Определение понятия. Понятие о тканях как группах сходных клеток появилось уже в трудах первых ботаников-анатомов в XVII в. Мальпиги и Грю описали важнейшие ткани, в частности ввели понятия о паренхиме и прозенхиме, основанные на форме составляющих их клеток. Эти термины оказались удобными для чисто описательных целей и удержались в современной анатомии.
Классификация тканей на основе физиологических функций была разработана в конце XIX— начале XX в. Швенденером и Габерландтом и сыграла важную роль в развитии анатомии растений, особенно одного из ее разделов — физиологической анатомии.
Однако принятие лишь одного принципа — функционального — неизбежно приводит к большим противоречиям и затруднениям по следующим причинам.
1. Большинство растительных тканей многофункционально, т. е. одна и та
же ткань может выполнять две или три функции (например, функции
проведения и укрепления, или функции ассимиляции, укрепления и запасания
веществ и т.д.). Кроме того, одна ткань может состоять из разнородных
элементов, выполняющих разные функции. В таком случае эту ткань называ-
ют сложной, в противоположность простой ткани, состоящей только
из однородных элементов.
2. Часто ткань с возрастом меняет функции или сохраняет лишь неко-
торые из первоначальных (древесина в начале своего существо-
вания выполняет преимущественно водопроводящую функцию, а потом ее те-
ряет и служит только для укрепления растения).
3. Клетки, сходные по строению и функции, т. е. относящиеся к одной
ткани, могут быть рассеяны поодиночке среди клеток других тканей. Примером могут
служить толстостенные опорные клетки в паренхиме цветоножек кубышки
или клетки с эфирными маслами в листе пеперомии.
Тканями называют устойчивые, т. е. закономерно повторяющиеся, комплексы клеток, сходные по происхождению, строению и приспособленные к выполнению одной или нескольких функций.
Так, у бактерий и примитивных водорослей все клетки одинаковы. У прочих водорослей число различных типов клеток также очень невелико, и у наиболее сложных бурых водорослей не превышает 10. У мхов уже насчитывается около 20, у папоротникообразных — около 40, а у покрытосеменных растений — около 80 различных типов клеток.
Классификация тканей. Ткани удобно рассмотреть в следующей системе.
I. Образовательные ткани (меристемы):
1) верхушечные (апикальные);
2) боковые (латеральные):
3) вставочные (интеркалярные);
4) раневые (травматические).
II. Ассимиляционные ткани,
III. Запасающие ткани.
IV. Аэренхима.
V. Всасывающие ткани:
VI. Покровные ткани:
1. первичная (эпидерма)
2) вторичная (перидерма);
3) третичная (корка, или ритидом).
VII. Ткани, регулирующие прохождение веществ:
1) эндодерма;
2) экзодерма.
VIII. Выделительные ткани:
1) наружные:
а) железистые волоски (трихомы) и выросты (эмергенцы),
б) нектарники,
в) гидатоды;
2) внутренние:
а) выделительные клетки (с эфирными маслами, смолами, кристаллами, танинами и т.д.),
б) многоклеточные вместилища выделений,
в) смоляные каналы (смоляные ходы),
г) млечники (членистые и нечленистые).
IX. Механические ткани (опорные, скелетные, арматурные):
1) колленхима;
2) склеренхима:
а) волокна,
б) склереиды.
X. Проводящие ткани:
1) ксилема (древесина);
2) флоэма (луб).
Все ткани за вычетом меристем называют постоянными тканями.
ОБРАЗОВАТЕЛЬНЫЕ ТКАНИ
Общие представления о меристемах. Растение растет всю жизнь. Длительный рост с образованием новых органов и тканей — характерная черта, отличающая растения от животных. Такой рост возможен благодаря наличию образовательных тканей, или меристем.
Меристемы состоят из недифференцированных и по внешнему виду одинаковых клеток, способных многократно делиться. Возникающие из меристем клетки дифференцируются, т. е. становятся различными и дают начало всем тканям и органам растения.
Меристемы могут сохраняться очень долго, в течение всей жизни растения (у некоторых деревьев тысячи лет), так как содержат некоторое число клеток (инициальных), способных делиться неопределенное число раз с сохранением меристематического характера
Распределение меристем в теле растения. У высших растений характерное распределение меристем устанавливается очень рано, уже на первых этапах развития организма. Оплодотворенная яйцеклетка делится и образует предзародыш (проэмбрио), составленный эмбриональными, т.е. меристематическими, клетками. На двух противоположных полюсах — на кончике зародышевого корешка и в первичной почечке — локализуются группы инициальных клеток, формирующих верхушечные (апикальные) меристемы. Эти меристемы наращивают корень и побег в длину. При ветвлении каждый боковой побег и каждый боковой корень также обязательно имеют верхушечные меристемы со своими инициалями.
Боковые (латеральные: лат. lateralis — боковой) меристемы в осевых органах (стеблях, корнях) образуют цилиндрические слои, на поперечных разрезах имеющие вид колец. Одни из боковых меристем возникают непосредственно под апексами и в тесной связи с апикальными меристемами. Боковые меристемы (камбий, феллоген) возникают позже, из клеток постоянной ткани путем их дедифференциации, т. е. обратного преобразования в меристему. Часто (например, у большинства однодольных растений) вторичные меристемы отсутствуют, и тогда все тело растения образовано одними, лишь первичными меристемами.
Молодые ткани формируются от основания к верхушке
Вставочные меристемы представляют собой активно растущие участки, расположенные у оснований междоузлий. Их можно назвать остаточными, поскольку они происходят от верхушечных меристем, но их преобразование в постоянные ткани задерживается по сравнению с соседними участками стебля.
От верхушечных и боковых меристем вставочные отличаются тем, что, во-первых, в них имеются некоторые дифференцированные элементы (например, проводящие), а во-вторых, в них нет инициальных клеток. Поэтому вставочные меристемы имеют временный характер и, в конце концов, превращаются в постоянные ткани.
Вставочные меристемы хорошо выражены у основания молодых листьев, в которых дифференциация тканей идет от верхушки к основанию органа.
Раневые (травматические) меристемы, как показывает название, возникают при залечивании поврежденных тканей и органов. Раневая меристема около пораненного места чаще всего возникает путем дедифференциации живых клеток с последующим образованием защитной пробки или других тканей.
Рост клеток меристемы. Клетки, возникшие в меристеме, делятся несколько раз, дифференцируются в клетки тех или иных постоянных тканей и оттесняются вновь образующимися клетками. Таким образом, каждая клетка в меристеме имеет ограниченный срок пребывания в составе меристемы и испытывает ряд превращений. Пока ее оболочка тонка и податлива к растяжению, она увеличивает свой объем и принимает размеры и форму, характерные для ее окончательного состояния. Часто оболочка растягивается очень неравномерно, и тогда возникает клетка, сильно вытянутая в одном направлении или снабженная выступами и ответвлениями (рис. 16). Такое неравномерное растяжение объясняется активным влиянием живых протопластов на растяжимость различных участков оболочки. Оболочки соседних клеток обычно растягиваются согласованно и не скользят (не сдвигаются относительно друг друга). Такой согласованный рост обеспечивает сохранность плазматических связей (плазмодесм) между ними.
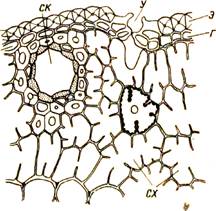
|
Рис 16. Часть поперечного разреза через хвою сосны:
Г — гиподерма; СК — полость смоляного канала,
окруженная клетками эпителия и толстостенными
волокнами; СХ — складчатая хлоренхима (в одной
клетке показаны хлоропласты и ядро); У — устьице;
Э - эпидерма.
АССИМИЛЯЦИОННЫЕ ТКАНИ
Строение и функции. Главная функция ассимиляционных тканей — фотосинтез. Именно в этих тканях синтезируется основная масса органических веществ и связывается энергия, получаемая Землей от Солнца. Процесс фотосинтеза определяет характер всей биосферы нашей планеты и делает ее пригодной для жизни человека.
Ассимиляционные ткани имеют относительно простое строение и состоят из довольно однородных тонкостенных паренхимных клеток. В их постенном слое цитоплазмы содержатся многочисленные хлоропласты. Эти ткани называют также хлорофиллоносной паренхимой или хлоренхимой.
Хлоропласты обычно располагаются одним слоем вдоль стенок клетки. Такое расположение имеет определенный приспособительный смысл: в клетке умещается большое число хлоропластов, которые в наименьшей мере затеняют друг друга и приближены к источнику СО2, поступающего извне. В зависимости от условий освещения и газообмена хлоропласты легко перемещаются (что хорошо видно в листьях элодеи).
В некоторых случаях увеличение поверхности постенного слоя цитоплазмы, а следовательно, и числа хлоропластов в клетке достигается тем, что оболочка образует складки, вдающиеся внутрь клетки, как у хвоинок сосны (рис 16). мощью электронного микроскопа и математические расчеты, в растущей клетке хлоренхимы число хлоропластов быстро увеличивается в 5 и более раз; увеличивается также число рибосом и тилакоидов в них. Общий объем хлоропластов может достигать 70—80% всего объема клеточного протопласта. После того как фотосинтез достиг максимума, во взрослой клетке наблюдаются изменения обратного характера, определяющие старение. Однако если в растущих клетках хлоропласты формируются у всех растений в течение 5—10 дней, то продолжительность их существования и скорость старения могут колебаться от немногих недель (у трав, листопадных деревьев) до нескольких лет (например, у вечнозеленых растений).Расположение в теле растения. Ассимиляционные ткани в теле растения чаще всего залегают непосредственно под прозрачной кожицей (эпидермой), что обеспечивает газообмен и хорошее освещение.
В хлоренхиме находятся большие межклетники, облегчающие циркуляцию газов. Просвечивая сквозь прозрачную эпидерму, хлоренхима придает зеленый цвет листьям и молодым стеблям.
Иногда хлоренхима находится в глубине стебля, под механической тканью или еще глубже, вокруг проводящих пучков. В последнем случае, вероятно, главное значение имеет не синтез углеводов, а выделение кислорода в процессе дыхания. Этот кислород потребляется в процессе дыхания внутренними тканями стебля, в первую очередь живыми клетками проводящих пучков, дыхание которых необходимо для интенсивной деятельности, связанной с проведением веществ.
Хлоренхима имеется также в органах цветка, в плодах. В редких случаях она образуется и в корнях, доступных свету (в воздушных корнях, в корнях водных растений)
ЗАПАСАЮЩИЕ ТКАНИ
Строение и функции. Вещества, синтезированные растением или воспринятые извне, могут откладываться в виде запасов. К накоплению запасных веществ способны все живые клетки. О запасающих тканях говорят в тех случаях, когда запасающая функция выступает на первое место.
Запасающие ткани широко распространены у многих растений и в самых различных органах. Запасаются они в семенах и служат для будущего развития зародыша.
У однолетних растений, проходящих весь жизненный цикл за один сезон,
обычно не бывает значительных отложений веществ в вегетативных органах. Многолетние растения накапливают запасы веществ как в обычных корнях и побегах, так и в специализированных органах — клубнях, корневищах, луковицах, расходуя эти запасы после периодов покоя.
| Рис. 17. Толстостенные клетки из семени финиковой пальмы.
|
Запасающие ткани состоят из живых, чаще всего паренхимных клеток.
Типы запасных веществ. Вещества накапливаются в твердом или растворенном виде. В виде твердых зерен откладываются крахмал и запасные белки. У некоторых растений запасным веществом могут служить гемицеллюлозы, входящие в состав оболочек. Например, много гемицеллюлоз содержат толстые клеточные оболочки в семенах финиковой пальмы (рис. 17). При прорастании семени гемицеллюлозы под действием ферментов превращаются в сахара, мобилизуемые проростком.
В растворенном виде накапливаются сахара, например в корнеплодах свеклы, моркови, в луковицах лука, в стеблях сахарного тростника, в мякоти плодов винограда, арбуза и т. д.
Растения, периодически испытывающие недостаток воды, иногда образуют особые водоносные запасающие ткани. Чаще всего эти ткани состоят из крупных тонкостенных паренхимных клеток, которые содержат слизи, помогающие удерживать воду (рис.18).

Рис. 18. Поперечный разрез листа сведы (Suaeda arcuata):
ВТ — водоносная ткань; ОК — обкладочные клетки; Хл —хлоренхима; Э -эпидерма.
.
АЭРЕНХИМА
Функции межклетников. Во всех органах и почти во всех тканях имеются межклетники, образующие связные системы. Несмотря на то что системы межклетников сообщаются с внешней атмосферой через многочисленные проходные отверстия в покровных тканях, газовый состав в межклетниках сильно отличается от газового состава атмосферы, так как клетки в процессе своей жизнедеятельности (фотосинтеза, дыхания, испарения) выделяют в межклетники одни газы и поглощают другие. Условия обитания и общая организация того или иного растения определяют характер циркуляции газов по межклетникам, необходимый для нормальной жизни.
Довольно часто в растениях образуется ткань с очень большими межклетниками. Если у такой ткани функция вентиляции выступает на первое место, то ее называют аэренхимой.
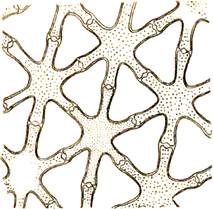
Рис. 19. Аэренхима в стебле ситника (Juncus effusus).
Строение аэренхимы. Чаще всего она представляет собой своеобразную модификацию паренхимы. Однако клетки в ней могут иметь самую различную форму. Особенно сильного развития аэренхима достигает у растений, которые обитают в среде, затрудняющей нормальный газообмен и снабжение внутренних тканей кислородом, например у растений, погруженных в воду или растущих на болотной почве. Прямыми экспериментами было показано, что кислород из надземных органов поступает в корневища и корни по межклетникам.
ВСАСЫВАЮЩИЕ ТКАНИ
Всасывающие ткани играют важную роль в жизни растений. Через них в тело растения из внешней среды поступают вода и растворенные в ней вещества. Они различны по структуре и по распространенности среди высших растений. Наибольшее значение имеет ризодерма (греч. риза — корень; дерма — кожа) — наружный слой клеток на всех молодых корнях. Через ризодерму в корень всасывается из почвы вода и поглощаются растворенные в ней вещества.
Остальные типы всасывающих тканей встречаются или в каких-то определенных систематических группах, или их наличие связано с приспособлением к особым условиям существования, например: воздушные корни орхидей (эпифиты не паразитируют на растении-хозяине, а используют их как подпорки, субстрат для подъема к свету). Воздушные корни приспособлены для поглощения влаги из воздуха, дождя или росы. На поверхности воздушных корней образуется веламен (лат. velamen-покров). Он соответствует ризодерме и всасывает влагу капиллярным путем.
Гаустории своеобразные присоски, внедряющиеся в организм растения-хозяина и соединяющегося с его проводящей системой характерны для растений - паразитов.
Гидропоты состоят из одной или из группы клеток, чаще всего на поверхности листьев у растений, погруженных в воду, и способны избирательно поглощать растворенные в воде вещества. Их можно видеть на нижней стороне плавающих листьев кубышки).
ПОКРОВНЫЕ ТКАНИ
Главное назначение покровных тканей — предохранять растения от высыхания, а также от других неблагоприятных воздействий внешней среды. В зависимости от времени и места возникновения различают три группы покровных тканей: 1) эпидерму, или кожицу, 2) перидерму, 3) корку.
Эпидерма. По происхождению эпидерма — первичная покровная ткань, поскольку она развивается непосредственно из первичной меристемы — верхнего слоя верхушечной меристемы стебля. Она покрывает зеленые органы растения. Чаще всего эпидерма состоит из одного слоя живых, плотно сомкнутых клеток, не содержащих хлоропластов. Стенки клеток обычно извилистые, благодаря чему достигается прочное соединение их между собой. Характерная особенность клеток эпидермы — неодинаковая толщина стенок. Стенки, которые граничат с внешней средой, более толстые, чем остальные. Кроме того, они бывают покрыты сплошным слоем кутикулы или воскового налета. Кутикула представляет собой бесцветную пленку бесструктурного вещества непроницаемую ни для воды, ни для газов. Иногда слой кутина достигает значительной толщины. Восковой налет, так же как и кутин, выделяется клетками эпидермы (рис. 20). У некоторых растений толщина воскового налета доходит до 5 мм, например у пальмы — Klopstockia cerifera. Защитные функции эпидермы усиливаются всевозможными выростами ее

|
Рис. 20. Узел стебля сахарного тростника (поперечный срез):
1 — эпидерма, 2 —воск в виде тончайших палочек
клеток — волосками (трихомами). Волоски исключительно разнообразны по своему строению (рис. 21). Кроме защитной функции, они иногда выполняют выделительную (железистые волоски).

Рис. 21. Волоски на эпидерме: А — картофеля Б — яблони; В — коровяка; Г — лоха; Д — железистые волоски табака; Е —последовательные стадии развития жгучего волоска крапивы (7— 3).
Процессы обмена веществ осуществляются с помощью устьиц. Устьице- это сквозное отверстие, ограниченное двумя клетками-замыкателями полулунной формы. Стенка замыкающих клеток утолщена неравномерно, участок стенки, примыкающей к устьичной щели, значительно толще (рис 22). Замыкающие клетки содержат хлоропласты. Как только солнечный свет падает на зеленую поверхность растения в клетках замыкателях начинается процесс фотосинтеза в результате чего образуется сахар.
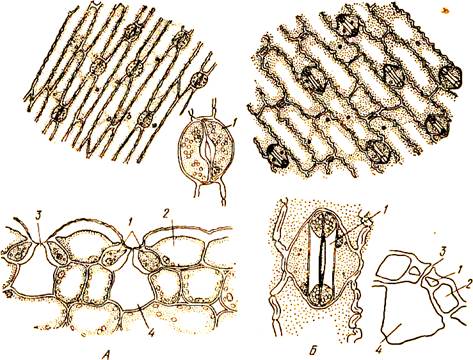
Рис. 22. Эпидерма: А — листа ириса (вид с поверхности и на поперечном срезе, отдельно устьице); Б — листа кукурузы (вид с поверхности, отдельно устьице с поверхности и на поперечном срезе): I — замыкающие клетки. 2 — побочные клетки, 3 — устьичная щель, 4 —воздухоносная полость
. Концентрация клеточного сока замыкающих клеток резко возрастает по сравнению с концентрацией клеточного сока близлежащих клеток эпидермы. В силу осмоса вода из них переходит в замыкающие клетки. При этом тонкая часть их стенки растягивается, утолщенные же части растягиваться не могут. Это приводит к увеличению кривизны клеток и образованию щели между ними. С прекращением фотосинтеза ночью или в жаркий полдень концентрация в замыкающих клетках и прилегающих клетках эпидермы уравнивается за счет оттока воды из замыкающих клеток, и устьица закрываются. Расположены устьица на обеих сторонах листовой пластинки, но преимущественно на нижней. У плавающих листьев водных растений устьица расположены, естественно, только на верхней стороне.
Перидерма развивается из вторичной меристемы — феллогена, который обычно возникает из одного ряда клеток паренхимы, расположенных непосредственно под эпидермой, а иногда в более глубоких слоях, или из клеток самой эпидермы. Эти клетки начинают делиться параллельно поверхности стебля и отделяют дочерние клетки преимущественно в центробежном направлении (феллема) и гораздо меньше в центростремительном (филлодерма) (рис. 22). Феллема (пробка) состоит из правильных радиальных рядов плотно расположенных клеток, стенки которых опробковели, т. е. пропитались суберином, в результате чего содержимое клеток отмирает. Таким образом возникает комплекс, состоящий из трех слоев: феллемы — пробки, феллогена — пробкового камбия и феллодермы — пробковой паренхимы, именуемый перидермой. 
Рис. 23. Перидерма (А) и чечевичка (Б) ветки бузины (поперечный срез):
1— эпидерма, 2 — кутикула, 3 — феллема (пробка), 4— феллоген (пробковый камбий), 5 — фелло-дерма (пробковая паренхима), 6 — паренхима коры, 7 — выполняющая ткань чечевички
Для газообмена и транспирации в перидерме имеются специальные образования — чечевички. Чечевичка представляет собой совокупность крупных паренхиматических более или менее округлых клеток с большими межклетниками, по которым проходят водяной пар и газы (см. рис. 22). Чечевички чаще всего образуются в тех местах, где были расположены участки эпидермы с устьицами.
У пробкового дуба пробковый камбий работает непрерывно до 35 лет. В этом случае откладывается очень мощный слой пробки, иногда достигающий 25 см толщины и более.
 Рис. 24. Корка дуба черешчатого (поперечный срез):
Рис. 24. Корка дуба черешчатого (поперечный срез):
1 — слои феллемы, 2 — участки отмерших тканей коры
Корка образуется на смену перидерме, поэтому ее иногда называют третичной покровной тканью. В типичных случаях корка встречается у деревьев. Поскольку утолщение ствола и ветвей дерева осуществляется из года в год, перидерма под напором разрастающегося в толщину ствола через два-три года разрывается, а в более глубоких слоях коры закладываются новые участки феллогена, дающие начало новым слоям пробки (рис. 24). Эти слои пробки и слои мертвых клеток, заключенные между ними, и составляют корку, обладающую более надежными защитными свойствами, чем перидерма.
ВЫДЕЛИТЕЛЬНЫЕ ТКАНИ
Общая характеристика. Эти ткани выделяют вещества (экскреты или секреты), исключающиеся из обмена веществ (метаболизма).
Растения выделяют вещества, очень разнообразные в химическом отношении, причем разные вещества могут выделяться сходными морфологическими структурами, а одинаковые вещества вырабатываются различными видами выделительных тканей. Остается неясным значение многих выделяемых веществ для самих растений. Вполне определенно можно говорить о функциях лишь немногих выделительных тканей (нектарников, пищеварительных желез).
Вероятно, некоторые вещества представляют собой отбросы, возникающие как побочные продукты в обмене веществ растений. То обстоятельство, что многие из таких веществ выделяются в меристемах, позволяет думать, что их образование сопровождает метаболизм при интенсивных процессах роста. Но также вероятно, что некоторые отбросные вещества в процессе приспособительной эволюции получили какую-то дополнительную, вторичную функцию. Например, некоторые смолы и эфирные масла делают растения несъедобными и отпугивают травоядных животных. Наличие смол в древесине хвойных придает ей большую устойчивость против гниения.
Выделяемые растениями вещества делятся на несколько групп. Наиболее характерную группу составляют терпены (сильно восстановленные производные непредельного углеводорода изопрена). К терпенам принадлежат эфирные масла, бальзамы, смолы и каучук. Терпены образуются чаще всего в смеси друг с другом. Например, живица, добываемая из стволов хвойных деревьев путем подсочки, состоит из целого ряда различных терпенов. Из живицы получают скипидар и канифоль, которые также представляют собой смеси химических соединений.
Растения могут также выделять полисахариды (слизи, сахара), белковые вещества, соли, воду.
Ультраструктура выделительных клеток определенным образом зависит от химической природы выделяемых веществ. Выделение терпенов связано с сильным развитием агранулярного эндоплазматического ретикулума, выделение слизи — с развитием аппарата Гольджи, а выделение белков — с развитием гранулярного ретикулума.
Несмотря на морфологическое и химическое разнообразие, можно указать на несколько типов наиболее часто встречающихся выделительных тканей.
Наружные выделительные ткани. Железистые волоски представляют собой трихомы, т. е. производные эпидермы. У пеларгонии железистый волосок состоит из многоклеточной ножки и одноклеточной головки, которая выделяет эфирные масла в пространство между целлюлозной оболочкой и кутикулой (рис. 54). При разрыве кутикулы экскрет изливается наружу, после чего может образоваться новая кутикула и накопиться новая капля экскрета. Железистые волоски могут быть сидячими, иметь многоклеточную головку и т. д. У мари и лебеды на листьях виден как бы мучнистый налет: он составлен головчатыми трихомами, выделяющими воду и соли (рис. 54, 4).
Иногда наружные железки имеют вид щитка на ножке (например,
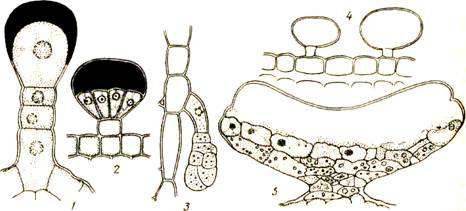
Рис. 54. Железистые волоски и пельтатная (щитовидная) железка:
/— волосок пеларгонии с экскретом, выделенным под кутикулу; 2— волосок розмарина; 3— волосок картофеля; 4-- пузырчатые волоски лебеды с водой и солями в вакуолях; 5 — пельтатная железка с листа черной смородины
пельтатные железкиу смородины, рис. 54, 5).
К наружным железистым тканям можно отнести жгучие волоски крапивы (рис. 55). Они имеют любопытные особенности, позволяющие им действовать наподобие миниатюрных шприцев, впрыскивающих едкие вещества под кожу животного или человека. Большая клетка, наполненная едким соком, находится в состоянии тургора. Вытянутый кончик волоска имеет окремневшие, очень хрупкие стенки, которые при легком прикосновении обламываются и образуют острые края (как у осколка стекла). Волосок впивается в кожу и выдавливает сок.
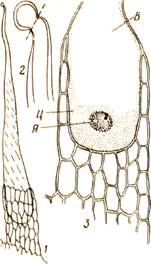 Рис. 55. Жгучий волосок крапивы:
Рис. 55. Жгучий волосок крапивы:
/—общий вид; 2 —окончание волоска
(показана линия облома); 3 —основание
волоска с цитоплазмой (Ц), ядром (Я) и вакуолью (В).
Нектарники выделяют сахаристую жидкость — нектар, служащий приманкой для насекомых-опылителей. Чаще всего они находятся в цветках. Выделительные клетки нектарников имеют густую цитоплазму и высокуюактивность обмена веществ. К нектарнику может подходить проводящий пучок (рис. 56).
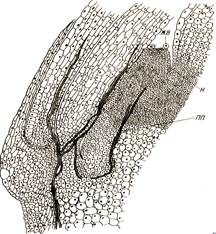
Рис. 56. Нектарник в цветке бархатцев:
ЖВ -- железистые волоски; Я — ткань нектарника; ПП — проводящий пучок.
Гидатоды (греч. гидор, гидатос — вода; одос — путь) выделяют наружу капельно-жидкую воду и растворенные в ней соли. Одну из наиболее сложноорганизованных гидатод можно наблюдать на листьях толстянки (рис. 57). Такая гидатода состоит из

Рис. 57. Гидатода в листе толстянки портулаковой:
1— вид с поверхности; 2 — на поперечном разрезе листа; ВУ — водяные устьица; Г — гиподерма; Об — обкладка; ПП — проводящий пучок; Э — эпидерма; Эп — эпитема.
проводящего пучка, связанного с проводящими тканями листа, мелких бесцветных живых клеток эпитемы, прилегающей к окончанию проводящего пучка, и участка эпидермы над эпитемой. От хлоренхимы листа гидатода отграничена непрерывным слоем бесцветных клеток, образующих обкладку. В эпидерме, лежащей над эпитемой, находятся водяные устьица, отличающиеся от обычных устьиц тем, что лишены подвижности, а щели постоянно открыты. У ряда растений каждая гидатода имеет одно крупное водяное устьице.
При временном избыточном поступлении воды в растение и ослабленной транспирации через гидатоды происходит гуттация, т. е. выдавливание капелек воды. Гуттацию можно наблюдать у многих растений, особенно травянистых (земляники, манжеток), по утрам после прохладных и влажных летних ночей. Так как гидатоды чаще всего расположены по краям листьев, на зубчиках, то как раз на них можно видеть висящие капельки воды.
Значение гуттации, видимо, заключается в том, что растение освобождается от избыточных воды и солей (за минуту до 180 капель воды).
К наружным выделительным тканям относят пищеварительные железки насекомоядных растений (росянки и др.). Жидкость, выделяемая этими железками, содержит вещества (ферменты, кислоты), с помощью которых перевариваются пойманные животные.
Внутренние выделительные ткани. Выделительные клетки рассеяны среди других тканей. Они накапливают различные вещества — оксалат кальция (в виде одиночных кристаллов, друз или рафид), терпены, слизи, танины и пр. Эфир- номасляные вещества характерны для ряда семейств — лавровых, магнолиевых, перечных, кирказоновых и др.
Вместилища для секретов очень разнообразны по форме, величине и происхождению.
Первые возникают из межклетников, заполненных выделенными веществами и окруженных живыми клетками эпителия (рис. 58). Вторые образуются на месте группы клеток, которые распадаются после накопления веществ. Есть промежуточные типы вместилищ.
Смоляные каналы (смоляные ходы) характерны для семейств зонтичных, аралиевых, многих сложноцветных, для хвойных и других. Смоляной канал представляет собой длинный трубчатый межклетник, заполненный смолой и окруженный живыми клетками эпителия. Эпителий выделяет экскрет внутрь полости и изолирует его от живых
 Рис. 58. Схема развития смоляного канала:
Рис. 58. Схема развития смоляного канала:
1—3— на поперечных разрезах; 4— на продольном разрезе; П — полость канала; Э — эпителий.
тканей (рис. 58). Смоляные каналы могут ветвиться, образовывать перемычки (анастомозы), создавая сложно разветвленную систему.
Вместилища могут быть короткими и округлыми и образовывать не смолу, а слизи, эфирные масла (хорошо развиты в кожуре цитрусовых - лимона, апельсина, мандарина).
Млечные трубки (млечники) представляют собой живые клетки, содержащие в вакуолях млечный сок, обычно молочно-белого цвета. У чистотела, однако, млечный сок ярко-оранжевый.
Млечный сок, вытекающий при перерезке млечников, кроме смол и каучука, содержит эфирные масла, белковые соединения, различные алкалоиды. Этот млечный сок, называемый латексом, добывают из стволов таких тропических каучуконосов, как гевея бразильская, культивируемая повсеместно в экваториальной зоне.
Для получения промышленного каучука латекс подвергают сложной технологической обработке.
Млечники бывают двух типов — членистые и нечленистые. Первые возникают из многих отдельных млечных клеток, которые в местах соприкосновения друг с другом растворяют оболочки, и их протопласты и вакуоли сливаются в единую разветвленную систему (рис. 60,1). Членистые млечники найдены у сложноцветных, маковых, колокольчиковых и многих других.
Нечленистый млечник представляет собой одну гигантскую клетку, которая, возникнув в зародыше, более не делится, а непрерывно растет, удлиняется и ветвится. Такими млечниками обладают молочайные, тутовые и другие (рис. 60,2 )
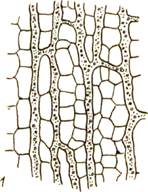
Рис. 60. Млечники:
1— членистый млечник; 2— нечленистый млечник.