| Рецепторы
Наиболее
селектив
ные
агонисты
| Мю
| Дельта
| Капа
| Сигма
| |
| D-Ala2-Me-
Phe-Gly-ol5-
Met-
энкефалин
(DAGO)
Мептазинол
Морфоцептин
Суфентанил
TRIMU-4
Бупренорфин
| D-Ala^D-
Leu5-
энкефалин
(DADL)
D-Pen'
D-Pen5-
Энкефалин
(DPDPE)
D-Ser2,
Leu5-
энкефалил-
Thr(DSLET)
| U50.488H
MR 2034
Тифлуа-
дом
UM 1072
| N-алилл-
норметазо-
цин
Фенцикли-
дин
| |
|
| Мет-энкефалин, Лей-энкефалин, Эндорфин, морфин
| |
| Антагонисты
| Налоксазон
Бета-
фуоксиморфин
| ICI 154„129
ICI 174„864
| Mr 2266 BS
| Нет
| |
| |
| |
| |
| Налоксоназин Налоксон, налтрексон, бета-фуналтрексамин
| |
| |
| Основные
эффекты
агонистов
| Аналгезия при
боли любой
природы
Угнетение дыхания
Симпатотормозное действие
Эпилептоген-
ный, кататони-
ческий эффек
ты
| Аналгезия
при боли
любой при
роды
Симпатоак-
тивирующее действие
| Аналгезия
при висцеральной
боли
Седативное действие
| Психотоми-
метический.
галлюцино-
генный
эффекты
| |
| |
| |
| |
| |
| |
| |
В нашей лаборатории разработаны экспериментальные модели, позволяющие анализировать изменения болевых реакций, интегрируемых сегментарными и супрасегментарными структурами головного мозга, при системном введении и непосредственном подведении к этим структурам агонистов различных типов опиатных рецепторов (подробнее см.: Зайцев А.А., 1986; Игнатов Ю.Д., Зайцев А.А., 1987). Полученные нами данные, подтвержденные в других лабораториях (Herman Z., Stachura Z, 1986; Jhamandus К. et al., 1986; Schmauss С., 1987), показали, что каждый агонист определенного типа опиатных рецепторов имеет своеобразную, присущую только ему последовательность вовлечения нейрофизиологических механизмов, что отражает определенную иерархию и рецепторную организацию опиоидергических процессов регуляции боли.
Сложность анализа рецепторных основ опиоидергической регуляции болевой чувствительности усугубляется тем, что на одних и тех же нейронах могут быть представлены опиатные рецепторы различных типов. Так, мю - и дельта-опиатные рецепторы являются частями макромолекулярного комплекса, взаимодействующего посредством аллостерического сцепления, т.е. мю- и дельта-рецепторы взаимосвязаны. С этих позиций представляется вполне уместной гипотеза, согласно которой высокоаффинные мю-рецепторы (мю1-cyбпопуляция) представляют собой общий субстрат для реализации болеутоляющего эффекта всех типов опиатных агонистов (Wolozin D. et al., 1982). Тем не менее есть все основания считать, что в значительной мере опиоидергические системы, формирующие ноцицептивные реакции, дифференцированны как по уровням ЦНС, так и по рецепторам.
В последние годы постулируется существование антиопиоидной системы, которая представлена короткими пептидами MIF-I, Pro-Leu-Gly-NH и Tyr-MIF-I, вызывающими гипералгезию и устраняющими эффекты опиатов и опиоидов принципиально отличным от налоксона способом (Kastin A., Dickson J., 1987). По-видимому, в организме существуют и антагонисты морфина, поскольку катионный хроматиновый белок гистон Н- устраняет эффекты морфина, но не препятствует действию метэнкефалина (Багров А.Я., 1988).
Многочисленными исследованиями показана общность механизмов аналгезии, возникающей при введении опиатов и стимуляции аналгетических зон мозга (подробнее см.: Basbaum A, Filds Н., 1984; Duggan A., 1985; Jensen Т., 1986). В обоих случаях при раздражении антиноцицептивных зон мозга и введении в них опиатов аналгезия опосредуется нисходящими дорсо- и вентролатеральными канатиками спинного мозга, за счет угнетения ответов нейронов заднего рога на ноцицептивные стимулы. В настоящее время не вызывает сомнения существование нисходящих энкефалинергических терминалей, источниками которых являются ядра шва и ретикулярные ядра мозгового ствола. Показано, что энкефалин-иммунореактивные нисходящие терминали образуют синаптические контакты на соме и проксимальных дентритах нейронов I и V пластин заднего рога — источников спиноталамического тракта (Dubner A. et al, 1984). Следовательно, энкефалинергическая модуляция сегментарного ноцицептивного входа осуществляется в основном пост-, а не пресинаптически, как считалось ранее. В спинном мозге выявлены терминали, содержащие одновременно энкефалин и серотонин, которые располагаются преимущественно в I и V зонах заднего рога. Эти терминали, по-видимому, супра-спинального происхождения и их источниками являются энкефалин- и серотонинсодержащие нейроны ядер шва (Tashiro S. et al., 1988).
Роль бета-эндорфина в реализации стимуляционной аналгезии до последнего времени остается дискуссионной. Ранее доминировало представление, что эта аналгезия связана с активацией эндорфинергических систем мозга и с освобождением эндогенного эндорфина. Особенно веские аргументы в пользу этого были получены при изучении стимуляционной аналгезии в клинике, поскольку было показано, что раздражение ЦСВ у человека вызывает аналгезию и одновременно и параллельно развивающееся увеличение эндорфиноподобных субстанций в ликворе желудочков мозга и в спинномозговой жидкости (Tsubokawa Т. et al., 1982). Такое представление нашло экспериментальное подтверждение в последние годы. Было показано, что электрическая стимуляция ЦСВ у крыс селективно активирует популяцию бета-эндорфинсодержащих нейронов этой области, что сопровождается ускорением освобождения бета-эндорфина. Разрушение бета-эндорфиновых нейронов в ЦСВ проявляется резким ослаблением стимуляционной аналгезии (Millan M. et al., 1987). Примечательно, что именно в ЦСВ увеличивается уровень бета-эндорфина при длительной боли, что весьма созвучно гипотезе о способности болевых сигналов активировать эндогенную эндорфиновую систему и дает основание предполагать усиление синтеза бета-эндорфина и ускорение его транспорта в терминалях (Pozzo S. et al., 1983).
Однако представление о ключевой роли эндорфиновой системы в реализации стимуляционной аналгезии разделяется не всеми исследователями. Так, по некоторым данным, повышение уровня эндорфина в спинномозговой жидкости происходит только во время имплантации электродов, что по сути является реакцией на это травматическое хирургическое воздействие (Dioune R. et al., 1982). Увеличение содержания бета-эндорфина не происходит при стимуляции различных зон таламуса, которая вызывает выраженное обезболивание (Tsubokawa Т. et al., 1984). Следует подчеркнуть, что в последние годы акцентировано внимание на стимуляционной таламической аналгезии, отличающейся нейрофизиологическими и нейрохимическими механизмами ее реализации (подробнее см.: Tsubokawa Т. et al., 1985; Roberts С, Rees A., 1986; Kupers К. et al., 1988). Весьма вероятно, что аналгезия, возникающая при стимуляции как таламуса, так и некоторых перивентрикулярных зон мозга, реализуется неопиоидергическими механизмами.
Несомненно, что в ЦНС существует несколько эндогенных аналгетических систем. Согласно гипотезе D. Мауеr и W. Watkins (1984), их четыре — нейронная опиатная, нейронная неопиатиая, гормональная опиатная и гормональная неопиатная. Несмотря на условность, такое разделение имеет определенный смысл, поскольку позволяет достаточно убедительно объяснить ортодоксальность и противоречивость ранее существовавших представлений о влиянии налоксона, о развитии толерантности и перекрестной толерантности к болеутоляющему эффекту опиатов и стимуляции аналгетических зон мозга. Есть основания считать, что раздражение аналгетических зон мозга запускает как опиатную, так и неопиатную нейронные системы мозга. Однако такое разделение является весьма упрощенным, поскольку они весьма тесно взаимосвязаны между собой как функционально, так и морфологически. Особенно отчетливо это показано в отношении нисходящей регуляции, в частности, что нисходящее серотонинергическое торможение на сегментарном уровне опосредуется энкефалинсодержащими тормозными нейронами (Basbaum A., 1982). Более подробную и экспериментально обоснованную схему взаимодействия этих систем представили R. Dubner и соавт. (1984). Согласно их схеме, ноцицептивные спиноталамические нейроны I слоя находятся под тормозным контролем со стороны энкефалинсодержащих нейронов I слоя и стеблевидных нейронов желатинозной субстанции, а спиноталамические нейроны V слоя — под контролем энкефалинсодержащих нейронов того же слоя. Серотонинергические нисходящие терминали модулируют активность спиноталамических нейронов как непосредственно, так и через изменение активности энкефалинсодержащих нейронов.
Большинство гипотез о способности аналгетиков усиливать нисходящее торможение базируется на известной общности проявлений стимуляционной и опиатной аналгезии, хотя эта общность не позволяет судить об их тождественности по существу. Было предложено достаточно много функциональных схем, согласно которым эта аналгезия формируется как за счет непосредственного угнетения релейных нейронов спинного мозга и особенно их активации через высокопороговые ноцицептивные входы, так и в результате усиления опиатами нисходящего торможения с различных антиноцицептивных структур ЦНС (подробнее см: обзоры Gebhart G., 1982; Le Bars D. et al., 1983; Duggan A., North R„ 1984; Fields H., Basbaum A., 1984; Duggan A., 1985). Однако одним из наиболее дискуссионных и наиболее уязвимых мест гипотезы о реализации опиатной аналгезии через эндогенные болеутоляющие системы является положение о способности опиатов усиливать нисходящее торможение и о способах этого усиления с учетом данных об отсутствии у них непосредственного активирующего действия на нейрональную активность. В большинстве исследований, выполненных на ненаркотизированных животных разных видов, выявлено отчетливое и налоксонообратимое угнетение активности клеток центрального серого вещества и ядер шва под влиянием опиатов и опиоидов.
С гипотезой об усилении опиатами нисходящего торможения не согласуются также данные о влиянии морфина на серотонинсодержащие нейроны мозгового ствола. Если морфин действительно увеличивает его торможение, то он должен усиливать нейрональную активность нейронов — источников нисходящих проекций. Однако и ранее уже было известно, что морфин при системном введении или при микроинъекциях в ядра шва не оказывает однонаправленного эффекта на импульсную активность этих клеток (Duggan A., North R., 1984). В опытах с регистрацией активности серотонинсодержащих нейронов большого и дорсального ядер шва у кошек в свободном поведении было показано, что морфин совершенно не усиливает их импульсную активность (Auerbach S. et al., 1985;ShimaK.et al, 1987).
Новое объяснение формирования опиатной аналгезии дали D. Le Bars с соавт. (1983, 1987) с позиций постулируемой ими системы диффузного ноцицептивного тормозного контроля DNIC. Их модель регуляции болевой чувствительности предполагает, что ее усиление (гипералгезия) или угнетение (гипоалгезия) обусловлены фармакологической модуляцией активности нейронов спинного мозга и DNIC, которая высокочувствительна к морфину в малых дозах, еще не влияющих на передачу ноцицептивных сигналов на сегментарном уровне. Как показано этими авторами, морфин в малых дозах сильно и налоксонобратимо угнетает тормозное влияние DNIC на нейроны спинного мозга. Согласно их гипотезе, боль возникает тогда, когда создается определенная контрастность между сигналами ноцицептивных и неноцицептивных нейрональных пулов, запускающих процессы перцепции боли. Опиатная аналгезия может возникать в результате уменьшения контрастности восходящего сигнала двумя способами: во-первых, за счет усиления исходной неноцицептивной активности нейронов, которое происходит под влиянием опиатов в малых дозах и обусловлено угнетением системы DNIC, и, во-вторых, за счет непосредственного угнетения опиатами в больших дозах передачи ноцицептивной информации на сегментарном уровне. Суммация супрасегментарного и сегментарного эффектов опиатов будет приводить к большему уменьшению контрастности сигналов и, следовательно, к усилению аналгезии. Такая гипотеза о действии морфина хорошо согласуется с существующим представлением о синергизме сегментарных и супрасегментарных механизмов опиатной аналгезии. В последнее время получены данные, позволяющие предполагать, что в запуске системы DNIC участвуют и супрасегментарные структуры, в частности, ретикулярная формация среднего мозга, активация которой при воздействии ноцицептивных стимулов на периферии осуществляется через восходящие в вентролатеральных канатиках спинного мозга спиноретикулярные проекции (Vilanueva L. et al., 1986). Реализация эффектов DNIC может осуществляться и за счет освобождения мет-энкефалина в спинном мозге (Le Bars D. et al., 1987).
Представленная схема формирования болеутоляющего действия морфина, несмотря на ее новизну и привлекательность, как и любая другая, не может объяснить все многообразие проявлений феномена опиатной аналгезии. Не ясно, как впишутся в рамки этой схемы болеутоляющие свойства других опиатов и опиоидных пептидов, отличающихся большей селективностью к определенным типам опиатных рецепторов. Недавно было показано, что болеутоляющее действие агонистов мю- и дельта-рецепторов, микроинъецируемых в ЦСВ и ретикулярную формацию мозгового ствола, неодинаково при его определении в тестах горячих пластин и отдергивания хвоста (Jensen S., Yaksh Т., 1986). Как видно на рис. 5, болеутоляющее действие мю-агонистов — морфина и суфентанила — при микроинъекции в крупноклеточное и парагигантоклеточное ретикулярные ядра было значительно больше в тесте горячих пластин, в то время как аналгетический эффект дельта-агонистов был одинаков в тестах горячих пластин и отдергивания хвоста. Это дало основание полагать, что в аналгетическом действии агонистов разных опиатных рецепторов роль усиления ими нисходящего торможения или непосредственного угнетения ноцицептивного потока на супрасегментарном уровне может быть различной. Имеются данные, что для мю-агонистов в большей степени свойственно усиление нисходящего торможения (Dauge V. et al., 1987).
В выполненных в нашей лаборатории экспериментах на бодрствующих крысах болевые реакции оценивали в тесте отдергивания хвоста и по порогу вокализации, отражающим процессы интеграции ноцицептивного сигнала на сегментарном и супрасегментарном уровнях ЦНС соответственно. Морфин и селективные агонисты мю-, дельта - и капа- опиатных рецепторов вводили в боковые желудочки головного мозга и интратекально. Все агонисты при интратекальном введении удлиняют латентный период в тесте отдергивания хвоста. Наибольшая активность выявлена у мю-агонистов, тогда как другие агонисты оказывали меньший эффект. Болеутоляющий эффект агонистов на данной модели обусловлен их непосредственным действием на спинной мозг и хорошо согласуется с существующими представлениями о содержании в нем различных типов опиатных рецепторов. Другая выраженность болеутоляющего эффекта агонистов при интратекальном введении выявлена в тесте вокализации. Только мю- и дельта-агонисты достоверно повышали порог голосовой реакции, что, по-видимому, свидетельствует об их воздействии на рецепторы, локализованные не только на нейронах, реализующих сегментарные ноцицептивные рефлексы, а и на нейрональных элементах, участвующих в формировании восходящего "болевого" потока импульсов.
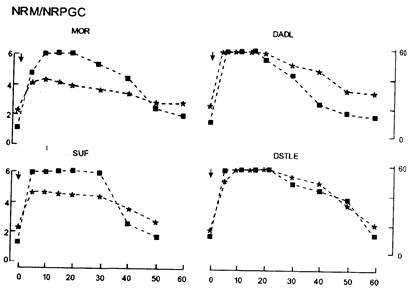
Рис. 5. Болеутоляющее действие агонистов опиатных рецепторов при микроинъекции в ретикулярные ядра мозгового ствола (по Т. Jensen, T Yaksh ^86).
По оси ординат — латентный период отдергивания хвоста (кружки) и латентный период облизывания лап в тесте горячих пластин (квадраты). По оси абсцисс — время, мин. Стрелкой обозначена микроинъекция морфина — МОК (15nmol), суфентанила — SUF (1,5nmol) DADL и dstle (nol,5nmol).
При внутрижелудочковом введении все опиоиды оказывают отчетливое болеутоляющее действие, определяемое по повышению порога вокализации. Характерно, что выраженность эффектов различных агонистов хорошо коррелируют с и имеющимися данными об их локализации в головном мозге (Mack R. et al., 1984) Наши данные хорошо согласуются с результатами исследований, показывающих, что аналгетический эффект агонистов дельта-рецепторов формируется как на супрасегментарном, так и на сегментарном уровнях (HeymanJ.etaL, 1987; MathiasenJ., VaughtJ., 1986). В последнее время в опытах на крысах и обезьянах получены весьма убедительные доказательства, что болеутоляющий эффект капа-агонистов формируется за счет их действия на супрасегментарные структуры (Sasson S, Kornetsky С., 1986; Crionkowsky A. et al 1987; Unterwald E. et al., 1987).
Таким образом, вся совокупность полученных нами данных свидетельствует, что болеутоляющее действие разных опиоидов опостредуется различными механизмами и на разных уровнях ЦНС. Более того, можно считать, что каждый агонист определенного типа опиатных рецепторов имеет своеобразную, присущую только ему последовательность вовлечения нейрональных субстратов и нейрофизиологических механизмов, формирующих его болеутоляющее действие.






