Нуклеокапсид вируса имеет форму икосаидра диаметром 50нм с липидной оболочкой, включающей белковые структуры – выступы высотой 6-8 нм (рис. 1.1) [15]. Применение поли- и моноклональных антител к Е1-протеину вируса гепатита С позволило S. Wotanabe с соавторами локализовать эти белки на оболочке вируса. Они чаще ассоциированы с липопротеинами низкой плотности или находятся в составе иммунных комплексов. От чего зависит величина плавучей плотности частицы вируса [14].
Нуклеокапсид вируса содержит однонитевую линейную молекулу РНК позитивной полярности длиной 9,4-9,5 тысяч нуклеотидов (рис. 1.2). В геноме выделяют две зоны: кодирующая структурные белки расположена у 5'-области РНК, кодирующая неструктурные (функциональные) – у 3'-области.
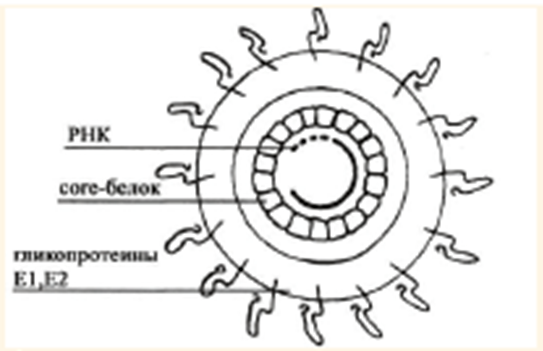
Рисунок 1.3 – Строение вириона вируса гепатита С
Одна открытая рамка считывания кодирует полипротеин длиной 3008-3037 а.о., который процессируется комбинацией вирусных протеиназ и протеиназ клетки-хозяина. На 5'- и 3'-концах генома вируса находится не транслируемый регион (5'- и 3'-NTR) [12, 14, 15].
Протяженность 5'-NTR РНК ВГС составляет около 340 нуклеотидов. Функция этого участка заключается в инициации трансляции. Специфически связываясь с рибосомами (благодаря наличию внутреннего рибосомального сайта связывания) и факторами трансляции клетки-хозяина, он направляет рибосому к инициирующему кодону (AUG) в позиции 342, после чего начинается синтез полипротеина [15, 21].
В 3'-NTR-части РНК вируса гепатита С выделяют три отдела: первый – протяженностью в 28-42 нуклеотида, второй – содержащий поли-U-/поли-А-тракт, и третий – терминальный, высококонсервативный участок протяженностью в 98 нуклеотидов [15]. По аналогии с другими флавивирусами можно предположить, что этот участок играет определенную роль в репликации вируса и упаковке нуклеиновой кислоты в вирусную частицу. Наличие в 3'-NTR-регионе РНК вируса гепатита С различных изолятов высококонсервативного участка нуклеиновых кислот определяет интерес к этому участку в связи с перспективой разработки эффективных лекарственных препаратов [14, 21].
Участок РНК вируса гепатита С, примыкающий к 3'-концу 5'-NTR- региона, протяженностью в 573 нуклеотида, кодирует белок, формирующий капсид вируса (С, или Core) (рис. 1.3). Его С-терминальный регион является необходимым для правильной укладки полипептидной цепи [22].
Выявлено три формы С-белка. Полноразмерная (р21) с молекулярной массой 21 кДа и усеченная (р19), которые обнаружены в мембране эндоплазматического ретикулума. Форма (р16) С-белка, обнаруженная в ядрышках инфицированных гепатоцитов [14, 21]. Он может оказывать влияние на человеческие онкогены, задействованные в развитии гепатоклеточной карциномы у пациентов с хроническим гепатитом С путем супрессии отдельных генов клетки-хозяина. Считается, что С-белки р16 и р21 разнонаправленно влияют на транскрипционную активность опухолевого репрессора р53: первый блокирует, а второй усиливает ее. Таким образом, специфически подавляется апоптоз инфицированных клеток [15]. Кроме того, капсидный белок играет основную роль в индукции митохондриальной дисфункции [23].
На своей поверхности С-белок несет различные высоко-консервативные В-клеточные эпитопы, существование которых чрезвычайно важно для выявления анти-ВГС в процессе лабораторной диагностики инфекции. Core-белок обладает способностью связываться с вирусной РНК, а также выполняет регуляторную роль в репликации вируса [14, 21, 24].
В Е1- и Е2-регионах РНК вируса гепатита С регистрируется значительная генетическая изменчивость и содержится информация о белках оболочки вируса, участвующих в проникновении вируса в клетку, в развитии иммунитета и ускользании от иммунного ответа организма за счет отбора мутантов, способных уклоняться от действия нейтрализующих антител [14, 15]. Кроме того, на мышиной модели показано, что участок Е1-Е2 совместно с Сore ускоряет опухолеобразование, блокируя способность клеток к апоптозу [25].
Зоны генома Е1 и Е2 кодируют белки с молекулярной массой 31 и 70 кДа соответственно. Имеются две формы белка Е2: gp70A (E2) и gp70B (E2-p7). Последний в структуре вируса не обнаружен, предполагается его участие в высвобождении вновь синтезированного вириона вируса из инфицированной клетки [14, 15]. Белки Е1 и Е2 сильно гликозилированы и содержат С-терминальный гидрофобный домен, функционирующий как мембранный якорь. Их созревание происходит при удалении С-концевой последовательности сигнальными пептидазами клетки [24]. Установлено, что при отсутствии белка Е2 происходит нарушение пространственной структуры оболочечного белка Е1 [14].
В неструктурной зоне, расположенной ближе к 3'-концу РНК вируса гепатита С, выделяют участки NS2, NS3, NS4A, NS4B, NS5A и NS5B. В отличие от других флавивирусов, в РНК вируса гепатита С отсутствует первая зона, кодирующая неструктурные белки (NS1), а к Е2-региону примыкает зона NS2 (рис. 1.4). Большинство белков, кодированных неструктурными зонами РНК вируса гепатита С, необходимо для репликации вируса [14, 15]. Антитела, вырабатываемые на неструктурные белки, не обладают полноценными протективными свойствами в отношении вируса гепатита С вследствие его высокой генетической вариабельности [24, 26, 27]. NS2 протеин является трансмембранным белком с молекулярной массой 23 кДа [14]. Его С-конец смотрит в просвет цистерн эндоплазматического ретикулума, N-конец – в цитозоль. Этот белок является цинк-зависимой аутопротеазой, разрезающей NS2 и NS3 белки [15].

Рисунок 1.4 – Организация генома вируса гепатита С
NS3 протеин, с молекулярной массой 70 кДа, выполняет несколько различных функций: является сериновой протеазой (амино-терминальная часть), отщепляя от полипротеина все остальные неструктурные белки (NS3/NS4A, NS4A/NS4B, NS4B/NS5A и NS5A/NS5B); обладает хеликазной и нуклеотидтрифосфатазной активностью [14, 15].
При репликации вируса NS3 белок связывается с поли-U последовательностью на 3'-конце вирусного генома своим РНК-связывающим доменом и затем происходит раскручивание двунитевой РНК. Одновременно идет гидролиз дезоксирубонуклеотидтрифосфатов, осуществляемый другим доменом NS3 [15].
P. Brunetal. (2010) в своей работе показал, что NS3 протеаза может ингибировать сигнальные пути трансдукции врожденного иммунитета. Белок способен специфически взаимодействовать с каталитической субъединицей клеточной протеинкиназы А, участвующей в передаче клеточных сигналов, влияя на переход клетки в состояние неконтролируемого роста. C другой стороны, взаимодействие NS3 с иммунной системой может влиять на течение заболевания либо со спонтанным очищением от вируса, либо развитием хронической инфекции [15, 28].
В NS4 регионе выделяют две зоны – NS4A и NS4B, которые кодируют два гидрофобных белка с молекулярными массами 8 и 26 кДа [14, 15]. NS4A протеин является кофактором для NS3 протеазы, образуя с NS3 белком единый комплекс и выполняя функцию якоря, удерживающего на мембране ядра клетки репликативный комплекс вируса гепатита С [14]. Он также необходим для гиперфосфорилирования NS5A протеина. Функция NS4B протеина остается неясной, предполагается, что он также принимает участие в формировании репликативного комплекса [15]. NS5 регион полипротеина построен из двух больших белков — NS5A (56 кДа) и NS5B (65 кДа). Они освобождаются из полипротеина с помощью NS3-NS4A протеазного комплекса.
NS5A белок высококонсервативен, на ядерной мембране инфицированных клеток он совместно с NS5B образует мембранно-связанный репликативный комплекс. NS5A протеин участвует в процессах фосфорилирования и играет важную роль в формировании механизмов устойчивости клеток к действию интерферона. Предположительно белок NS5A ингибирует один или несколько клеточных белков (например, взаимодействует с интерферон-индуцированной клеточной протеинкиназой), участвующих в начальных стадиях противовирусного действия интерферона [12, 14].
NS5B протеин консервативен и является РНК-зависимой РНК-полимеразой с молекулярной массой 68 кДа, обеспечивая репликацию/ транскрипцию генома вируса [15]. Изучение функциональной роли вирусных белков, кодированных неструктурной зоной РНК вируса гепатита С, прежде всего, важно для поиска новых лечебных препаратов, которые могли бы эффективно блокировать репликативную активность вируса.





