Типы клеток взрослого человека
Ороговевающие эпителиальные клетки
Кератиноциты эпидермиса (диффенцирующиеся эпителиальные клетки)

Базальная клетка эпидермиса (стволовая клетка)
Кератиноцит ногтей
Базальная клетка ногтевого ложа (стволовая клетка)
Клетки стержня волоса
Клетка сердцевины волоса
Кортикальная
Кутикулярная
Клетки оболочки корня волоса
Кутикулярная
Слоя Huxley (Гексли)
Слоя Henle (Генле)
Наружная
Клетка волосяного матрикса (стволовая клетка)
Клетки влажных плоских барьерных эпителиев
Поверхностная эпителиальная клетка многослойного плоского эпителия роговицы, языка, ротовой полости, пищевода, прямой кишки, дистальной части уретры, влагалища
Базальная клетка тех же видов эпителия (стволовая клетка)
Клетка мочевыводящих путей эпителия (выстилающая мочевой пузырь и мочевыводящие пути)
Эпителиальные клетки, специализированные на экзокринной секреции
Клетки слюнной железы
Слизистая клетка (секрет богат полисахаридами)
Серозная клетка (секрет богат гликопротеиновыми ферментами)
Клетка железы фон Эбнера в языке (секрет служит для омывания вкусовых сосочков)
Клетка молочной железы, секретирующая молоко
Клетка слезной железы, секретирующая слезы
Клетка церуминовой железы уха, секретирующая ушную серу
Клетка эккриновой потовой железы, секретирующая гликопротеины (темная клетка)
Клетка эккриновой потовой железы, секретирующая малые молекулы (светлая клетка)
Клетка апокриновой потовой железы (выделяет пахучий секрет, чувствительна к половым гормонам)
Клетка железы Молля в веке (специализированная потовая железа)
Клетка сальной железы, секретирующая богатое липидами кожное сало
Клетка боуменовой железы в носу (секретирует жидкость, омывающую обонятельный эпителий)
Клетка бруннеровой железы в двенадцатиперстной кишке, секретирующая щелочной раствор слизи и ферментов
Клетка семенного пузырька, секретирующая компоненты семенной жидкости, включая фруктозу (как источник энергии для движения спермиев)
Клетка предстательной железы, секретирующая другие компоненты семенной жидкости
Клетка бульбоуретральной железы, секретирующая слизь
Клетка бартолиниевых желез, секретирующих жидкость для увлажнения влагалища
Клетка железы Литтре, секретирующая слизь
Клетка эндометрия матки, секретирующая в основном углеводы
Изолированная бокаловидная клетка дыхательного и пищеварительного трактов, секретирующая слизь
Слизистая клетка выстилки желудка
Замоченная клетка желез желудка, секретирующая пепсиноген
Обкладочная клетка желез желудка, секретирующая HCI
Клетка ацинуса поджелудочной железы, секретирующая пищеварительные ферменты и бикарбонат
Клетка Панета в тонком кишечнике, секретирующая лизоцим
Пневмоцит II типа в легком, секретирующий сурфактант
Клетка Клара легких (функция неизвестна)
Сократительные клетки
Клетки скелетных мышц

Красные (медленные)
Белые (быстрые)
Промежуточные
Мышечное веретено с ядерной сумкой
Мышечное веретено с ядерной цепочкой
Клетка-сателлит (стволовая клетка)
Клетки сердечной мышцы
Обычные
Узловые
Волокна Пуркинье
Клетки гладкой мускулатуры (разные)

Миоэпителиапьные клетки
Радужной оболочки
Экзокринных желез
Чувствительные рецепторы
Фоторецепторы
Палочки
Колбочки
Чувствительные к синему
Чувствительные к зеленому
Чувствительные к красному
Слуховые рецепторы
Внутренние волосковые клетки кортиева органа
Наружные волосковые клетки кортиева органа
Рецепторы гравитации и ускорения
Волосковая клетка I типа вестибулярного аппарата
Волосковая клетка II типа вестибулярного аппарата
Вкусовые рецепторы

Клетка II типа вкусового сосочка
Рецепторы запаха
Обонятельный нейрон
Базальная клетка обонятельного эпителия (стволовая клетка обонятельных нейронов)
Рецепторы pH крови
Клетка каротидного тельца
I типа
II типа
Тактильные рецепторы
Меркелевы клетки эпидермиса
Первичные обязательные нейроны (разные)
Рецепторы температуры
Первичные терморецепторные нейроны (разные)
Чувствительные к холоду
Чувствительные к теплу
Рецепторы боли
Первичные нейроны, воспринимающие боль (разные)
Рецепторы положения и напряжений в скелетно-мышечной системе
Проприоцептивные первичные чувствительные нейроны (разные)
Автономные нейроны
Холинэргические (разные)
Адренэргические (разные)
Пептидэргические (разные)
Клетки хрусталика
Клетка переднего эпителия хрусталика
Волокно хрусталика (клетка, содержащая кристаллин)
Пигментные клетки

Меланоцит
Эпитальная клетка пигментного слоя сетчатки
Половые клетки
Оогоний/ооцит
Сперматоцит
Сперматогоний (стволовая клетка сперматоцита)
Питающие клетки
Клетка фолликула яичника
Клетка Сертоли (в семеннике)
Эпителиальная клетка тимуса
Клеточные фенотипы различны по морфологиям. Морфология клеток в основном определяется их функцией. Клетки сходные по морфологии, функции, биохимии составляют ткань. Основные типы клеток:
1. Клетки эпителия. Внешняя поверхность тела и почти все внутренние поверхности покрыты непрерывным слоем клеток, который называется эпителием и состоит из эпителиальных клеток. Эта ткань отличается плотными межклеточными контактами и формирует прочный покров, непроницаемый для жидкостей.

Эпителиальные клетки полярны, так как их поверхности (апикальная и базолатеральная) различаются. Эти клетки имеют широкий спектр специализации, и одной из наиболее значимых функций является их способность секретировать жидкости. Иногда эти вещества секретируются непосредственно на поверхность клетки, в других случаях клетки эпителия формируют протоки или каналы, через которые секрет выводится из ткани. Некоторые эпителиальные клетки выделяют секрет непосредственно в кровь; эти клетки обычно расположены вблизи капиллярной сети.
Клетки соединительной ткани. Клетки соединительной ткани выполняют опорную, соединительную и питательную функции в отношении клеток других тканей. Многие типы клеток этой группы производят значительные количества внеклеточного матрикса. Организация этого матрикса в основном имеет белковую природу; он содержит различные типы коллагенов и другие структурные белки, такие как фибронектин, ламинин и витронектин. Одна из основных функций соединительной ткани — синтез уникальных типов коллагена и других молекул матрикса. Важным структурным элементом, состоящим из соединительнотканных клеток, является базальная пластинка; на ней расположено большинство эпителиальных клеток. Хотя базальная пластинка в основном состоит из веществ, синтезированных клетками соединительной ткани, другие типы клеток тоже иногда вносят вклад в ее состав.
Фибробласт — относительно недифференцированная клетка соединительной ткани.

Он служит клеткой-предшественником, из которой формируются другие клетки соединительной ткани, включая адипоциты (клетки жировой ткани), гладкие мышечные клетки и клетки, продуцирующие костную и хрящевую ткань. Определенные фибробласты специализированы на продукции, модификации и перестройке костей и хрящей. Эти клетки называются остеобластами, остеоцитами, хондробластами и хондроцитами. Построение кости и хряща — динамический процесс, который длится в течение всей жизни организма. Клетки крови, включая эритроциты, моноциты, нейтрофилы, базофилы, эозинофилы, тромбоциты, происходят из клеток соединительнойткани. Эти клетки продуцируются миелоидной тканью внутреннего отдела костей, называемой костным мозгом.
Клетки мышечной ткани. Третьей основной тканью является мышца. Хотя все клетки этой ткани специализированы на сокращении, они обычно заметно отличаются друг от друга морфологически и функционально. Существуют четыре вида сократимых клеток: клетки скелетных или поперечнополосатых мышц, клетки сердечной мышцы, клетки гладкой мускулатуры (производные фибробластов) и мио- эпителиальные клетки (производные эктодермы).
Клетки скелетных мышц обычно имеют сильно удлиненную форму, их часто называют мышечными волокнами. Отдельное мышечное волокно (клетка) представляет собой синцитий, который содержит множество ядер и общую цитоплазму. Новые мышечные волокна образуются при слиянии миобластов. Клетки сердечной мышцы, как и клетки поперечнополосатых мышц, формируют синцитий. Сократительные белки (миозин, актин и другие структурные белки) организованы в правильную линейную структуру, называемую саркомером. Клетки гладкой мускулатуры не формируют синцитий. Они собраны в длинные пучки, и их сокращение гораздо слабее и длительнее, чем у клеток поперечнополосатой или сердечной мышцы. Гладкомышечные клетки, важный компонент кровеносных сосудов, участвуют в распределении кровотока, особенно в системе микроциркуляции. Миоэпителиальные клетки также не имеют поперечной исчерченности и развиваются, скорее, из эктодермы, чем из мезодермы, которая является предшественником всех остальных мышечных клеток. Эти сокращающиеся клетки регулируют ответ определенных чувствительных клеток, таких как клетки радужной оболочки, потовых, молочных и других желез, реагирующих на сенсорные стимулы.
Клетки нервной ткани. Нервные клетки, четвертый класс основных тканей, характеризуются своей «раздражимостью» и способностью проводить электрические импульсы. Эти клетки составляют основную коммуникативную сеть организма.

Нейроны делятся на три больших группы: униполярные, биполярные и мультиполярные. Эта классификация отражает различия в количестве и расположении отростков, исходящих из тела нервной клетки.Простейший униполярный нейрон имеет один главный отросток со многими ответвлениями; одно из них служит аксоном, а другие — дендритами. У беспозвоночных в основном найдены униполярные нейроны. Биполярные нейроны имеют два основных отростка: дендритный отросток с множеством ответвлений, который проводит информацию от периферии к телу клетки, и аксон, который передает информацию от клетки к другим нейронам или клеткам-мишеням. Аксоны нейронов заканчиваются межнейронными соединениями, которые называются синапсами. Электрические импульсы передаются от одного нейрона к следующему путем секреции химических веществ — нейротрансмиттеров в области синаптического контакта.
Биполярные нейроны передают сигналы в центральную нервную систему (ЦНС) через каскады межнейронных связей. Мультиполярные нейроны преобладают в ЦНС позвоночных. Эти клетки имеют единственный аксон и сложную сеть дендритных контактов. Например, мультиполярный аксон двигательного нейрона спинного мозга имеет умеренное количество дендритов — около 10 000. Примерно 2000 этих дендритных контактов приходится на тело клетки и около 8000 — соединяется с дендритным деревом. Клетка Пуркинье мозжечка, самыйбольшой мультиполярный нейрон, имеет около 150 000 дендритных контактов.
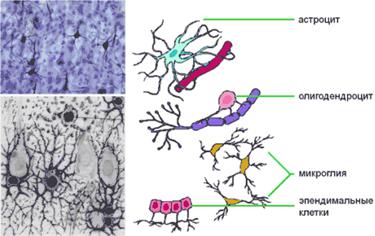
Второй крупный класс клеток нервной системы составляют клетки глии. В ЦНС клеток глии позвоночных примерно в 10-15 раз больше, чем нейронов. Клетки олигодендроглии, шванновские клетки и астроциты — три основных типа глиальных клеток. В ЦНС клетки глии выполняют самые разнообразные функции, в том числе:
I - служат опорными элементами для нейронов, разделяют и изолируют нейроны друг от друга;
II - продуцируют миелин, служащий изолирующей оболочкой и необходимый для некоторых видов нейронов;
III - выполняют роль «уборщиков», удаляя остатки клеток;
IV - участвуют в формировании непроницаемого клеточного барьера между головным мозгом и капиллярами, так называемого гематоэнцефалического барьера.
Клетки глии выполняют и другие важные функции в ЦНС, которые пока не изучены до конца.
При морфологическом, биохимическом и функциональном разнообразии клеток, которое было представлено выше клетки имеют основные структуры:
- ядро;
- цитоплазма;
- оболочка (плазматическая мембрана и гликокаликс)

1. Клетки, синтезирующие белки (синтетический тип) — фибробласты; эпителиоциты некоторых желез; нейроны; плазмоциты, секретирующие антитела.
При световой микроскопии активность этих клеток оценивают по следующим критериям: размер ядра, состояние хроматина, количество ядрышек; насыщенность цитоплазмы РНК — базофилия цитоплазмы при окрашивании гематоксилином и эозином, положительная реакция на пиронин (метод Браше); для нейронов используется метод Ниссля.
Типичными ультрамикроскопическими признаками клеток, активно синтезирующих белки, является крупное ядро с превалированием эухроматина (где происходит транскрипция с образованием мРНК), наличие ядрышек (образующих субъединицы рибосом) вблизи кариолеммы, богатой ядерными порами (для эффективного транспорта между ядром и цитоплазмой. Типичный набор органелл в цитоплазме включает развитую гранулярную эндоплазматическую сеть и комплекс Гольджи. Плазмолемма неровная с инвагинациями и признаками везикуляции, может иметь зоны адгезивных и специализированных контактов.
В некоторых случаях для визуализации белок-синтезирующих клеток используют иммуноцитохимическое исследование, направленное на выявление специфического секреторного продукта — например, гормона или муцинов.
2. Клетки, участвующие в метаболизме липидов (гепатоциты, адипоциты) и/или продуцирующие стероидные гормоны (эпителий коркового вещества надпочечников, клетки гранулезы фолликулов и желтого тела яичника, гландулоцит яичка (клетки Лейдига)).
Типичными особенностями этих клеток является активное ядро (синтез ферментов) с ядрышками. В цитоплазме развиты органеллы, принимающие участие в метаболизме липидов и углеводов — гладкая эндоплазматическая сеть, комплекс Гольджи, митохондрии (с тубуло-везикулярными кристами при синтезе стероидов); включения липидов и/или гликогена. Поверхность таких клеток, как правило, имеет микроворсинки, повышающие обменную площадь плазмолеммы.
3. Клетки, выполняющие функцию детоксикации и цитопротекции (гепатоциты, клетки Клара, пигментные клетки).
Имеют развитую гладкую эндоплазматическую сеть, в составе которой — ферменты антиоксидантной системы, пероксисомы, протеасомы, цитоплазма, богатая белками теплового шока. В пигментных клетках (меланоциты, пигментный эпителий глаза) могут присутствовать специализированные структуры — меланосомы, защищающие клетки от ультрафиолетового излучения.
4. Фагоцитирующие клетки (нейтрофилы, клетки системы фагоцитирующих мононуклеаров).
Способны к распознаванию и захвату веществ, частиц, микроорганизмов. Плазмолемма таких клеток неровная с инвагинациями и выростами. Цитоплазма богата лизосомами, образующимися за счет комплекса Гольджи, развита система эндосом. Для выявления таких клеток и оценки их активности используют цитохимические, иммуноцитохимические методы и электронную микроскопию.
5. Транспортирующие клетки (эпителий кишки, почки, цилиарный эпителией глаза, эпендимоциты, клетки исчерченных протоков слюнных желез, эпителий слизистой оболочки желчного пузыря, эндотелий сосудов).
Типичной особенностью является полярность - базальный полюс содержит ядро, фиксирован к базальной мембране, плазмолемма формирует инвагинации и складки, между которыми расположены митохондрии. Апикальный полюс клетки имеет многочисленные микроворсинки и пиноцитозные пузырьки. Селективность транспорта достигается закрытием межклеточных пространств с помощью плотных контактов (обеспечивающих химическую изоляцию) и адгезивных поясков и десмосом (обеспечивающих механическую связь между клетками).
6. Клетки с высокой механической резистентностью (эпителиоциты многослойных эпителиев).
Выполнение такой функции возможно за счет специализации плазмолеммы, мощного цитоскелета и развитых межклеточных контактов. Для плазмолеммы характерно наличие особых белков подмембранного слоя, обеспечивающих увеличение ее толщины в 3 — 5 раз (инволюкрин, кертолинин и пр.). В структуре цитоскелета доминирующие элементы — это промежуточные филаменты, которые могут объединяться и формировать фибриллы, связанные между собой белком филлагрином. Между клетками развиты контакты, обеспечивающие механическую связь — десмосомы, адгезивные соединения, информационный обмен — щелевые контакты. Мощный цитоскелет присутствует также в нейронах, имеющих отростки.
7. Клетки, способные к сокращению (гладкие миоциты, кардиомиоциты, скелетная мышечная ткань, миоэпителиоцигы, миофибробласты).
В цитоплазме таких структур развит сократительный аппарат — миофиламенты, которые могут формировать миофибриллы. Их наличие в цитоплазме определяет феномен поперечной исчерченности. Поскольку сокращение — это энергозависимый процесс, элементы цитоскелета всегда связаны с митохондриями, а также с цистернами гладкой эндоплазматической сети, которые депонируют Са2+. Иммуноцитохимическое выявление сократимых клеток основано на определении белков миофиламентов — актина, тропонина, тропомиозина, миозина. Кроме того, ряд нарушений процесса сокращения связан с изменением опорных структур и белков плазмолеммы, что определяет интерес к выявлению десмина, дистрофина и пр.
Это далеко не полный перечень функциональных типов клеток, ряд клеток совмещает в себе структуры и функции нескольких типов. Примером могут быть сенсорные клетки органов чувств, механорецепторные клетки и пр.
Лекция № 2 Органоиды клетки
Органоиды клетки
Органоиды, или органеллы, - постоянные специфические структуры цитоплазмы, выполняющие определенные функции, необходимые для поддержания жизнедеятельности клетки.
Различают органоиды общего значения и специальные органоиды. Органоиды общего значения имеются во всех клетках и выполняют функции, общие для всех клеток – митохондрии, рибосомы, эндоплазматическая сеть, комплекс Гольджи, лизосомы, пероксисомы, цитоскелет и клеточный центр.
Органоиды специального значения имеются только в клетках какого-то определенного типа и обеспечивают выполнение функций, присущих только этим клеткам.
Мембранные органоиды:
- ядро;
- эндоплазматическая сеть;
- аппарат Гольджи;
- митохондии;
- лизосомы;
- пероксисомы;
- вакуоли.
Немембранные органоиды
- рибосомы;
- клеточный центр;
- цитоскелет.
Эндоплазматическая сеть открыта Портером в 1945 году. Морфология – сеть канальцев и цистерн сложенных мембранами. Различают гранулярную (шероховатую, зернистую) и гладкую ЭПС.

Гранулярная ЭПС содержит рибосомы на наружной стороне мембраны, обращенную к гиалоплазме. Гладкая ЭПС не содержит рибосомы. В скелетных мышцах ЭПС носит название саркоплазматический ретикулум. ЭПС пронизывает всю клетку. Полость ЭПС сообщается с перинуклеарным пространством ядра, а мембрана ЭПС с плазматической мембраной ядра. На рибосомах гранулярной ЭПС синтезируются секреторные белки, предназначенные для выведения из клетки, а также белки лизосом и внеклеточного матрикса.
Синтез указанных белков включает 5 этапов:
1. Связывание сигнал, распознающий частицы (SRP), состоящий из 6-ти белков и малой цитоплазматической РНК (srpРНК) с сигнальной последовательность полипептидной цепи.
2. Взаимодействие комплекса матричной РНК (мРНК) – рибосома - SRP со специфическим белком-рецептором (SRP-рецептором), находящимся на цитоплазматической стороне мембраны.
3. Освобождение SRP от комплекса; связывание рибосомы с белок-транслоцирующим комплексом мембраны шероховатой ЭПС транслоконом и встраивание сигнальной последовательности в канал транслокона.
4. Возобновление трансляции и перенос растущей полипептидной цепи в полость ЭПС.
5. Удаление сигнальной последовательности от полипептида под действием фермента.
 В полости ЭПС синтезированные водорастворимые белки подвергаются фолдингу и гликозилированию путём присоединения к белкам однотипных олигосахаридных цепочек. В результате фолдинга белки приобретают специфическую конформацию, и в составе транспортных пузырьков, отделяющихся от мембраны ЭПС, транспортируются в комплекс Гольджи, где подвергаются дальнейшим модификациям и сортировке.
В полости ЭПС синтезированные водорастворимые белки подвергаются фолдингу и гликозилированию путём присоединения к белкам однотипных олигосахаридных цепочек. В результате фолдинга белки приобретают специфическую конформацию, и в составе транспортных пузырьков, отделяющихся от мембраны ЭПС, транспортируются в комплекс Гольджи, где подвергаются дальнейшим модификациям и сортировке.
Наряду с секреторными белками на гранулярной ЭПС синтезируется большая часть полуинтегральных и интегральных белков. В ЭПС происходит также синтез мембраны липидов и осуществляется «сборка» компонентов мембраны. При этом увеличения общей площади мембранной ЭПС не происходит. Это связано с тем, что некоторые фрагменты мембраны ЭПС постоянно утрачиваются в составе отделяющихся от нее транспортных пузырьков, которые переносят их в аппарат Гольджи и далее в другие мембранные структуры клетки:
- плазмалемму;
- лизосомы.
Кроме того, ЭПС, как считают, участвуют в образовании пероксисом. Таким образом, грЭПС служит «фабрикой» мембран для плазмалеммы, аппарата Гольджи, лизосом и других мембранных структур клетки. ЭПС обеспечивает также сегрегацию (разделение) белков, предназначенных на экспорт, и лизосомных гидролаз от литозольных белков.
Агранулярная (гладкая) эндоплазматическая сеть представляет собой замкнутую сеть трубочек, канальцев, цистерн и везикулярных образований размером 50 —100 нм и образуется, по-видимому, из гранулярной ЭПС.
На цитоплазматической поверхности гладкой ЭПС синтезируются жирные кислоты, холестерол и большая часть липидов клетки, в том числе почти все липиды, необходимые для построения клеточных мембран. Поэтому гладкую ЭПС нередко называют «фабрикой липидов». В клетках печени (гепатоцитах) с мембранами гладкого эндоплазматического ретикулума связан фермент, обеспечивающий образование глюкозы из глюкозо-6-фосфата. Эта реакция имеет большое значение в поддержании уровня глюкозы в организме человека. Кроме того, гладкая эндоплазматическая сеть осуществляет детоксикацию (обезвреживание) ксенобиотиков, а также ядовитых веществ, образующихся в процессе метаболизма и подлежащих удалению из организма. Большую роль в процессах детоксикации играют ферменты семейства цитохрома Р450, катализирующие реакции гидроксилирования или микросомального окисления. В результате указанных реакций липофильные (нерастворимые) вещества становятся более гидрофильными (растворимыми), заключаются в другие реакции и затем выводятся из организма.
Помимо указанных основных функций, гладкая эндоплазматическая сеть выполняет ряд дополнительных. В ее мембранах локализуются Са2+ - насосы, активно закачивающие ионы Са2+ из цитозоля в полость саркоплазматической сети. Поэтому концентрация ионов Са2+ в полости гладкой эндоплазматической сети достигает 10-3 моль/л, т.е. почти в 10 000 раз выше, чем в цитозоле. Под действием химических сигналов ионы Са2+ переносятся из полости гладкой ЭПС через ионные каналы, встроенные в ее мембрану, в гиалоплазму, где связываются с определенными белками, влияющими на множество внутриклеточных процессов: активацию или инактивацию ферментов, экспрессию генов, освобождение антител из клеток иммунной системы и др.
В мышечных волокнах гладкая ЭПС имеет структурные и функциональные особенности и называется саркоплазматической сетью. При возбуждении плазмалеммы выход ионов Са2 из гладкой эндоплазматической сети в цитозоль стимулирует сокращение миофибрилл.
В организме человека эндоплазматическая сеть особенно хорошо развита в клетках, синтезирующих гормоны, в клетках печени (гепатоцитах) и некоторых клетках почек.
Комплекс Гольджи (КГ), или аппарат Гольджи, - пластинчатый комплекс, расположен вблизи ядра, между ЭПС и плазмалеммой. Его структурнофункциональная единица — диктиосома представляет собой стопку из 5—20 плоских одномембранных мешочков (цистерн), имеющих диаметр около 1 мкм, внутренние полости которых не сообщаются друг с другом. Количество таких мешочков в стопке обычно не превышает 5-10, а расстояние между ними составляет 20—25 нм.
В каждой диктиосоме различают три части, проксималъную (cis-полюс), обращенную к ЭПС, медиальную и дистальную (trans- полюс), обращенную к плазмалемме. К дистальной цистерне транс-полюса примыкают многочисленные трубочки и пузырьки, образующие транс-сеть аппарата Гольджи.

Белки, синтезированные на шероховатой эндоплазматической сети в составе отпочковывающихся от нее мембранных пузырьков, покрытых клатрином, транспортируются к цис-полюсу аппарата Гольджи. В аппарате Гольджи осуществляется химическая модификация транспортируемых белков. Этот процесс происходит поэтапно по мере транспорта белков от цис- к транс-полюсу аппарата Гольджи, каждая цистерна которого содержит характерный для нее набор ферментов. Гликопротеины, несущие первоначально одинаковые олигосахаридные цепочки, в проксимальных цистернах подвергаются последовательной перестройке, специфической для белков каждого вида. В процессе гликозилирования одни олигосахаридные остатки удаляются, другие добавляются с образованием дополнительных ветвей олигосахаридов. В аппарате Гольджи некоторых специализированных клеток синтезируются полисахариды, которые, соединяясь с белками, образуют протеогликаны, составляющие основу вещества межклеточного матрикса. Таким образом, химическая перестройка белков в цистернах цис-полюса аппарата Гольджи осуществляется главным образом путем их гликозилирования, тогда как в транс-полюсе происходит в основном сульфатирование и фосфорилирование белков. Некоторые белки в комплексе Гольджи подвергаются протеолитическому расщеплению, в результате чего первоначально неактивные молекулы приобретают биологическую активность. Например, таким путем в клетках поджелудочной железы образуется инсулин.
В комплексе Гольджи протекают реакции синтеза мембранных гликолипидов. При этом углеводные компоненты в молекулах синтезированных мембранных гликолипидов, также как и у гликопротеинов, всегда оказываются обращенными в полость цистерн аппарата Гольджи. Этим объясняется топология углеводных остатков молекул мембранных гликолипидов и гликопротеинов при включении их в состав плазмалеммы, где они участвуют образовании гликокаликса.
В транс-сети осуществляется сортировка белков и упаковка их в мембранные пузырьки, покрытые клатрином. Белки, предназначенные для экспорта, заключаются в одни пузырьки, лизосомные белки — в другие, мембранные белки — в третьи. В специальные пузырьки упаковываются также белки, присущие ЭПС, например ферменты фолдинга, которые случайно оказались в комплексе Гольджи.
Сортировка белков осуществляется при помощи встроенных в мембрану транс-сети КГ особых белков-рецепторов, способных специфически взаимодействовать с определенными химическими группировками — маркерами молекул отбираемых белков. В результате участок мембраны, несущей рецепторы, связанные с маркерами отбираемых молекул, обособляется от КГ с образованием мембранного пузырька, покрытого клатрином, транспортирующего нужное вещество к месту назначения. Например, маркером лизосомальных ферментов служит присоединяющийся к ним в комплексе Гольджи олигосахарид, содержащий остаток маннозо-6-фосфата. Для белков ЭПС — ферментов фолдинга и др., которые случайно оказались в КГ, таким маркером служит последовательность из четырех аминокислот, по которой они узнаются и упаковываются в пузырьки, а затем возвращаются обратно в ЭПС. Этот процесс получил название рециклизации.
Секреция экспортных белков осуществляется посредством конститутивной и регулируемой секреции.
В случае конститутивной секреции которая характерна для всех клеток, транспортные пузырьки непрерывно переносятся от аппарата Гольджи к плазмалемме. Таким путем, например, доставляются к поверхности клетки элементы гликокаликса. А также многие продукты, составляющие основу межклеточного вещества. Посредством конститутивной секреции осуществляется также транспорт мембранных белков плазмалеммы.
Регулируемая секреция свойственна лишь специализированным клеткам, секретирующим биологически активные вещества в ответ на действие сигналов, например гормонов.
Таким образом, основными функциями комплекса Гольджи являются химическая модификация, накопление, сортировка, упаковка в секреторные пузырьки и транспорт по назначению белков и липидов, синтезированных в эндоплазматическом ретикулуме. В комплексе Гольджи образуются лизосомы и синтезируются некоторые полисахариды.
Лекция № 1 «Клеточная организация жизни» 9.09.2016 г
Клетка – основное понятие при изучении медико-биологических дисциплин. Термин «Клетка или cellula» ввел Роберт Гук в 1665 году, английский физик и ботаник. Рассматривая при помощи увеличительных стекол срез пробки, обнаружил, что он состоит из ячеек, которые он назвал клетками. Более полутора столетий занял т.н. описательный период, где описывалась морфология животных и растительных клеток.
В 1939 году зоолог Т.Шванн и ботаник М.Шлейден доказали, что все растительные и животные организмы построены из клеток. Впервые это обобщение было сформулировано в 1939 году создателем клеточной теории Т.Шванном, согласно которой, все организмы животных и растений состоят из клеток, каждая клетка организма функционирует независимо от других и все клетки возникают из бесструктурного вещества неживой природы.
В 1855 году выходят работы немецкого врача Рудольфа Вирхова оказавшее большое влияние на развитие клеточной теории. В монографии «Целлюлярная патология» он убедительно доказал, что клетки являются постоянными структурами, и возникают только путём размножения себе подобных, – «всякая клетка происходит от другой клетки. Там где возникает клетка, ей должна предшествовать клетка, подобная тому, как животное происходит только от животного, растение только от растения». Т.е. Вирхов опроверг ошибочное представление Т.Шванна об образовании клеток из бесструктурного вещества и обосновал одно из ключевых положений клеточной теории «каждая клетки из клетки» (omnis cellula e cellula). В этой работе он также обосновал положение что патология формируется на клеточном уровне, что в последствии бы доказано развитием медицины, а именно патология возникает на клеточном уровне.
Карл Бэр обосновал положение – клетка является не только единицей строения, но и единицей развития живых организмов на основе работ по яйцеклеткам млекопитающим.

Создание клеточной теории – новой этап в развитии биологии. В современной трактовке клеточная теория включает следующие основные положения:
1. Клетка - элементарная единица живого, структурная и функциональная единица жизни.
2. Клетки всех организмов сходны (гомологичны) по строению, выполняемым функциям, химическому составу, обмену веществ, что объясняется единством их происхождения.
3. Размножение клеток происходит путём их деления, и каждая новая клетка образуется в результате деления исходной (материнской) клетки. Клетка - элементарная единица размножения.
4. В многоклеточных организмах клетки специализированы по выполняемым функциям, объединены целостной системой тканей и органов, связанные между собой нейрогуморальными формами регуляции. Благодаря деятельности клеток в многоклеточных организмах осуществляется рост, развитие, обмен веществ и энергии.
5. Клетки многоклеточных организмов тотипотентны, т.е. обладают генетическими потенциями всех клеток живого организма, но отличаются друг от друга разной экспрессией (активностью) различных генов, дифференцировкой, что приводит к морфологическому и функциональному разнообразию клеток. (Задание на дом: пояснить основные положения клеточной теории).
Клетка - упорядоченная, структурированная, открытая система, т.к. именно ей присуща самое главное свойство живого - обмен веществ – пластический, энергический, информационный с другими клетками многоклеточного организма и внешней средой.


Клетка – характеризуется биологическими свойствами:
- саморегуляция;
- самовоспроизведение;
- самообновление.
В основе саморегуляции клетки (организма) лежит механизм гомеостаза. Понятие гомеостаза было введено Уолтером, Кэнноном и разработано Клодом Бернаром.
Гомеостаз - относительное постоянство внутренней среды клетки (организма). Различают гомеостаз физиологический и развития.
Гомеостаз физиологический – это генетически детерминированная способность организма сохранять реакции, например: рН крови, осмотическое давление в клетке.
Гомеостаз развития - это генетически детерминированная способность организма сохранять реакции под действием внешней среды, например: при удалении одной почки вторая почка выполняет двойную нагрузку и обеспечивает мочевыделительную функцию.
В основном реакции организма имеют смешанный характер, т.е. гомеостаз и физиологический и развития, например: повышение количества эритроцитов в крови при подъеме в горы. При уменьшении количества кислорода в составе горного воздуха возрастает количество эритроцитов в крови - газообмен остаётся в норме.
В основе самовоспроизведения лежит размножение клеток путём:
- митоза;
- мейоза;
- амитоза;
- эндомитоза.
 Митотически делятся соматические клетки (тело – сома, soma) по формуле: 2n
Митотически делятся соматические клетки (тело – сома, soma) по формуле: 2n
 2n
2n
2n
Разновидностью митоза является эндомитоз, при котором после репликации хромосом не происходят разрушения ядерной оболочки и образование веретена деления. Клетка становится полиплоидной – с увеличенным числом хромосомных наборов. Полиплоидия, в отличие от митоза, осуществляется без снижения специфической функциональной активности клетки и характерна для высокоспециализированных тканей и органов (гепатоциты – печень, кардиомиоциты – сердце).
Путём мейоза в организме человека образуются половые клетки в специализированных железах – половых (яичниках и семенниках) по формуле:
 n
n
n
 2n
2n
n
n
Амитоз - прямое деление соматических клеток путём перетяжки,






