АНТИГЕНЫ И ИХ СВОЙСТВА.
АНТИГЕНЫ - это любые генетически чужеродные агенты, вызывающие различные формы иммунного ответа.
Способны:
- вызывать антителообразование и другие формы иммунного ответа;
- взаимодействовать с антителами и рецепторами на Т- и В-лимфоцитах (TCR и BCR).
Это вещества организма человека, микробов, разнообразные химические соединения, которые специфически распознает иммунная система. К антигенам относится огромное число биологически активных макромолекул, как правило, имеющих органическое происхождение (различные биополимеры). Это, в первую очередь, белки (самые сильные антигены) и их комплексы с полисахаридами, нуклеиновыми кислотами растительного, животного, микробного происхождения или искусственно синтезированные вещества. Антигенами также являются бактерии, грибы, простейшие, вирусы, клетки и ткани животных, различные части и органоиды клеток (ЦПМ, рибосомы, митохондрии), микробные яды, экстракты гельминтов, яды многих змей и пчел.
К антигенам не относятся низкомолекулярные вещества органической и неорганической природы, которые являются структурными компонентами клеток и тканей или образуются в норме в результате обмена веществ (аминокислоты, хлорид натрия, этанол и др.).
Экзогенные антигены попадают в организм из окружающей среды (через пищеварительный тракт, дыхательные пути, кровь). Эндогенные антигены образуются внутри организма (аномальные белки, опухолевые клетки, вирусные белки и др.).
Свойства антигенов.
1. Чужеродность - структурное отличие от собственных молекул, в первую очередь белков, из которых состоит организм, которые определяют индивидуальность организма, и информация о структуре которых хранится в генах. Поэтому чужие белки (антигены) распознаются иммунной системой, как чужеродные - не свои. Чужеродность – это есть отличие данного антигена от антигенов организма.
2. Антигенность (иммуногенность) - способность антигена при введении ворганизм индуцировать иммунный ответ, т.е. вызывать образование антител или сенсибилизированных (иммунных) лимфоцитов.
3. Специфичность - способность взаимодействовать только с теми антителами, которые образовались в ответ на этот антиген. Специфичность определяется наличием на поверхности молекулы антигена характерного участка, который связывается с антителом. Этот участок называется – антигенная детерминанта или эпитоп. Он представляет собой определенную химическую группировку (на белке это мозаика поверхностно расположенных аминокислот), которая как "ключ к замку" соответствует определенному участку (активному центру) на молекуле антитела или рецепторам на поверхности Т-и В- лимфоцитов (TCR и BCR). Именно этот участок определяет специфичность взаимодействия антител и лимфоцитов с антигенами. Т.о. можно сказать, что антитела связывают и нейтрализуют только те антигены, которые вызвали их образование. Антигены могут иметь несколько эпитопов (поливалентны).
Виды антигенов.
Полные антигены способны вызывать синтез антител и реагировать с ними. Это наиболее сильные антигены. К ним относятся белки и их комплексы с углеводами, липидами и нуклеиновыми кислотами. Они имеют 2 и более эпитопа, т.е. являются 2-х и более валентными.
Неполные антигены или гаптены - самостоятельно не вызывают выработку антител, но приобретают эту способность при присоединении с высокомолекулярными белковыми носителями. К гаптенам относятся вещества с небольшим молекулярным весом; они имеют одну детерминанту: полисахариды, нуклеиновые кислоты, липиды, пептиды.
Полугаптены – имеют более низкий молекулярный вес, это неорганические радикалы (йод, бром, нитрогруппа и др. радикалы), которые так же, как и гаптены, неспособны вызывать образование антител, но приобретают эту способность после соединения с белками. Такие йодированные или бромированные белки вызывают образование антител, специфичных йоду и брому, т.е. тем детерминантам, которые расположены на поверхности полного антигена, а не белку-носителю.
CD-антигены – молекулы на поверхности клеток, по которым их отличают от других клеток (маркируют). Насчитывается около 350 этих антигенов. На поверхности лимфоцитов имеются CD3, CD4, CD8-антигены.
Гетероантигены – одинаковы антигены у представителей разных видов Например, антигены эритроцитов человека с первой группой крови и антигены возбудителя чумы; стрептококки группы А и антигены клубочков почек человека. Антигенная мимикрия.
Аллоантигены (изоантигены) - разные антигены внутри одного вида (антигены эритроцитов человека с разными группами крови). К ним относятся антигены главного комплекса гистосовместимости (MHC I и II классов). Эти антигены участвуют в презентации антигена ( антигенного пептида ) Т-лимфоцитам. MHC I класса представляют внутриклеточные антигены CD8-цитотоксическим Т-лимфоцитам (ЦТЛ). Имеют все клетки организма (кроме эритроцитов, нейронов). Они являются маркерами «своего» для антигенраспознающих клеток, которые уничтожают клетки с измененными молекуламиMHC I класса. MHC II класса представляют внеклеточные антигены CD4 ТH-лимфоцитам (хелперам). Имеют определенные антигенпрезентирующие клетки (АПК), активированные Т-лимфоциты.
Аутоантигены – собственные антигены организма человека: поврежденные или измененные в генетическом отношении компоненты собственных клеток (приобретенные), а также собственные структуры, отделенные от контакта с иммунной системой организма гематоэнцефалическим, гематотестикулярным и др. барьерами – врожденные (например, мозг, хрусталик глаза, щитовидная железа, сперматозоиды).
Суперантигены – антигены микробов, способные без предварительной переработки (презентации) активировать большие группы лимфоцитов (поликлональная активация). Это сопровождается гиперпродукцией различных медиаторов иммунного ответа, что вызывает синдром общей интоксикации организма. [После поликлональной активации наступает дефицит лимфоцитов – иммунодефицит]. Свойства суперантигенов проявляют энтеротоксины стафилококков, токсин синдрома токсического шока, вирус бешенства, микоплазмы, стрептококки, кампилобактер и др.
Существуют также тимусзависимые антигены, которые индуцируют синтез антител с участием Т-клеток, и тимуснезависимые, индуцирующие синтез антител без помощи Т-клеток.
Большинство антигенов относится к тимусзависимым, к тимуснезависимым относятся липополисахариды клеточных стенок грамотрицательных бактерий, синтетические полимеры и др.
АНТИГЕНЫ МИКРООРГАНИЗМОВ.
Микроорганизмы представляют собой сложный комплекс антигенов.
Группоспецифические антигены встречаются у разных видов одного и того же рода или семейства. Видоспецифические антигены - у разных представителей одного вида. Типоспецифи-ческие антигены (или вариантные) - у разных вариантов в пределах одного и того же вида. Последние подразделяют на серологические варианты (серовары).
Среди бактериальных антигенов различают Н-, О-, К- и др.
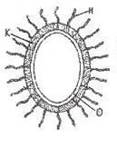
Н – жгутиковые антигены;
К – капсульные антигены;
О – соматический антиген клеточной стенки.
Жгутиковый Н-антиген представлен белком флагеллином жгутиков бактерий. Он разрушается при нагревании, при 5б-80°С, а после обработки фенолом сохраняет свои свойства. Н-антиген получают путем инактивации микробной взвеси формалином.
Соматический О-антиген. Ранее полагали, что О-антиген заключен в содержимое клетки, ее соме, поэтому назвали его соматическим антигеном. Впоследствии оказалось, что этот антиген связан с клеточной стенкой бактерий. О-антиген грамотрицательных бактерий связан с липополисахаридами клеточной стенки. Детерминантными группами этого сложного комплексного антигена являются концевые повторяющиеся звенья полисахаридных цепей (О-специфическая цепь ЛПС), присоединенных к ее основной части. Число и состав сахаров у разных бактерий неодинаковы. Чаще всего в них содержатся гексозы (глюкоза, галактоза, рамноза и др.), аминосахар N-ацетилглюкозамин. У грамотрицательных бактерий О-антиген является их эндотоксином.
О-антиген термостабилен: он сохраняется при кипячении в течение 1-2 часов.
При иммунизации живыми культурами, имеющими жгутики, образуются антитела к О- и Н-антигенами, а при иммунизации кипяченой культурой образуются антитела только к О-антигену.
К-антигены (капсульные) так же, как О-антигены, тесно связаны с липополисахаридами клеточной стенки и с капсулой, но в отличие от О-антигена содержат главным образом кислые полисахариды: глюкуроновую, галактуроновую и другие уроновые кислоты.
По чувствительности к температуре К-антигены подразделяются на А-, В- и L-антигены. А-антигены выдерживают кипячение в течение более 2-х часов. В-антигены не разрушаются при t 60°C в течение часа, L-антигены разрушаются при нагревании до 60°С.
К-антигены располагаются более поверхностно, чем О-антигены, и часто маскируют их. Поэтому для выявления О-антигена нужно кипячением разрушить К-антиген.
Капсульные антигены полисахаридной природы выявлены у пневмококков, клебсиелл и других бактерий, образующих выраженную капсулу. У сибиреязвенных бактерий капсульный антиген состоит из полипептидов.
К капсульным антигенам относится так называемый Vi-антиген брюшнотифозных и некоторых других энтеробактерий, обладающих высокой вирулентностью, в связи с чем данный антиген получил название антигена вирулентности.
Протективный антиген был получен из отечной жидкости сибиреязвенного карбункула. Это термолабильный белок (разрушается при 56°С в течение 30 мин). Он обладает сильно выраженными иммуногенными свойствами, обеспечивающими иммунитет к соответствующему агенту. Протективный антиген образуют возбудители чумы, бруцеллеза, туляремии, коклюша при попадании в организм хозяина, однако он не является их постоянной составной частью.
АНТИТЕЛА И ИХ СВОЙСТВА.
АНТИТЕЛА - это белки глобулиновой фракции крови, которые специфически соединяются с антигенами, вызвавшими их образование.
Они называются иммуноглобулинами и обозначаются Ig. По химическому составу они являются гликопротеидами (белок + углевод) и образуются В-лимфоцитами (плазматическими клетками).
Функции антител: 1) участвуют в реакциях образования комплексов антиген-антитело в результате чего происходит связывание, нейтрализация и выведение антигена из организма;
2) усиливают фагоцитоз вместе с другими опсонинами;
3) активация комплемента;
4) входят в состав рецепторов В-лимфоцитов.
Основные свойства антител:
1) гетерогенность – способность вырабатывать на одну молекулу антигена столько антител, сколько у антигена эпитопов;
2) специфичность - способность антител вступать в реакцию только с тем антигеном, на который выработалось данное антитело.
Структура антител
По химическому составу иммуноглобулины относятся к гликопротеидам, т.к. их молекула состоит из протеина и углеводов.
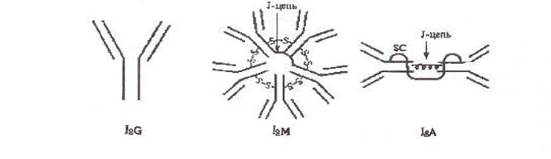
Ig характеризуют по молекулярной массе, константе седиментации и т.п. Различия этих свойств позволили разделить их на 5 классов: IgG, IgM, IgA, IgE и IgD. IgM, G, А имеют подклассы. Все классы и подклассы различаются по аминокислотным последовательностям.
 Структурной единицей антител является мономер, состоящий из четырех полипептидных цепей - двух тяжелых Н-цепей (от англ. heavy - тяжелый) и двух легких L-цепей (от англ. light - легкий), соединенных между собой дисульфидными мостиками -S-S-. Легкие и тяжелые цепи отличаются по молекулярной массе: мол. масса L-цепей 23000, мол. масса Н-цепей 50000-70000. Соответственно каждому классу иммуноглобулинов М, G, A,E, D различают пять типов тяжелых цепей: μ (мю), γ (гамма), α (альфа), ε (эпсилон), δ (дельта). Легкие цепи всех пяти классов являются общими и бывают двух типов: κ (каппа) и λ (лямбда). В одной и той же молекуле могут быть только идентичные L-цепи.
Структурной единицей антител является мономер, состоящий из четырех полипептидных цепей - двух тяжелых Н-цепей (от англ. heavy - тяжелый) и двух легких L-цепей (от англ. light - легкий), соединенных между собой дисульфидными мостиками -S-S-. Легкие и тяжелые цепи отличаются по молекулярной массе: мол. масса L-цепей 23000, мол. масса Н-цепей 50000-70000. Соответственно каждому классу иммуноглобулинов М, G, A,E, D различают пять типов тяжелых цепей: μ (мю), γ (гамма), α (альфа), ε (эпсилон), δ (дельта). Легкие цепи всех пяти классов являются общими и бывают двух типов: κ (каппа) и λ (лямбда). В одной и той же молекуле могут быть только идентичные L-цепи.
Как в Н- так и в L-цепях имеется вариабельная V-часть (от англ. various -разный), в которой последовательность аминокислот непостоянна. Остальная часть Н- и L-цепей называется константной С-частью (от англ. constant - постоянный).
При обработке меркаптоэтанолом разрушаются дисульфидные связи, и молекула иммуноглобулина распадается на отдельные цепи полипептидов.
При воздействии протеолитическим ферментом - папаином Ig распадается на три фрагмента: два некристаллизующихся, содержащих антигенсвязывающие центры (активные центры) и названных Fab-фрагментами I и II (от англ. fragment antigen binding – фрагменты, связывающие антиген) и один Fc-фрагмент кристаллизующийся (от англ. fragment crystalizable). Fab-I и Fab-II фрагменты сходны по свойствам и аминокислотному составу и отличаются от Fc-фрагмента. Fab- и Fc-фрагменты соединены между собой гибкими участками Н-цепи (шарнир), благодаря чему молекулы Ig имеют гибкую структуру.
Активный центр (антигенсвязывающий) образован вариабельными концевыми участками легких и тяжелых цепей (V-частъ) и представляет собой полость, пространственная конфигурация которой в точности повторяет пространственную конфигурацию антигенной детерминанты (как перчатка повторяет форму руки). Активный центр занимает около 2% поверхности молекулы Ig.
Соединение антигена с антителом обеспечивают ван-дер-ваальсовы силы, гидрофобные и электростатические взаимодействия, водородные связи.
Таким образом, каждый активный центр формируется двумя цепями - легкой и тяжелой, т.е. полноценной активностью антител ни одна из цепей в отдельности не обладает.
Валентность антитела определяется числом его активных центров.
Полные антитела имеют не менее двух активных центров. Они могут связать две молекулы антигена, образуя сетевую структуру (конгломераты), которая выпадает в осадок (видимые изменения).
Неполные антитела имеют один функциональный активный центр, они не образуют сетевой структуры иммунных комплексов (АГ+АТ), т.е. осадка не образуют.
Моноклональные антитела распознают только одну антигенную детерминанту.
Поликлональны е антитела (например, сыворотки) распознают несколько антигенных детерминант в составе антигена.
КЛАССЫ ИММУНОГЛОБУЛИНОВ.
Иммуноглобулины класса М состоят из пяти мономеров соединенных полипептидной цепью (J-цепь) и являются десятивалентными, обладают наиболее высокой авидностью. Это наиболее ранний класс антител, обнаруживаемый при первичном попадании антигена (антитела других классов синтезируются позднее). Обладают свойствами агглютининов, преципитинов, лизинов, опсонинов. Активируют комплемент. Наличие IgM антигенов конкретного возбудителя указывает на наличие острого инфекционного процесса.
Иммуноглобулины класса G являются основным классом антител (до 75% всего Ig), двухвалентны. При первичном контакте с антигеном синтезируются после IgM, а при вторичном - образуются первыми. Единственные из иммуноглобулинов, проникающие через плаценту, обеспечивая пассивный иммунитет плода. Обладают свойствами агглютининов, преципитинов, лизинов, опсонинов. Активируют комплемент.
Обнаружение высоких титров IgG к антигенам конкретного возбудителя указывает на то, что организм находится на стадии реконвалесценции или конкретное заболевание перенесено недавно.
Иммуноглобулины класса А существуют в нескольких формах: моно-, ди- и тримерной. Мономерная форма встречается в сыворотке крови (10-15% всех иммуноглобулинов). Иммуноглобулин А синтезируется плазматическими клетками слизистых оболочек и подслизистых тканей дыхательных путей и кишечного тракта. Часть попадает в кровь, но большая часть остается в слизистых оболочках и играет существенную роль в местном иммунитете, поскольку препятствует адгезии патогенных микроорганизмов на эпителиальных клетках слизистых оболочек.
Секреторный IgA отличается от сывороточного наличием секреторного компонента (SC), выделяемого эпителиальными клетками, что защищает IgA от разрушения ферментами. В отличие от сывороточного секреторный IgA активирует комплемент, что ведет к стимуляции местной фагоцитарной защиты.
Иммуноглобулины класса Е или реагины участвуют в аллергических реакциях. IgE обладают выраженной цитофильностъю, т.е. способностью присоединяться к базофилам и тучным клеткам, в результате чего эти клетки выделяют гистамин и гистаминоподобные вещества, являющиеся медиаторами аллергической реакции. Комплемент не активируют.
Иммуноглобулины класса D обнаруживаются на поверхности развивающихся В-лимфоцитов. Их биологическая роль не установлена. Некоторое увеличение титров IgD находят при беременности, у больных бронхиальной астмой, системной красной волчанкой и лиц с иммунодефицитами. Комплемент не активируют.






