе происходит перестройка всей структуры ядра для деления. Ядро увеличивается в объеме, хромосомы становятся видимыми вследствие спирализации, постепенно исчезает ядрышко, растворяется ядерная оболочка, центриоли попарно расходятся к полюсам. Между полюсами протягиваются нити ахроматинового веретена — формируется аппарат, обеспечивающий расхождение хромосом к полюсам клетки. Считывание генетической информации с молекул ДНК становится невозможным: синтез РНК прекращается, ядрышко исчезает. После окончательного распада ядерной оболочки хромосомы беспорядочно размещаются в цитоплазме.
b) В метафаз е спирализация хромосом достигает максимума. Отчетливо видна структура хромосом, их легко сосчитать и изучить их индивидуальные особенности. На этой стадии видно, что каждая хромосома состоит из двух хроматид, соединенных между собой только в области центромеры. Хромосомы располагаются в экваториальной плоскости клетки. Образуется экваториальная (метафазная) пластинка. Веретено деления уже полностью сформировано и состоит из нитей, соединяющих полюса с центромерами хромосом.
c) В анафазе вязкость цитоплазмы уменьшается, центромеры разъединяются и каждая хроматида становится самостоятельной хромосомой. Нити веретена, прикрепленные к центромерам, тянут хромосомы к полюсам клетки. Таким образом, в анафазе хроматиды, удвоенных еще в интерфазе хромосом, точно расходятся к полюсам клетки. В этот момент в клетке находятся два двойных набора хромосом. Число хромосом в соматических клетках всегда парное (диплоидное). Оно образуется после слияния двух половых клеток, в которых всегда бывает одинарное (гаплоидное) число хромосом. Каждый гаплоидный набор обозначается через п, а диплоидный —через 2п. Количество ДНК, соответствующее диплоидному набору хромосом, обозначается как 2с. Два диплоидных набора хромосом, образовавшиеся на стадии анафазы, обозначаются как 4и4с.
d) Телофаза — заключительная фаза митоза. Хромосомы деспирализуются, становятся плохо заметными, но не исчезают. На каждом полюсе клетки вокруг хромосом образуется ядерная оболочка из мембранных структур цитоплазмы. Воссоздаются ядрышки.
П: 2n4c; М: 2n4c; А: 2n2c – 4n4c; Т: 2n2c
На заключительном этапе клеточного деления происходит цитокинез — деление цитоплазматической части клетки. Этот процесс заканчивается образованием в экваториальной зоне клетки перетяжки, которая разделяет делящуюся клетку на две дочерние меньших размеров.
Регуляция клеточного цикла осуществляется окружающими клетками и гуморальными факторами. Существенную роль играют циклоны (особые белки, образующиеся под действием генетической программы), они индуцируют митоз, контролируют длительность его периодов. У животных обнаружены биологически активные вещества (кейлоны), которые показывают насколько они способны ингибировать деление клеток и синтез ДНК.
Амитоз – прямое деление ядра клетки, без формирования веретена деления. В норме он наблюдается в высокоспециализированных тканях, в клетках, которым уже не предстоит делиться: в эпителии и печени позвоночных, в зародышевых оболочках млекопитающих, в клетках эндосперма семени растений. Амитоз наблюдается также при необходимости быстрого восстановления тканей (после операций и травм). Амитозом также часто делятся клетки злокачественных опухолей.
Эндомитоз – процесс умножения числа хромосом в ядрах клеток без образования веретена деления и без деления клеток, в результате чего возникают ядра с увеличенным числом хромосом — полиплоидные ядра. Так возникают двухъядерные клетки (например, клетки печени у человека). В результате серии эндомитоза возникают гигантские полиплоидные клетки красного костного мозга – мегакариоциты. ∞n∞c
Политения – наличие в ядре некоторых соматических клеток гигантских многонитчатых (политенных) хромосом возникших в результате многократного удвоения ДНК без деления клетки.
Политенные хромосомы, так как содержат большое число копий генов, усиливают их экспрессию. Это, в свою очередь, увеличивает производство необходимых специализированной клетке белков. Обнаружена в слюнных железах ряда двукрылых. 2n∞c
20) Мейоз как процесс формирования гаплоидных клеток. Рекомбинация наследственного материала как резерв наследственной (генотипической) изменчивости и основа преадаптации. Генетический груз, его значение.
Мейоз – это процесс образования гаплоидных клеток, т. е. клеток, имеющих половинный набор хромосом. Его можно рассматривать как второй тип деления клеток. Мейоз также можно рассматривать и как специфичный вариант клеточной дифференцировки. Таким способом образуются половые клетки (гаметы) и споры.
Мейоз состоит из двух последовательных делений: редукционное деление (мейоз-1) и эквационное деление (мейоз-2). В каждом из них различают 4 фазы: профазу, метафазу, анафазу и телофазу. Таким образом, весь процесс мейоза условно можно разбить на 8 этапов, плавно переходящих один в другой. Мейоз начинается после S-периода, т. е. после репликации хромосом.
Профаза-1. Наиболее сложная, длительная и важная стадия мейоза. Помимо процессов, аналогичных процессам профазы митоза (спирализация хромосом, разрушение ядерной мембраны, исчезновение ядрышка, образование веретена деления), определяющее значение для всего последующего процесса имеет конъюгация гомологичных хромосом – синапсис. Соединенные пары гомологов называются бивалентами.
Гомологичные хромосомы связывает особая структура, образованная из белков кариоплазмы – синаптонемный комплекс (СК). В бивалентах гомологичные хромосомы могут обмениваться гомологичными участками. Такой процесс называется кроссинговером.
В связи с длительностью и многообразием процессов профазы-1 ее обычно подразделяют на 5 подстадий.
Лептотена – начало спирализации и уплотнения хромосом.
Зиготена – начало (с отдельных участков) и завершение синапсиса гомологичных хромосом. Происходит формирование СК.
Пахитена – укорочение и утолщение бивалентов (стадия толстых нитей).
Диплотена – гомологичные хромосомы бивалентов начинают расходиться (разрушается СК), но они связаны в нескольких зонах контакта – хиазмах. Число хиазм в биваленте может быть различным (обычно 2–3), в длинных хромосомах больше, чем в коротких. Хиазмы часто показывают, что между хроматидами происходит кроссинговер.
Диакинез – хромосомы достигают максимальной спирализации. Исчезают хиазмы, и к концу диакинеза хромосомы остаются связанными только в теломерных участках.
В конце профазы-1 центриоли расходятся к полюсам клетки.
Метафаза-1. Завершается формирование веретена деления. Биваленты концентрируются в экваториальной плоскости клетки.
Анафаза-1. Гомологичные хромосомы расходятся к полюсам клетки. Каждая хромосома состоит из двух хроматид, соединенных общей центромерой.
Телофаза-1. Обычно очень короткая. У полюсов клетки группируются гаплоидные наборы хромосом, в которых представлен только один из пары гомологов. Восстанавливаются структура ядра и ядерная мембрана. Происходит частичная деспирализация хромосом. В конце телофазы-1 наступает цитокинез и образуются две клетки с гаплоидным набором хромосом.
После телофазы-1 вновь образованные клетки сразу вступают в мейоз-2, который проходит по типу обычного митоза.
Профаза-2. Частично деспирализованные хромосомы хорошо различимы. Начинается процесс обратной спирализации хромосом. Разрушается ядерная мембрана, формируется веретено деления, центриоли начинают расходиться к полюсам клетки.
Метафаза-2. Хромосомы выстраиваются в экваториальной плоскости. Центромеры прикрепляются к микротрубочкам образованного веретена деления.
Анафаза-2. Происходит разделение центромер, и каждая хроматида становится самостоятельной хромосомой. Дочерние хромосомы направляются к полюсам клетки.
Телофаза-2. Формируются новые ядра с гаплоидным набором хромосом. Хромосомы деконденсируются. Наступает цитокинез.
В результате мейоза образуются из 1 клетки 4 дочерние - генетически разнородные с гаплоидным набором.
Важным следствием мейоза является обеспечение генетического разнообразия гамет в результате рекомбинации хромосом и кроссинговера. Мейоз создает возможность для возникновения в гаметах новых генных комбинаций, что ведет к изменениям в генотипе и фенотипе потомства.
Кроссинговер. Этот процесс происходит в профазе I мейоза в то время, когда гомологичные хромосомы тесно сближены в результате конъюгации и образуют биваленты. В ходе кроссинговера осуществляется обмен соответствующими участками между взаимно переплетающимися хроматидами гомологичных хромосом. Этот процесс обеспечивает перекомбинацию отцовских и материнских аллелей генов в каждой группе сцепления. В разных предшественниках гамет Кроссинговер происходит в различных участках хромосом, в результате чего образуется большое разнообразие сочетаний родительских аллелей в хромосомах.
Расхождение бивалентов в анафазе I мейоза. В метафазе I мейоза в экваториальной плоскости ахромативнового веретена выстраиваются биваленты, состоящие из одной отцовской и одной материнской хромосомы. Расхождение гомологов, которые несут разный набор аллелей генов в анафазе I мейоза, приводит к образованию гамет, отличающихся по аллельному составу отдельных групп сцепления
В связи с тем, что ориентация бивалентов по отношению к полюсам веретена в метафазе I оказывается случайной, в анафазе I мейоза в каждом отдельном случае к разным полюсам направляется гаплоидный набор хромосом, содержащий оригинальную комбинацию родительских групп сцепления. Разнообразие гамет, обусловленное независимым поведением бивалентов, тем больше, чем больше групп сцепления в геноме данного вида. Оно может быть выражено формулой 2n, где п — число хромосом в гаплоидном наборе. У человека п = 23, и разнообразие гамет, обусловленное этим механизмом, соответствует 223, или 8388608.
Кроссинговер и процесс расхождения бивалентов в анафазе I мейоза обеспечивают эффективную рекомбинацию аллелей и групп сцепления генов в гаметах, образуемых одним организмом.
Преадаптация - эволюционные признаки, которые первоначально не имели явной приспособительной ценности, но которые дают особи, популяции, виду или биоценозу возможность выжить в изменившихся условиях среды. Генетическим механизмом преадаптации является накопление в популяциях скрытого резерва наследственной изменчивости в ходе мутаций.
Генетический груз — накопление летальных и сублетальных «-» мутаций, вызывающих при переходе в гомозиготное состояние выраженное снижение жизнеспособности особей, или их гибель. Это не только совокупность «вредных» генов, но и материал для эволюции, генетическая адаптация.
21) Геномный уровень организации генетического аппарата, его специфический вклад в явления наследственности и биологической изменчивости.
Геном – вся совокупность наследственного материала, заключенного в гаплоидном наборе хромосом клеток данного вида организмов. Геном видоспецифичен, так как представляет собой тот необходимый набор генов, который обеспечивает формирование видовых характеристик организмов в ходе их нормального онтогенеза
Геномный уровень организации наследственного материала, объединяющий всю совокупность хромосомных генов, является эволюционно сложившейся структурой, характеризующейся относительно большей стабильностью, нежели генный и хромосомный уровни. На геномном уровне система сбалансированных по дозам и объединенных сложнейшими функциональными взаимосвязями генов представляет собой нечто большее, нежели простую совокупность отдельных единиц. Поэтому результатом функционирования генома является формирование фенотипа целостного организма. В связи с этим фенотип организма нельзя представлять как простую совокупность признаков и свойств, это организм во всем многообразии его характеристик на всем протяжении индивидуального развития.
В то же время допустимость рекомбинации единиц наследственности в генотипах особей обусловливает генетическое разнообразие их, что имеет важное эволюционное значение. Мутационные изменения, реализующиеся на геномном уровне организации наследственного материала — мутации регуляторных генов, обладающих широким плейотропным действием, количественные изменения доз генов, транслокации и транспозиции генетических единиц, влияющие на характер экспрессии генов, наконец, возможность включения в геном чужеродной информации при горизонтальном переносе нуклеотидных последовательностей между организмами разных видов, — оказываясь иногда эволюционно перспективными, вероятно, являются основной причиной ускорения темпов эволюционного процесса на отдельных этапах исторического развития живых форм на Земле. Таким образом, поддержание постоянства организации наследственного материала на геномном уровне имеет первостепенное значение для обеспечения нормального развития, организма и воспроизведения у особи в первую очередь видовых характеристик.
22) Геномные мутации. Геном и здоровье человека.
Геномные мутации – это изменения числа хромосом
1. Гаплоидия – уменьшение в кариотипе соматических клеток количества хромосом вдвое (2n → n);
2. Полиплоидия – увеличение (кратное n) в кариотипе соматических клеток количества хромосом;
· Аутоплоидия,
· Аллоплоидия;
3. Анэуплоидия (анэусомия) – не кратное гаплоидному (n) изменение в кариотипах соматических клеток количества отдельных хромосом:
· Нулисомия – отсутствие обоих гомологов (у людей нет);
· Моносомия – утрата одного из гомологов, у человека 45,х0 – синдром шерешевского-тернера, моносомий по аутосомам у людей нет;
· Полисомия – дополнительная(ые) половые хромосомы х или у (синдром клейнфельтера), а также аутосомы: трисомия полная по хромосоме 21 – синдром дауна;
· Частичная трисомия по критическому участку хромосомы 21 - q22.3 (фермент супероксиддисмутаза) – синдром дауна в более мягкой форме;
1. Нулисомия, моносомии по аутосомам у людей не совместимы с жизнью; описаны жизнеспособные организмы-мозаики;
2. Полисомии по половым хромосомам – гетерогенность симптоматики, возможны незначительные отклонения;
3. Поли(три)сомии по аутосомам: болезнь дауна -треть погибает внутриутробно, типичная симптоматика, нарушения со стороны системы иммунитета, снижена продолжительность жизни, смерть от инфекций; синдромы при трисомиях 13, 18 и др. – полулетальные мутации с разнообразной симптоматикой;
III. Организменный (онтогенетический) уровень организации биологических систем
1) Размножение организмов. Бесполое и половое размножение. Их сущность, биологическое и эволюционное значение.
Размножение — присущее всем живым организмам свойство воспроизведения себе подобных, обеспечивающее непрерывность и преемственность жизни.
Различают два: бесполое и половое. В основе классификации форм размножения лежит тип деления клеток:
митотический (бесполое), мейотический (половое).
Бесполое размножение поддерживает и усиливает влияние стабилизирующей формы естественного отбора, способствует поддержанию наибольшей приспособленности к мало меняющимся условиям обитания. Происходит без образования гамет, с участием соматических клеток. Участвуют только один организм или его части. Клеточная основа митоз. От одной особи образуется идентичное потомство, клон. Достигается быстрое увеличение численности особей определённого вида. Способствует сохранению, уже имеющихся приспособлений (адаптаций). Является эволюционно более давним
Формы:
1. бинарное деление
2. шизогония, или множественное деление
3. почкование - дочерние особи формируются из выростов тела материнского организма (почек).
4. спорогония - многократное деление ооцисты с образованием спорозоитов.
5. вегетативное размножение (пр.клубника)
6. почкование (гидра)
7. спорообразование (пр. папаротники)
8. фрагментация (пр. планария)
9. полиэмбриония (двойняшки)
Половое размножение способствует движущей форме естественного отбора. Обеспечивает генетическое разнообразие особей и высокий уровень фенотипической изменчивости потомства, чем обеспечивается эволюционная и экологическая пластичность живых существ. В основе лежи





