Открытие опиатных рецепторов и их эндогенных лигандов опиоидных пептидов явилось одним из важных достижений медицины и предопределило значительный успех в изучении боли и аналгезии. В настоящее время имеется очень большое количество публикаций по проблеме опиатных рецепторов. Значительная часть из них рассматривается в ряде обзоров последних лет (Булаев В.М., 1982; Смагин В.Г. и др., 1983; Игнатов Ю.Д., 1984; Robson L. et al., 1983; Paterson S. et al., 1984; Holt Y. et aL, 1985; Kosterlitz U., Paterson S., 1985; Yu V. et al., 1986). Общепризнано, что собственно опиатными рецепторами в ЦНС, опосредующими аналгетический эффект опиатов и опиоидов, являются мю-, дельта - и капа-рецепторы.
Сигма-рецепторы рассматриваются в качестве мест специфического связывания, через которые реализуются психотомиметические эффекты некоторых препаратов. С распределением сигма-рецепторов в головном мозге четко совпадают места связывания фенциклидина, обладающего умеренным болеутоляющим и очень сильным психотомиметическим действием. Допускается, что сигма-рецепторы реализуют психогенный, но не аналгетический эффект их лигандов, а соотношение сродства аналгетиков к мю- и дельта-рецепторам может быть своеобразным предиктором выраженности у них психотомиметических свойств (Khazan N. et al., 1984; Zukin S. et al., 1984).
Известно, что количество опиатных рецепторов неодинаково в различных отделах ЦНС и варьирует у различных животных и человека. Большое количество мю-рецепторов содержится в областях ЦНС, участвующих в интеграции болевой чувствительности, в частности, в желатинозной субстанции спинного мозга, в стволовых ядрах шва, в таламусе, гипоталамусе, в IY слое сенсомоторной коры. Дельта-рецепторов особенно много содержится в структурах лимбической системы, в перегородке и гипоталамусе. Мю- и дельта - опиатные рецепторы имеют не только различную локализацию в ЦНС, но и разное развитие в онтогенезе. Формирование мю-рецепторов у крыс происходит в первые 2-3 недели, а дельта-рецепторы отсутствуют у новорожденных крыс и появляются со 2-й по 4-ю неделю после рождения (Me DowellJ., Kitchen I., 1986). Примечательно, что аналгетическое действие опиатов у крыс проявляется только через неделю после рождения, т.е. в то время, когда в ЦНС имеются не только мю-, но и капа - и дельта-рецепторы. Распределение капа-рецепторов в структурах головного мозга животных также имеет существенные видовые особенности (Clark M., 1988). Наиболее высокое содержание капа-рецепторов выявлено в желатинозной субстанции заднего рога шейного и поясничного отделов спинного мозга. Каппа-рецепторы в отдельных структурах головного мозга человека составляют до 40% от общего числа опиатных рецепторов. Наибольшее их количество содержится во фронтальной коре, гипоталамусе, в периакведуктальном сером веществе среднего мозга. Опиатные рецепторы, особенно мю- и капа- выявлены не только в нейрональных элементах, но и в глиальных клетках, что может обусловливать метаболическую и функциональную связь между нейронами и глией (Абутидзе К.Т. и др., 1987).
На современном этапе доказано существование не только отдельных типов рецепторов, но и их подтипов, или изорецепторов — мю1, мю2; капа1, капа2; дельта1, дельта 2,— аналогично тому, как подразделяются рецепторы классических нейромедиаторных систем (Nabeshima Т. et al., 1985; Pasternak G, Wood P., 1986).
Известно, что опиатные рецепторы локализуется на наружной поверхности клеточной мембраны. Однако в опытах на изолированных нейронах виноградной улитки с помощью методов фиксации мембранного потенциала и внутриклеточного диализа получены данные (Громов Т.А., 1992; Громов Л.А. и др., 1986), свидетельствующие о локализации на внутренней поверхности мембраны, по крайней мере, мю- и дельта-рецепторов.
После выявления в структурах ЦНС опиатных рецепторов возникла гипотеза о существовании эндогенных лигандов этих рецепторов. В 1975 г. J. Hughes и соавт. выделили из мозговой ткани эндогенный пептид, названный ими энкефалином. Он оказывал морфиноподобное действие, которое блокировалось антагонистом опиатных рецепторов налоксоном, и его распределение в головном мозге совпадало с локализацией опиатных рецепторов. Открытие этого и других эндогенных лигандов опиатных рецепторов явилось тем событием, которое во многом определило направления дальнейших исследований и современные представления об опиоидергической системе как базовой в регуляции болевой чувствительности.
Эндогенным опиоидным пептидам посвящено очень большое количество исследований (подробнее см.: Ведерникова Н.Н., Майский А.И., 1981; Харкевич Д.А., Титов М.И., 1982; Смагин В.Г. и др., 1983; Калюжный Л.В., 1984; Олейник В.А., Халангот Н.Д., 1985; Виноградов В.А. и др., 1986; Громов Л.А., 1992; Сох В., 1982; Copolov D., Helrne R., 1983; Terenius L., 1985; Kosterlits H, PatersonS., 1985; Willre J., 1986; Traynor J., 1987; и др.). В настоящее время доказано существование по крайней мере трех основных опиоидергических систем: энкефалиновой, эндорфиновой и динорфиновой (рис. 4). Основными предшественниками опиоидных пептидов являются проопиомеланокортин, проэнкефалин, продинорфин, которые обнаружены не только в мозге, но и в других органах (Metters К., Rossier S., 1987).
Энкефалинооая система образована смесью двух пентапептидов с общей NH2-тepминалью, различающихся по одной аминокислоте — метионину (мет-энкефалин) или лейцину (лей-энкефалин). Оба энкефалина образуются в результате протеолиза предшественника — проэнкефалина А. Гипофизарные мет- и лей-энкефалины образуются не только в гипофизе, как считалось ранее, но и в гипоталамусе и оттуда поступают в нейрогипофиз (ZamirN., 1985).
Энкефалины содержатся преимущественно в интернейронах и широко представлены в центральной и периферической нервной системе. Предполагается, что они выполняют функцию нейротрансмиттеров и (или) нейромодуляторов, контролируя высвобождение различных нейромедиаторов. Особенно большое содержание энкефалинов отмечается в полосатом теле, гипоталамусе, в желатинозной субстанции задних рогов спинного мозга, в продолговатом и среднем мозге, в заднем таламусе, в ядрах мозжечка и некоторых других структурах головного мозга.

Pre - Pro - Enkepholin A (263 residues)
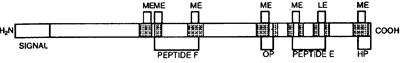
Pre - Pro - Enkepholin В = Pre - Pro - Dynorphin (256 residues)

Рис. 4. Схема структуры предшественников опиоидных пептидов (по М. Millan, 1986).
АСТН — адренокортикотропин, CLIP — кортикотропиноподобный промежуточный пептид, DYN — динорфин, END — эндорфин, HP — гептапептид, LPH — липотропин, MSH — мелоностймулирующий гормон, NE — неоэндорфин, ОР — октапептид.
Высокая концентрация эндорфинов обнаруживается в гипофизе, где они локализуются в секреторных гранулах и высвобождаются вместе с кортикотропином. Прекурсором эндорфинов является проопиомеланокортин, точнее его фрагмент бета-липотропин, не обладающий опиатоподобной активностью. В результате гидролиза бета-липотропина, содержащего 91 аминокислоту, образуется бета - эндорфин (бета-липотропин 61-91), а также а-эндорфин (61-76) и гамма-эндорфин (61-77). Помимо гипофиза, эндорфины обнаруживаются в некоторых отделах гипоталамуса, таламуса, среднего мозга и моста, где им придается значение нейромедиаторов или нейро-модуляторов ноцицепции. В целом эндорфины более ограничено распространены в ЦНС, чем энкефалины, и их распределение по отделам головного мозга также неодинаково. Незначительная эндорфиноподобная иммунореактивность отмечается в некоторых периферических органах — надпочечниках, плаценте, вегетативных ганглиях.
Основными представителями динорфинов являются гептадекапептид динорфин А, тридекапептид динорфин В, а также выделенный из гипоталамуса декапептид а-неодинорфин и их более короткие фрагменты (динорфин 1-13, динорфин 1-8 и др.) и аналоги (бета-неодинорфин). Все динорфины тесно взаимосвязаны в гипоталамусе и являются продуктами протеолитических преобразований одного прекурсора — продинорфина.
В последние годы (подробнее см.: Сох В., 1982; Neubert K. et al., 1986) обнаружен еще целый ряд эндогенных опиатоподобных пептидов. Бета-казоморфины выделены из гидрилизата бычьего казеина, а затем из молока различных животных и человека. Группа бета-казоморфинов включает в себя ряд соединений с различным содержанием аминокислот, а также альфа-казеин. В синаптосомах головного мозга быков и крыс обнаружен дипептид, получивший название киоторфин. Он прямо не влияет на опиатные рецепторы, но оказывает опиатоподобное действие за счет стимуляции выделения энкефалинов. В последние годы получены данные о существовании в мозге и спинномозговой жидкости человека и некоторых животных таких новых опиоидных пептидов, обладающих болеутоляющей активностью, как дерморфин, лейморфин, риморфин (Conneras P. et al., 1987).
Поскольку все эндогенные опиаты являются пептидами, в тканях организма они подвергаются интенсивному гидролизу, что определяет непродолжительность их эффекта (подробнее см.: Schwartz J., 1983; Rocfues В., Fourme-Zaliski M., 1985; McKelvy J., 1986). Выделено несколько групп пептидаз, имеющих разную специфичность и направленность действия на структуру опиатов. В настоящее время ведется активный поиск ингибиторов этих ферментов, рассматриваемых в качестве потенциальных лекарственных препаратов для пролонгации эффекта энкефалинов и эндорфинов. Гидролиз энкефалинов на уровне Tyr1-Gly2 неспецифически осуществляется аминопептидазами, представителями металлоферментов. Кроме металлопептидаз, обнаружены дипептидиламинопептидазы, воздействующие на связь аминокислот Gly2-Gly3. На уровне Gly3-Phe4 гидролиз энкефалинов осуществляется ангиотензинконвертирующим ферментом (пептидилпептидгидролазой), который является важным звеном метаболизма в ренин-ангиотензинной системе. Наибольшая специфичность гидролизующего эффекта присуща так называемым энкефалиназам, точка приложения которых аналогична ангиотензинконвертирующему ферменту. В отличие от последнего, энкефалиназы мало чувствительны к депримирующему эффекту каптоприла, однако их эффективность отчетливо тормозится бестатином, тиорфаном, пуромицином. Менее исследован катаболизм эндорфинов, по крайней мере этот процесс имеет существенные особенности. Предполагают, что эндорфины инактивируются эндопептидазами, в том числе энкефалиндеградирующейаминопептидазой.
В настоящее время не вызывает сомнений, что энкефалины, эндорфины и динорфины являются звеньями общей эндогенной антиболевой системы головного мозга и вместе с тем в ее рамках имеют присущие каждому из них особенности механизмов регуляции болевой чувствительности. Эти особенности в значительной мере обусловлены различиями в нейрофизиологических, в частности, синаптических эффектах разных эндогенных опиатных пептидов. Энкефалины за счет деполяризации мембран нейронов мозга способствуют мобилизации Са из внутриклеточных депо и усиливают активируемый ионами кальция транспорт ионов калия, причем функциональная организация механизмов, запускаемых при активации афферентных волокон различного диаметра, неоднозначна. При стимуляции А-волокон тонически малоактивный тормозный энкефалинергический нейрон активируется, вследствие чего уменьшается ощущение боли. При ноцицептивной стимуляции С-волокон энкефалинергический нейрон через тормозный вставочный нейрон тормозится и облегчается передача болевых импульсов к мотонейронам спинного мозга и к супрасегментарным структурам, участвующим в формировании ответных реакций организма на боль. Эндорфины вызывают гиперполяризацию нейронов ЦНС. Они вместе с динорфинами представлены преимущественно в длинноволоконных системах, тормозящих распростанение болевых импульсов на разных уровнях головного мозга. Кроме того, эндорфины выделяются в кровь, где выполняют функцию нейрогормонов и опосредуют эндокринные сдвиги при боли.
Однако ключевое значение в биологическом действии эндогенных опиатов имеет их аффинитет к различным типам опиатных рецепторов. Так, установлено, что мет- и лей-энкефалины имеют наибольшее средство к дельта-опиатным рецепторам, причем удаление одной аминокислоты из их структуры полностью устраняет болеутоляющее действие. Динорфины высокоселективны к капа-опиатным рецепторам, и эта селективность существенно возрастает при укорочении их структуры. Эндорфины, напротив, слабо взаимодействуют с капа-рецепторами, но обладают большим аффинитетом к мю - и дельта-опиатным рецепторам. Даже при значительном укорочении эндорфинов, например, у бета-эндорфина 1-31 до бета-эндорфина 1-16, опиатная активность сохраняется.
В последние годы созданы и изучены стабильные аналоги эндогенных пептидов и других химических соединений; отличающиеся селективностью к определенному типу опиатных рецепторов (табл. 1) (Clark J. et aL, 1986). Болеутоляющий эффект опиатов при воздействиях любой природы реализуется через мю-опиатные рецепторы, при боли термической природы важную роль играет также их влияние на бета-рецепторы, а при боли висцерального генеза — на капа-рецепторы. Основное значение в вызываемом опиатами угнетении дыхания придается их влиянию на мю2-субпопуляцию опиатных рецепторов и дельта-опиатные рецепторы, тогда как седативный, гипотермический, каталептогенный эффекты и толерантность к опиатам формируются через мю1-рецепторы. С активацией метэнкефалинергических и капа-опиоидергических механизмов связывают симпатоактивирующий эффект. Роль дельта-опиатных рецепторов в опиатной аналгезии дискутируется, однако им придается важное значение в психотропном, в частности, дисфорическом действии опиатов. Следует особо подчеркнуть, что данные о селективности болеутоляющего действия различных агонистов получены исключительно на моделях, позволяющих оценить изменение лишь простых ноцицептивных реакций, которые отражают не только аналгетическое действие веществ, а могут возникать вследствие нарушения моторной сферы.
Таблица 1.






