При рассмотрении химического состава растений отмечалось, что углерод составляет почти половину их сухого вещества. Атомы углерода образуют скелет всех органических соединений, а способность их вступать в реакцию с другими элементами обусловливает огромное число этих соединений.
Опыты с водными культурами с большой достоверностью показали, что подавляющую часть углерода растения получают не из почвы. С другой стороны, если поместить растение в атмосферу, очищенную от углекислого газа, например, под стеклянный колпак, в который воздух поступает только через слой натронной извести, то оно начнет обнаруживать признаки голодания. Это свидетельствует о том, что подавляющую часть потребности в углероде растения покрывают за счет углекислого газа атмосферы, составляющего 0,03% общего объёма воздуха.
Добывание растением из углекислого газа воздуха нужного им для
построения своего тела углерода представляет один из важнейших
процессов в жизни растения и всего органического мира. Животные и чело
век не могут усваивать углерод непосредственно из углекислого газа и
для питания употребляют уже выработанные растением органические со-
единения.
Организмы, использующие для питания углекислый газ воздуха, называются автотрофными. Организмы, не способные усваивать углекислый газ воздуха и питающиеся за счет органических веществ, выработанных другими живыми существами, называются гетеротрофными. Кроме животных к ним принадлежат незеленые растения — грибы, бактерии, некоторые высшие растения и др.
Для того чтобы растение могло использовать углекислый газ воздуха, необходимы совершенно определенные условия: свет и наличие хлорофилла. Процесс образования органических веществ из неорганических — углекислого газа и воды, происходящий в зеленых растениях на свету, называется фотосинтезом, или ассимиляцией. Его можно выразить следующим схематическим уравнением:
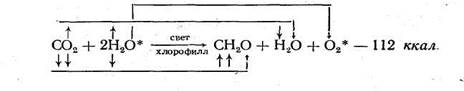
Фотосинтез — это окислительно-восстановительный процесс: от молекулы воды отнимается водород (окисление), который восстанавливает молекулу СО2. Исследования советских (А. Н. Виноградов, Р. В. Тейс) и американских (С. Рубен, М. Камен и др.) ученых показали, что свободный кислород выделяется из молекулы воды, а не из молекулы углекислого газа, как это считалось раньше. Своеобразие фотосинтеза заключается в том, что он, в отличие от подавляющего большинства других процессов, идет с увеличением свободной энергии системы. Поглощенная пигментами солнечная энергия не растрачивается, а накапливается в продуктах реакции в форме потенциальной химической энергии.
ЛИСТ КАК ОРГАН ФОТОСИНТЕЗА.
ХЛОРОПЛАСТЫ
Углекислый газ усваивается в зеленых пластидах клетки — хлоропластах. Следовательно, чтобы послужить материалом для синтеза углеводов, углекислый газ должен быть поглощен клетками, содержащими хлоропласты. Такие клетки составляют основную массу листа — мезофилл. Сверху лист покрыт эпидермой и кутикулой, мало проницаемой для газов. Основной путь, по которому углекислый газ проникает внутрь листа, — устьица. Хотя площадь устьичных отверстий, даже когда они полностью открыты, составляет незначительную часть всей листовой поверхности (не более 1%), диффузия газа через них, в соответствии с законами физики (закон Стефана), идёт с большой скоростью и эпидерма не представляет почти никакого препятствия для проникновения углекислого газа внутрь листа. Мезофилл листа обычно состоит из палисадной (столбчатой) и губчатой паренхимы. Палисадная паренхима расположена на верхней стороне листа и состоит из плотно прилегающих друг к другу клеток, вытянутых перпендикулярно к поверхности листа и богатых хлоропластами. Эту ткань можно рассматривать как ассимиляционную по преимуществу. Форма палисадных клеток способствует оттоку продуктов ассимиляции. Нижние концы клеток примыкают к особым собирающим клеткам, которые в свою очередь сообщаются с проводящими пучками. Благодаря этому устанавливается постоянный ток ассимилятов из листьев в другие органы растения. Губчатая паренхима находится ближе к нижней эпидерме. Клетки ее расположены рыхло и содержат значительно меньше хлоропластов. Считают, что эта ткань облегчает проветривание листа. Достигнув поверхности хлорофиллоносных клеток, углекислый газ растворяется в водe, которая всегда пропитывает их стенки. Затем уже в виде Н2СОз он диффундирует сквозь стенку, проникает в цитоплазму и достигает зеленых пластид, которыми поглощается.
Общая поверхность хлоропластов огромна. Так, в листе бука она примерно в 200 раз превышает его площадь и у столетнего дерева достигает 2 га. Это значительно облегчает растениям добывание углекислого газа из воздуха. Хлоропласты состоят из белково-липидной стромы и пигментов, легко извлекаемых органическими растворителями. Важнейший из пигментов — хлорофилл. Кроме хлорофилла, хлоропласты содержат желтые пигменты — каротиноиды (каротин и ксантофилл). Хлорофилл выполняет функцию поглощения световой энергии и переносит ее на восстановление углекислоты, а также химически участвует в этом процессе. Однако он функционирует только в сочетании со стромой. Строма является носителем ферментов, участвующих в сложных реакциях фотосинтеза. В строме из продукта фотосинтеза – сахара вырабатывается крахмал (первичный или ассимиляционный). Если затенить отдельные участки листа, то на светлом фоне получаются темные фигуры (проба Сакса).

|

|
Рнс.. Обнаружение первичного крахмала при помощи пробы Сакса. А—лист, частично затененный; Б — лист после обработки спиртом и йодом
По химической природе хлорофилл — сложный эфир дикарбоновой кислоты — хлорофиллина и двух спиртов — метилового и фитола р. Хлорофилл содержит четыре соединенных между собой остатка пиррола, которые образуют порфириновое кольцо, центральный атом которого — Mg. По строению хлорофилл весьма близок к красящему веществу крови — гему. В его состав также входит порфириновое кольцо, однако в центре расположен атом Fe. Это сходство было показано Ч. В. Ненцким и польским ученым Л. Мархлевским. К. А. Тимирязев считал установление этого сходства едва ли не самым крупным открытием в области химического изучения хлорофилла.
Исследование многих сотен самых разнообразных видов высших растений показало, что хлорофилл у них совершенно одинаковый. Общее количество хлорофилла в растениях составляет около 1 % от сухого веса. Хлорофилл в хлоропластах находится не в свободном виде, а связан с белком, образуя хлороглобин.
Для образования хлорофилла в растениях необходимо несколько совершенно определенных условий: присутствие пропластид, способных к позеленению, света и солей железа.
Развивающиеся в темноте растения имеют желтый цвет. Они называются этиолированными. Если их выставить на свет, они быстро зеленеют. Считают, что они содержат особое вещество, называемое протохлорофиллом,_образуюшееся в.темноте_и_ под влиянием света_легко_ превращающееся в хлорофилл.
Если выращивать растения при полном отсутствии солей железа, то они тоже будут бледно-желтого цвета и быстро гибнут от истощения. Это явление называется хлорозом. Поскольку железо не входит в состав хлорофилла, считают, что оно служит специфическим катализатором, без которого не осуществляются какие-то подготовительные стадии позеленения. Хлороз часто наблюдается в природе, особенно у растений, растущих на почвах, богатых известью.
Кроме того, у растений иногда наблюдается явление альбинизма — неспособность образовывать хлорофилл даже при самых благоприятных для этого условиях.
УЧАСТИЕ ПИГМЕНТОВ В ПОГЛОЩЕНИИ СВЕТА. РАБОТЫ К. А. ТИМИРЯЗЕВА
Хлорофилл обладает избирательным поглощением световой энергии. Наиболее интенсивно поглощение происходит в красных лучах спектра (длина волны от 650 до 680 ммк) и сине-фиолетовых (длина волны около 470 ммк). Зеленые лучи и часть красных не поглощаются, они и придают хлорофиллу изумрудно-зеленый цвет. Желтые пигменты — каротин и ксантофилл — поглощают свет в зеленой и синей частях спектра.
Энергетическая сторона процесса фотосинтеза глубоко вскрыта и разъяснена в работах К. А. Тимирязева. Он показал, что фотосинтез осуществляется только в лучах спектра, поглощаемых хлорофиллом. Дальнейшие исследования полностью подтвердили это положение. Процесс фотосинтеза в различных частях спектра идет неодинаково. К. А. Тимирязев показал, что максимум ассимиляции приходится на красные лучи, которые несут максимум энергии и полнее всего поглощаются хлорофиллом. В сине-фиолетовых лучах ассимиляция идет слабее, так как они несут меньше энергии. Вопросу о значении в фотосинтезе отдельных частей спектра К. А. Тимирязев придавал большое принципиальное значение. До него господствовало мнение, что свет служит только раздражителем. Этой точки зрения придерживались и современники К. А. Тимирязева немецкие ученые Ю. Сакс и В. Пфеффер. К.А. Тимирязев показал, что свет является источником энергии и необходим для фотосинтеза.
На возбуждение одной молекулы хлорофилла требуется один квант, поэтому в красных лучах, несущих большое число мелких квантов, большее число его молекул перейдет в возбужденное состояние
Кроме избирательного поглощения световой энергии, хлорофилл обладает свойством флуоресценции: в отраженном свете он кажется кроваво-красным, так как отражает поглощенные лучи с изменением длины их волны. Это указывает на значительную фотохимическую активность хлорофилла. Коэффициент поглощенной лучистой энергии в фотосинтезе чрезвычайно низок- 1%- 5%, редко до 10%. Большая часть переходит в тепловую энергию, либо повышает температуру и рассеивается в окружающем пространстве.
ХИМИЗМ ФОТОСИНТЕЗА
Несмотря на простоту суммарного уравнения фотосинтеза, этот процесс отличается чрезвычайной сложностью. Это обусловливается сложностью углеводной молекулы, которая не может сразу возникнуть из таких простых веществ, как СО2 и Н2О; трудностью окисления и восстановления этих прочных соединений; участием в реакциях световой энергии. Исследования показали, что фотосинтез включает не только несколько фотохимических реакций, но и ряд ферментативных, так называемых темновых реакций.
Использование методов меченых атомов (изотопов С, Р, О, N), разделительной хроматографии на бумаге, электрофореза, ионообменной

Рис. 134. Цикл фотосинтетических превращений углерода по Кальвину
очистки и разделения и некоторых других позволило выявить химизм фотосинтеза.
Рядом работ установлено, что первым этапом усвоения углекислоты
является присоединение СО2 к какому-то акцептору (веществу, воспри-
нимающему, присоединяющему другое вещество), карбоксилирова-
ние его:
R • Н + СО2_→ R • СООН.
Таким образом, фотосинтетическому восстановлению подвергается углерод не углекислого газа, а карбоксильной группы. Широкие исследования по выяснению природы первичных акцепторов и путей фотосинтетического превращения углерода были проведены американским ученым Кальвином и его сотрудниками. Схема фотосинтетических превращений углерода, по Кальвину, представлена на рис. 134. Он считает, что процесс фотосинтеза носит циклический и разветвленный характер: одна ветвь этого цикла ведет к образованию прямых устойчивых продуктов фотосинтеза — углеводов, другая имеет циклический характер и приводит к образованию акцептора СО2 — рибулезодифосфата, который вовлекает в фотосинтетический цикл все новые и новые молекулы СО2.
Наряду с этими сложными превращениями углерода за счет энергии света образуются богатые энергией органические соединения фосфора, в. частности аденозинтрифосфат (АТФ). Этот процесс называется фотосинтетическим фосфорилированием:
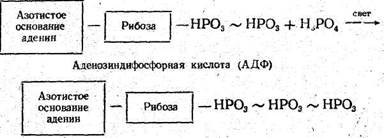
Аденозинтрифосфорная кислота (АТФ)
Энергия макроэргических (богатых энергией, ~) фосфатных связей АТФ идет на восстановительные процессы. При гидролизе макроэргических связей освобождается 7000—16 000 кал на грамм-молекулу.отщепленного фосфата.
К световым реакциям фотосинтеза относится:
1) разложение воды (активированный хлорофилл + 2Н2О-инактивированный хлорофилл + +4Н+2О),
2) фотосинтетическое фосфорилирование,
3) синтез аминокислот и белков.
К темповым реакциям фотосинтеза относятся:
1) фиксация СО2 акцептором,
2) перенос активного водорода на соединение, в котором фиксирована молекула СО2,
3) восстановление акцептора СО2,
4) образование Сахаров.
Приведенное описание фотосинтетического превращения углерода далеко не исчерпывает всей сложности этого процесса. В частности, считают, что акцептором СО2 может быть не только рибулезодифосфат, но и другие соединения.
-
МИНЕРАЛЬНОЕ ПИТАНИЕ РАСТЕНИЙ
ВЕЩЕСТВА, ПОЛУЧАЕМЫЕ РАСТЕНИЯМИ ИЗ ПОЧВЫ
Нет ни одного элемента, который бы не был обнаружен в растениях. Элемент может быть случайной примесью и накапливаться в растениях в больших количествах, или же находиться в нем в ничтожно малом количестве, но безусловно необходим. Установлено, что растение может успешно развиваться, если в питательном растворе находится лишь семь элементов: К, Са, Mg, S, Fe, N и Р. Это мнение держалось в науке более 50 лет, но было установлено, что многие другие элементы играют важную роль в жизни растений. Было установлено, что для нормального роста и развития растений необходимы в ничтожно малых количествах Mg, Zn, Cu, Al, I, Md и др.
Получаемые растениями из почвы вещества по химической природе можно разделить на две группы: металлоиды и металлы.
Металлоиды поступают в растения в виде анионов соответствующих солей. Они необходимы для образования органических веществ. Металлы поступают в растения в виде катионов. Они находятся в клетках в свободном состоянии или слабосвязанном и служат регуляторами жизненных процессов. Например, магний входит в состав хлорофилла, железо и медь- ферментов и т, д.
МЕТАЛЛОИДЫ
Азот. Поступает в растение в виде анионов NO3 и NO2, так и виде катиона NН4. Его значение в жизни растений очень велико.
Фосфор воспринимается растениями в виде анионов солей фосфорной кислоты РО4.В белковые молекулы он входит в той же окисленной форме. В растениях образуются сложные эфиры фосфорной кислоты и фосфатиды которые являются необходимой составляющей цитоплазмы как и белки. Фосфор стоит в центре всего энергетического обмена клетки. Витамины и некоторые ферменты проявляют свое действие лишь в соединении с фосфорной кислотой.
При разложении растительных остатков фосфорная кислота высвобождается в виде неорганических солей и может снова использоваться растениями.
Сера усваивается лишь в виде аниона серной кислоты SO4, источником ее служат растворимые соли. Используется для синтеза белка за счет продуктов фотосинтеза- углеводов, входит в состав горчичных и чесночных масел, участвует в дыхании и росте.
При перегнивании растительных остатков сера отщепляется от белковой молекулы в виде сероводорода, который не усваивается растениями и очень ядовит для корней. В пригодную форму он переводится серобактериями, окисляющими сероводород и серную кислоту.
МЕТАЛЛЫ
Калий содержится в клетках меристемы и молодых органах. Много калия в корнеплодах, клубнях, крахмалистых семенах. Калий обладает большой подвижностью. Из старых, отмирающих органов передвигается в более молодые жизнедеятельные части растения(реутилизация). Калийные удобрения почти всегда оказывают благоприятное воздействие на урожайность сельскохозяйственных культур.
Натрий присутствует в золе растений часто в больших количествах, однако особого значения для жизнедеятельности не имеет и может быть исключен из питательного раствора. Только голофиты- растения, свойственные засоленным почвам, лучше растут в присутствии солей натрия. Из культурных растений к этому типу относятся сахарная свекла, дикий предок которой произрастает на засоленных почвах Средиземного моря.
Магний содержится преимущественно в молодых органах и семенах (до 10-15% золы). Физиологическое действие его близко к действию калия. Магний входит в состав некоторых металлоорганических соединений, в частности хлорофилла, он может активизировать действие некоторых ферментов. Влияние магния зависит от состава почвы. Резко отзываются на внесение магнезиальных удобрений растения, растущие на легких песчаных и супесчаных почвах.
Кальций необходим для роста молодых тканей. Он входит в состав цитоплазматических структур и ядер. Соединения кальция с пектиновыми веществами составляют основу срединных пластинок, склеивающих стенки клеток друг с другом. Многие ферменты активны лишь в присутствии ионов кальция. Он способствует увеличению вязкости цитоплазмы и влияет на поступление веществ в клетку. Одна из важных функций кальция – нейтрализация щавелевой кислоты, образующейся как побочный продукт обмена веществ. При отсутствии кальция наблюдается резкое угнетение корневой системы, Особенно много кальция в старых органах.
МИКРОЭЛЕМЕНТЫ
Микроэлементы необходимы растениям в ничтожно малых количествах и в больших дозах становятся ядовитыми.
Железо участвует в образовании хлорофилла как катализатор. Оно входит в состав окисленных ферментов, играет чрезвычайную роль в процессе дыхания. Возможно железо участвует в процессе фотосинтеза и окислительно-востановительных процессах в клетке как переносчик электронов.
Цинк входит в состав некоторых ферментов. При отсутствии цинка наблюдается угнетение роста молодых проростков, заболевание цитрусовых и тунга.
Марганец активизирует работу многих ферментов, играет большую роль в восстановлении нитратов в растении, влияет на окислительно-востановительные процессы превращения железа. Для нормального роста растениям необходимо ничтожно малое количества марганца, поэтому внесение марганцевых удобрений не всегда дает положительный результат. Отзывчивы на такие удобрения сахарная свекла, хлопчатник, табак и другие культуры.
Бор при его недостатке у растений отмирают точки роста., нарушается расположение элементов ксилемы и флоэмы и полная потеря ими проводимости. Бор благоприятно влияет на цветение и плодоношение. Борные удобрения необходимы на известковый почвах. Очень отзывчивы на эти удобрения сахарная свекла, бобовые травы. При повышенных концентрациях он оказывает угнетающее действие на растения.
Медь оказывает влияние на окислительно-востановительную систему, входит в состав ряда ферментных систем.
При недостатке того или иного элемента у растений наблюдаются определенные симптомы повреждений, по которым можно определить, какого элемента не хватает. Так, хлороз листьев показывает на нехватку железа, отмирание корней- недостаток кальция, отмирание точек роста на недостаток бора. Однако такая диагностика улавливает далеко зашедшую форму голодания, которую внесением удобрений уже не исправить.
ПЕРЕДВИЖЕНИЕ ВЕЩЕСТВ В РАСТЕНИЯХ.
ВОДНЫЙ РЕЖИМ РАСТЕНИЙ. ДЕЙСТВИЕ НА РАСТЕНИЯ ВЫСОКИХ И НИЗКИХ ТЕМПЕРАТУР
ДВА ТОКА ВЕЩЕСТВ В РАСТЕНИИ
Существование организма растения как единого целого, физиологическая взаимосвязь отдельных органов, расположенных в неодинаковых физических средах и выполняющих различные функции, возможны лишь при условии передвижения минеральных и органических веществ.
Факт передвижения веществ в растении по двум направлениям был установлен еще в 1679 г. Мальпиги путем кольцевания. Если удалить со стебля участок коры в виде кольца, то листья на нем остаются живыми и не обнаруживают никаких признаков завядания, а плоды бывают даже больше, чем на не окольцованных ветвях. Это показывает, что передвижение воды и минеральных веществ из почвы осуществляется по древесине (ксилеме). Этот ток веществ был назван восходящим. У верхнего же края кольцевого выреза происходит застой питательных веществ и разрастание тканей коры в виде наплыва. Если наплыв не восстановит удаленную часть коры, то корни отмирают от истощения и все растение погибает. Значит, пластические вещества из листьев в корень передвигаются по коре (в основном по флоэме). Этот ток веществ был назван нисходящим.
Очень долго в науке держалось мнение, что по флоэме передвигаются только органические вещества, а по ксилеме — только вода и минеральные вещества. Однако исследования последних лет с применением метода меченых атомов показали, что по флоэме могут передвигаться не только органические, но и минеральные вещества. В основном это калий, фосфор, частично кальций. Причем эта миграция может осуществляться в любом направлении. После того как было доказано, что в корне тоже синтезируются органические вещества, стало ясно, что и по ксилеме вверх передвигаются не только минеральные, но и органические вещества. Кроме того, установлено, что минеральные и органические вещества из корня могут подниматься вверх по флоэме.
Законы, управляющие передвижением органических веществ, до сих пор изучены мало. Исследования показали, что скорость передвижения органических веществ во много раз больше скорости диффузии, что проводящие пучки отличаются очень интенсивным дыханием и что клетки флоэмы способны не только проводить органические вещества, но и подвергать их различным превращениям. Это позволило сделать предположение, что органические соединения перемещаются по флоэме не вследствие пассивного перетекания растворов или диффузии, а в результате каких-то обменных реакций, непрерывно происходящих в ситовидных трубках между цитоплазмой и передвигающимися молекулами.
ПЕРЕДВИЖЕНИЕ ВОДЫ В РАСТЕНИИ
Путь, который проходит вода в растении, делится на две части: 1) по живым клеткам от корневого волоска до сосудов центрального цилиндра корня и от сосудов листа до клеток мезофилла, испаряющих воду в межклетники; 2) по мертвым клеткам проводящей системы — сосудам и трахеидам.
Путь воды по живым клеткам исчисляется миллиметрами, однако он представляет большие трудности, так как при переходе от одной клетки к другой вода встречает значительное сопротивление, поэтому этим способом вода не может передаваться на большие расстояния. Большую часть пути вода проходит по мертвым, пустым, вытянутым в длину клеткам — трахеидам или же по полым трубкам — сосудам.
Поглощение воды и ее перемещение вверх осуществляется в результате совместного действия следующих факторов: корневого давления (нижний концевой двигатель), транспирации (верхний концевой двигатель), сил сцепления молекул воды.
. Жидкость, вытекающая при плаче растений, называется пасокой. Химический состав ее непостоянен. Весной, когда происходит гидролиз запасных углеводов, она богата сахарами, органическими кислотами и содержит мало минеральных веществ. Выделение капельно-жидкой воды может происходить и через листья, через особые водяные устьица — гидатоды Это явление получило название гуттации. Гуттация происходит в умеренно теплой и влажной, насыщенной водяными парами атмосфере, когда возникает диспропорция между поступлением воды и ее испарением. Наиболее часто она встречается у растений тропической и субтропической зон и иногда происходит с такой силой, что создает впечатление дождя. Из растений умеренной зоны активно гуттируют ива, картофель, гречиха и др. Плач растений и гуттация не являются только осмотическими процессами, так как они прекращаются при действии на корни веществами, ингибирующими дыхание. Прежде чем попасть в сосуд корня, вода, поглощенная корневым волоском, должна проделать путь по живым клеткам коровой паренхимы. Согласно Д. А. Сабинину, такой односторонний ток воды возможен только при различии в обмене веществ в разных частях клетки, при котором на одном полюсе клетки образуется больше тургорогенных веществ, чем на другом, а следовательно, возникают большее осмотическое давление и большая сосущая сила. Поступление воды из клеток в сосуды происходит вследствие того, что раствор, находящийся в сосудах, обладает большей сосущей силой, чем близлежащие клетки. Сосуды — это мертвые клетки без цитоплазмы и их сосущая сила равна всей величине осмотического давления раствора (S = P), в то время как в живых клетках существует еще тургорное давление и S = P—Т. Вода, находящаяся в сосудах и трахеидах, имеет вид тонких нитей, которые своими нижними концами упираются в паренхимные клетки корня, а верхними — как бы подвешены к испаряющим клеткам листа. Для того чтобы вода передвигалась вверх, необходимо, чтобы испаряющие клетки обладали достаточной сосущей силой, которая тем больше, чем сильнее испарение. В клетках листьев древесных растений она достигает 10— 15 атм.
Однако, как показал русский ученый Е. Ф. Вотчал, поднятие воды на большую высоту по сосудам возможно лишь при условии существования непрерывных водных нитей, которое обеспечивается силами сцепления молекул воды между собой и со стенками сосудов. Сила сцепления достигает 300—350 атм.
ТРАНСПИРАЦИЯ
Испарение воды растением не только чисто физический, но и физиологический процесс, так как на него большое влияние оказывают анатомические и физиологические особенности растения. Этот процесс назван тринспирацией.
Испарение воды в листе происходит с поверхности клеток мезофилла, По вычислениям Тэррела, эта поверхность у растений умеренно влажных местообитаний в 12—19 раз превышает наружную поверхность листьев, а у растений засушливых местообитаний — в 17—30 раз. Парообразная вода попадает в межклетники и через устьичные щели диффундирует наружу. Такая транспирация называется устьичной. Площадь устьичных щелей составляет около 1 % общей площади листа. Однако, как уже отмечалось в отношении диффузии газа при фотосинтезе, диффузия пара через устьица идет с той же скоростью, с какой она шла бы при отсутствии эпидермы. Одна из важнейших особенностей устьичного аппарата — способность открывать и закрывать устьичные отверстия. Стенка замыкающих клеток имеет неодинаковую толщину: часть стенки, примыкающая к щели, значительно утолщена, в то время как остальная часть стенки остается тонкой. Это приводит к тому, что при насасывании воды тонкая наружная часть стенки растягивается значительно больше, чем толстая, кривизна клеток увеличивается, и щель раскрывается. При уменьшении объема замыкающей клетки тонкая стенка распрямляется, и щель закрывается. В основе процесса, обусловливающего изменение тургора в замыкающих клетках, лежит превращение крахмала в сахар и обратно, которое вызывается изменением хода ферментативных реакций. Большое влияние на открывание устьиц 
Рис. 143. Изменение транспирации в зависимости от изменения основных метеорологических показателей в течение дня:
/ — обшая солнечная радиация, 2 — дефицит насыщения, 3 — температура, 4 — интенсивность
транспирации
оказывает свет. На свету устьица закрываются лишь с большим трудом. Это фотоактивное открывание устьиц носит приспособительный характер: через устьица проникает в лист углекислый газ и для процесса фотосинтеза необходимо, чтобы устьица были открыты в светлые часы суток. Пользуясь различными методами, можно проследить за ходом устьичных движений в течение суток. В ясную не очень жаркую и сухую погоду у большинства растений устьичные щели открываются на рассвете, наиболее широко они раскрыты в утренние часы, к полудню они начинают сужаться и закрываются несколько раньше захода солнца. В сухую и жаркую погоду устьичные щели к полудню закрываются полностью, а к вечеру опять открываются. У разных растений устьица ведут себя неодинаково. Так, у картофеля, капусты и некоторых других растений устьица обычно открыты круглые сутки, у хлебных злаков устьица на ночь закрываются. Большинство растений в этом отношении занимает промежуточное положение. Движение устьичного аппарата в зависимости от внешних условий весьма сложны и не всегда поддаются учету.
Наряду с устьицами в испарении воды участвует и вся поверхность листьев, несмотря на то, что она покрыта кутикулой. Эта форма транспирации называется кутикулярной. У взрослых листьев кутикулярная транспирация в 10—20 раз слабее, чем устьичная.
Транспирация имеет следующее значение: 1) создает непрерывный ток воды, 2) облегчает передвижение минеральных веществ от корня к листьям, 3) защищает листья от перегрева.
Количество воды, которое растение пропускает через себя, огромно. Одно растение подсолнечника или кукурузы за вегетационный период испаряет более 200 кг воды.
Транспирация зависит от метеорологических условий: температуры воздуха, света, ветра, дефицита насыщения воздуха парами воды, а также от количества воды в растении. В результате получаются те сложные кривые, которые характеризуют суточный ход этого процесса в природе (рис. 143). Опыты показали, что для нормального развития растения не нуждаются в том огромном количестве воды, которое они теряют в естественных условиях и что транспирация очень часто может быть сокращена с пользой для них. Так, наиболее пышное развитие растений наблюдается во влажном тропическом климате, где влажность почвы и воздуха очень высоки. В оранжереях растения растут лучше, если поддерживать влажность на возможно более высоком уровне. Даже в полевой культуре применяют освежительные поливы дождеванием в целях повышения влажности воздуха и снижения транспирации.
ВОДНЫЙ БАЛАНС РАСТЕНИЙ
Растения, живущие на суше, должны поддерживать цитоплазму клеток в достаточно насыщенном водой состоянии. Поэтому они имеют ряд особенностей в своем строении, которые, с одной стороны, обеспечивают снижение количества теряемой воды (кутикула, покрывающая все надземные части, восковой налет, волоски и т. д.), а с другой — быструю подачу воды из почвы к листьям (мощная корневая система, хорошо развитая проводящая система и т. д.). В то же время, чтобы успешно шел процесс фотосинтеза, необходим тесный контакт хлорофиллоносных клеток с окружающей атмосферой. Это приводит к непрерывному испарению воды клетками, которое усиливается нагреванием листа вследствие поглощения хлорофиллом солнечной энергии, также необходимой для фотосинтеза. Это глубокое внутреннее противоречие между углеродным питанием и водным режимом К. А. Тимирязев назвал «необходимым злом», так как в условиях засухи оно может привести к гибели растения. Это противоречие кладет глубокий отпечаток на строение растений и на всю их жизнедеятельность.
Одно из важнейших условий нормального функционирования высших наземных растений — сведение водного баланса, т. е. соотношения между приходом и расходом воды, без длительного и глубокого дефицита. В умеренно влажные и не слишком жаркие дни это условие выдерживается. Но в ясные летние дни к полудню транспирация настолько увеличивается, что возникает водный дефицит, который при достаточной влажности почвы достигает 5—10%, а при недостатке влаги в почве возрастает до 25% и более. Это вполне нормальное явление. Дальнейшему увеличению водного дефицита препятствует способность растений под влиянием потери воды регулировать свою транспирацию в довольно широких пределах.
Однако эта регуляция имеет свои границы и при значительном возрастании транспирации и иссушении почвы наступает резкое нарушение водного баланса, которое внешне выражается в завядании. При этом клетки теряют тургор, листья, и молодые побеги повисают вниз. Завядаиие еще не означает утрату растением жизнедеятельности. Если растение своевременно снабдить водой, то тургор восстанавливается. Различают два типа завядания растений: временное и длительное. Первое наблюдается при сильном возрастании транспирации, когда поступающая из почвы вода не успевает покрывать ее трату. При этом листья, больше всего расходующие воду, теряют тургор и вянут, а остальные органы растения содержат еще достаточное количество воды. При ослаблении транспирации, например к вечеру, водный дефицит исчезает и растение оправляется без добавочного увлажнения почвы. Большого вреда растениям временное завядание не приносит, но все же снижает урожай, так как приостанавливает фотосинтез и рост. Длительное завядание наблюдается, когда почва не содержит достаточного количества доступной для растений воды. При этом водный дефицит за ночь не исчезает, и к утру: растения оказываются не вполне насыщенными водой и не в состоянии нормально функционировать. В этих условиях тургор постепенно падает во всех органах растения, вплоть до корневых волосков, так как завядшие листья, обладая большой сосущей силой, оттягивают от них воду. Корневые волоски отмирают, поэтому даже при обильном поливе растения восстанавливают прежнюю скорость водоснабжения только через несколько дней, когда образуются новые корневые волоски. Исследования Н. А. Максимова, Н. М. Сисакяна и других показали, что завядание вызывает глубокое влияние на состояние биоколлоидов клетки, которое ведет к нарушению обмена веществ. Процессы гидролиза усиливаются, синтетические процессы задерживаются. Это отражается на всех физиологических функциях растения — фотосинтезе, дыхании, передвижении веществ, росте и т. д. Урожай падает, зерно получается щуплым. Длительное завядание вызывает необратимые изменения, и клетки в конце концов отмирают даже при возобновлении водоснабжения. В то же время завядание — весьма действенный способ задержать транспирацию в наиболее опасные для растения периоды. В завядшем состоянии потеря воды растением в 5—10 раз меньше, чем в благоприятном периоде.
У разных растений завядание наступает при потере неодинакового количества воды. Так, подсолнечник и картофель не завядают при потере 25—30% воды, а другие растения, особенно теневые, завядают уже при потере 2—3% воды. Соотношение между добыванием и расходом воды зависит от многих факторов. Это обусловливает чрезвычайное разнообразие типов наземных растений по отношению к водному режиму.
ДЕЙСТВИЕ НА РАСТЕНИЯ НЕДОСТАТКА ВЛАГИ
И ВЫСОКИХ ТЕМПЕРАТУР.
ЗАСУХОУСТОЙЧИВОСТЬ И ЖАРОСТОЙКОСТЬ
Засуха — это резкое проявление недостатка влаги, приводящее к нарушению водного режима растений. Засуха бывает атмосферная и почвенная. Атмосферная засуха характеризуется высокой температурой и малой относительной влажностью воздуха (10—20%). Она приводит к завяданию растений. Сопровождающая атмосферную засуху высокая температура воздуха вызывает сильный нагрев растений. Большие повреждения растениям наносят суховеи — очень сухие горячие ветры. При этом высыхает и отмирает значительная часть листьев. При атмосферной засухе корневая система остается неповрежденной. При большой продолжительности атмосферная засуха вызывает иссушение почвы — почвенную засуху. Она более опасна для растения, так как приводит к длительному завяданию. Уже отмечалось, что завядание растений нарушает обмен веществ и значительно снижает урожайность.
Разные части растений реагируют на засуху неодинаково. Так, уменьшение содержания воды в листьях приводит к повышению их сосущей силы, и они начинают отсасывать воду от конусов нарастания стебля, бутонов и завязавшихся плодов. Это вызывает гибель цветков или их стерильность, образование щуплого зерна — захват. Верхние листья дольше сохраняют свою жизнедеятельность при засухе, чем нижние, так как оттягивают от них воду. Эта особенность верхних листьев объясняется тем, что они находятся в условиях несколько затрудненного водос






