Это система взаимоотношений между членами одной популяции. Формы совместного существования особей в популяции могут быть различны. Они зависят от того, одиночный или групповой образ жизни свойствен виду.
Одиночный образ жизни. Полностью одиночное существование организмов в природе не встречается, так как при этом было бы невозможным осуществление их основной жизненной функции – размножения. Однако на определённых стадиях жизненного цикла особи популяций многих видов обособлены и независимы друг от друга. Подобный образ жизни ведут, например, некоторые водные обитатели с наружным способом оплодотворения (актинии), насекомые (божьи коровки, жуки-жужелицы и др.). Нередко виды с одиночным образом жизни образуют временные скопления особей в период, предшествующий размножению, в местах зимовок и т. д. Так, божьи коровки осенью собираются группами в сухой подстилке возле комлей деревьев, щуки и сомы – в зимовальных ямах на дне водоёма. Однако подобные скопления не сопровождаются установлением тесных связей между особями.
Дальнейшее усложнение отношений внутри популяции происходит по двум направлениям: усилению связи между половыми партнёрами и возникновению контактов между родительскими и дочерними поколениями. В популяциях на этой основе формируются семьи, которые очень разнообразны по составу и длительности существования.
Семейный образ жизни усиливает связи между родителями и их потомством. Известный вид такой связи – забота одного из родителей об отложенных яйцах: охрана кладки, насиживание и др. При этом забота о птенцах продолжается до вылета их из гнезда, а у некоторых крупных млекопитающих (тигры, медведи) детёныши воспитываются в семье в течение нескольких лет до наступления их половой зрелости. В зависимости от того, кто из родителей берёт на себя уход за потомством, различают семьи отцовского, материнского и смешанного типов. В семьях с устойчивым образованием пар в охране и выкармливании детёнышей принимают участие оба родителя.
При семейном образе жизни территориальное поведение животных выражено наиболее ярко: различные сигналы, маркировка, ритуальные формы поведения и прямая агрессия обеспечивают владение участком, достаточным для выкармливания потомства.
Дальнейшее усложнение поведенческих связей в популяциях приводит к формированию более крупных объединений животных – колоний, стай и стад.
Колонии – групповые поселения оседлых животных, которые могут существовать длительное время или создаваться на период размножения как у птиц (грачи, чайки, гагары и др.). По сложности взаимосвязей между особями колонии животных очень разнообразны – от простых территориальных скоплений одиночных форм до объединений, в которых отдельные члены выполняют разные функции в жизни популяции. Примером элементарной колонии являются поселения морских жёлудей, которые возникают на основе хемотаксиса, свойственного личинкам. Взаимоотношения между особями в такой колонии сводятся к стимуляции синхронного полового созревания членов колонии.
Более сложной формой колонии являются поселения животных, в которых некоторые функции их жизни выполняются сообща, что повышает шансы на выживание отдельных особей. Такими общими функциями чаще всего становятся защита от врагов и предупредительная сигнализация. Птицы сообща изгоняют даже крупных хищников, с которыми они не справились бы поодиночке.
Колонии млекопитающих (сурков, пищух, пеструшек и др.) обычно возникают на основе разрастания семейных групп с сохранением связей между отпочковывающимися семьями.
Сложные колонии формируются на основе сильно разрастающейся семьи общественных насекомых – пчёл, термитов, муравьёв. В них насекомые сообща выполняют большинство основных функций: размножения, защиты, обеспечения кормом себя и потомства, строительства и др. При этом существуют обязательное разделение труда и специализация отдельных особей, возрастных групп на выполнение определённых операций. Члены колонии действуют на основе постоянного обмена информацией друг с другом. По мере усложнения колонии поведение, а нередко строение и физиология отдельной особи всё больше подчиняются интересам всей колонии.
Стаи – временные объединения животных, которые облегчают выполнение каких-либо функций в жизни вида: добычу пищи, защиту от врагов, миграцию. Стайность наиболее широко распространена среди рыб и птиц, у млекопитающих характерна для многих собачьих. В стаях сильно выражены подражательные реакции и ориентация на соседей.
По способам координации действий стаи делятся на два типа: 1) без выраженного лидера; 2) стаи с лидерами, на которых ориентируются остальные особи. Объединения первого типа характерны в основном для рыб, но встречаются также у перелётной саранчи, мелких птиц и некоторых других видов. Стаи с лидером характерны для крупных птиц и млекопитающих. В процессе групповой охоты хищников, например, волков, лидер организует засады, захват лидера и другие действия, которые требуют согласованности действий всех членов стаи.
Стада – более постоянные объединения животных по сравнению со стаями. Здесь осуществляются все основные функции жизни вида. В основе группового поведения животных в стадах лежат взаимоотношения доминирования – подчинения, основанные на индивидуальных различиях между особями. Одним из вариантов организации стад являются группы с временными или относительно постоянными лидерами, которые определяют направление перемещения, места кормления, реакцию на хищников и т. д. Лидером становится более опытный член стада, которое действует как единое целое, подражая лидеру. Деятельность лидера не направлена непосредственно на подчинение других особей.
Наибольшей сложностью отличается организация стад с вожаками и иерархическим соподчинением особей. В отличие от лидеров поведение вожаков направлено на активное управление стадом: специальными сигналами, угрозами и прямой агрессией. Ранг животного в группе определяется многими факторами: возрастом, физической силой, опытом и др. Как правило, над слабыми доминируют сильные и опытные с устойчивым типом нервной системы. Животные низшего ранга ведут себя покорно перед всеми остальными, подходят к пище и водопою в последнюю очередь. Они изгоняются с лучших мест отдыха, не подпускаются к самкам и т. п.
Иерархически организованному стаду свойственны закономерный порядок перемещения, определённая организация при защите, расположение на местах отдыха и т.д. Например, при передвижении стад павианов в центре – в наибольшей безопасности – находятся самки с детёнышами или беременные, по краям вожаки, молодые самцы и неразмножающиеся самки. Впереди и позади стада располагаются крупные самцы, готовые отразить нападение хищников.
Биологический смысл иерархической системы организации стада заключается в создании согласованного поведения группы, выгодного для всех её членов. После «расстановки сил» животные не тратят лишней энергии на индивидуальные конфликты, в целом группа получает преимущества, подчиняясь наиболее сильным и опытным особям.
Эффект группы – это оптимизация физиологических процессов, ведущих к повышению жизнеспособности особей при их совместном существовании. Проявляется эффект группы как реакция отдельных особей на присутствие других членов своего вида. Например, у овец вне стада резко учащается пульс и дыхание, а при виде приближающегося стада эти процессы нормализуются. Эффект группы проявляется также в ускорении темпов роста животных, повышении плодовитости, увеличении средней продолжительности жизни индивидуумов и др. Вне группы многие животные перестают размножаться. Так, голуби некоторых пород не откладывают яйца, если не видят других птиц. Эффект группы не проявляется у видов, ведущих одиночный образ жизни. Если таких животных заставить жить вместе, у них нарушается нормальный ход многих физиологических процессов, повышается раздражительность.
Положительный эффект группы проявляется до некоторого оптимального уровня плотности популяции. Если животных становится слишком много, это грозит всем недостатком ресурсов среды. Тогда вступают в действие механизмы, приводящие к снижению численности особей в группе.
Динамика популяций
Репродуктивный потенциал и рост популяции. Динамика популяции – это процессы изменений её основных биологических показателей во времени. Особенно большое значение имеет изменение её численности. Любая популяция теоретически способна к неограниченному росту численности, если её не лимитируют факторы среды. В таком случае скорость роста численности популяции будет зависеть только от величины репродуктивного (биотического) потенциала, свойственного виду. Репродуктивный потенциал отражает максимальное число потомков от одной пары (или одной особи) за единицу времени (сутки, год или за весь жизненный цикл). Величина репродуктивного потенциала неодинакова у разных видов и зависит от размеров особей, продолжительности их жизни, уровня эволюционного развития и смертности в популяциях. Корреляция видового уровня плодовитости и средней нормы гибели установлена для различных групп живых организмов. Растения, грибы производят огромное количество спор, так как большинство из них погибает, не попав в благоприятные для развития условия. Рыбы с пелагической икрой, доступной для поедания многими животными, откладывают её в огромном количестве, тогда как виды, укрывающие каким-либо способом свою икру, имеют более низкую плодовитость. Плодовитость сильно зависит от степени развития заботы о потомстве или обеспеченности яиц питательными веществами. Например, акулы, яйца у которых крупные, с большим запасом питательных веществ и имеют плотную защитную оболочку, откладывают несколько яиц за сезон. Зелёная жаба, не заботящаяся о потомстве, производит за сезон 8–12 тысяч икринок, тогда как жаба-повитуха, наматывающая икру на задние лапы, – всего 150 штук.
Таким образом, появление в эволюции каких-либо форм заботы о потомстве чётко коррелирует со снижением видовой нормы плодовитости. У таких видов снижается смертность в раннем возрасте и сохранение высокого репродуктивного потенциала биологически не выгодно. Обратная зависимость наблюдается между плодовитостью и средней продолжительностью жизни, свойственной данному виду. Долгоживущие виды отличаются меньшей плодовитостью. В реальной обстановке конкретные величины как плодовитости так и смертности постоянно колеблются, соответственно этому меняются и уровень численности популяции, и тенденции её изменения во времени. Если бы всё потомство выживало, то численность любой популяции увеличивалась бы в геометрической прогрессии. Такой рост выражается экспоненциальной кривой (рис. 10).
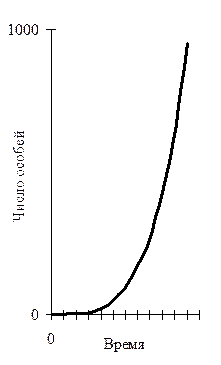 Рис. 10. Теоретическая кривая роста популяции
Рис. 10. Теоретическая кривая роста популяции
В природе биотический потенциал популяции никогда не реализуется полностью. Это может наблюдаться в отдельных случаях в течение короткого периода времени, когда, например, быстро размножающиеся организмы (бактерии, грибы, насекомые) осваивают какой-либо субстрат или среду, где нет конкурентов.
Естественный рост популяции никогда не реализуется в форме экспоненциальной модели, так как он ограничен комплексом факторов внешней среды (наличием ресурсов, конкурентов и др.) и реально складывается как результат соотношений меняющихся значений рождаемости и смертности. Наиболее близко естественный рост численности популяций отражает логистическая модель роста, в которой изменение численности во времени выражается S-образной кривой (рис.11).
 Рис. 11. Логистическая кривая роста популяций
Рис. 11. Логистическая кривая роста популяций
В соответствии с логистической моделью рост численности популяции некоторое время идёт медленно. Затем кривая численности резко возрастает и наконец выходит на уровень, определяемый ёмкостью среды. Этот конечный уровень отражает уравновешенность процессов рождаемости и смертности в соответствии с наличием пищевых и иных ресурсов среды.
Колебания численности популяции. Рассмотренные выше модели роста численности популяции представляют собой «идеальные» схемы, основанные на видовых особенностях механизма самоподдержания популяции. Сложность условий, в которых функционируют естественные популяции, приводит к тому, что их численность после достижения верхнего уровня роста подвержена изменениям. Такие колебания могут возникать либо в результате изменения физической среды, вследствие чего повышается или снижается верхний предел численности, либо в результате внутрипопуляционных и межпопуляционных взаимодействий. После того как верхний предел численности популяции достигнут, она может некоторое время оставаться на этом уровне (рис. 12, кривая 1) или сразу резко упасть (рис. 12, кривая 2). Возможна и такая ситуация, при которой численность популяции «перескакивает» через уровень (кривые 3, 4). В этом случае кормовые и другие ресурсы, необходимые для жизни популяции, накоплены ещё до начала её роста, а механизмы регуляции численности ещё не действуют. Например, при вселении животных туда, где их раньше не было.
 1 2 3 4
1 2 3 4
Рис. 12. Различные типы (1–4) колебания численности популяции
Колебания численности популяции могут быть непериодическими (вызванными воздействием каких-либо факторов) и периодическими (циклическими, повторяющимися через определённый период времени). Непериодические колебания численности носят непредвиденный характер.
Циклические изменения численности с периодом в несколько лет известны для популяций некоторых арктических млекопитающих и птиц. Например, у зайца-беляка и рыси период колебаний численности равен примерно 9–10 лет (рис. 13).

Рис. 13. Периодические колебания популяций зайца-беляка (1) и рыси (2), установленные по числу шкурок, заготовленных Компанией Гудзонова пролива (из Р. Дажо, 1975)
Более короткие, 3- и 4-годичные, циклы характерны для многих обитателей тундры: песца, полярной совы, лемминга, полёвок, лисиц. Подобные колебания численности известны также для многих насекомых (бабочек, саранчи и др.)
 Характер закономерных периодических изменений численности популяций видоспецифичен и связан с особенностями биологии вида, его физиологии и места в естественных экосистемах. С.А. Северцов, ещё в 40-х годах прошлого века проанализировавший многолетний ход динамики численности популяций у многих видов млекопитающих и птиц, установил три фундаментальных типа: 1) стабильный, 2) лабильный, 3) эфемерный (рис. 14).
Характер закономерных периодических изменений численности популяций видоспецифичен и связан с особенностями биологии вида, его физиологии и места в естественных экосистемах. С.А. Северцов, ещё в 40-х годах прошлого века проанализировавший многолетний ход динамики численности популяций у многих видов млекопитающих и птиц, установил три фундаментальных типа: 1) стабильный, 2) лабильный, 3) эфемерный (рис. 14).
Рис. 14. Типы динамики населения по С.А. Северцову:
I – стабильный,
II – лабильный,
III – эфемерный
Стабильный тип характеризуется малой амплитудой и длительным периодом колебаний численности (10–29 лет). Он свойствен крупным животным с большой продолжительностью жизни, поздним наступлением половой зрелости и низкой плодовитостью. Это соответствует низкой норме естественной смертности, так как имеются эффективные механизмы адаптации к неблагоприятным факторам. Примером могут служить копытные млекопитающие, китообразные, приматы, орлы и др.
Лабильный тип динамики отличается закономерными колебаниями численности с периодами от 5 до 11 лет и более значительной амплитудой (численность меняется в десятки раз). Характерен для животных с продолжительностью жизни 10–15 лет, ранним половым созреванием и более высокой плодовитостью (зайцеобразные, крупные грызуны, многие птицы, рыбы и др.).
Эфемерный тип динамики отличается резко неустойчивой численностью: она может возрастать в сотни раз и очень быстро падать. Перепады от максимума до минимума происходят очень быстро, иногда в течение одного сезона. Длина цикла составляет до 4–5 лет, в течение которых «пик» численности занимает не более одного года. Такой тип динамики характерен для видов с большой плодовитостью, высокой нормой гибели и с продолжительностью жизни не более 3 лет (мелкие грызуны, насекомые с коротким циклом развития).
Рассмотренные типы динамики показывают связь этих процессов с биологическими особенностями видов и их популяций, отражают разные жизненные стратегии. Эта мысль лежит в основе концепции экологических стратегий, разработанной американскими экологами Э. Мак-Артуром и Э. Уилсоном (1967). Суть этой концепции сводится к тому, что успешное выживание и воспроизводство вида возможно либо путём совершенствования адаптированности организмов и их конкурентоспособности, либо путём интенсификации размножения, что компенсирует повышенную гибель особей и в критических ситуациях позволяет быстро восстановить численность. Первый путь назван «К-стратегией» – это крупные формы с большой продолжительностью жизни, численность их лимитируется преимущественно внешними факторами. К-стратегия означает «отбор на качество», повышение адаптивности и устойчивости; «r-стратегия» – «отбор на количество» через компенсацию больших потерь высокой плодовитостью. Виды r-стратегии легко осваивают местообитания с нестабильными условиями, благодаря высокой плодовитости быстро восстанавливают любые потери в популяции.
Аналогичные жизненные стратегии свойственны и растениям. Английский учёный Д. Грайм выделил среди них такие стратегии:
конкурентная – достигающая высокой плотности в оптимальных местообитаниях;
стресстолерантная – устойчивые к неблагоприятным факторам, но малопродуктивные виды, заселяющие менее благоприятные места;
рудеральные – отличаются высоким репродуктивным потенциалом и быстрым ростом, заселяют антропогенные места.
Как видно из изложенного, в экологических стратегиях растений и животных обнаруживается большое сходство, что говорит об общности наиболее фундаментальных экологических основ динамики популяций в разных группах живых организмов.
Факторы динамики численности. Факторы, ответственные за изменение численности популяций в современной экологии, принято делить на две группы: 1) факторы, не зависящие от плотности популяции; 2) факторы, зависящие от плотности популяции.
К первой группе относятся абиотические факторы среды. Они действуют на уровне организма, действие их одностороннее, т.е., организмы не могут оказать на них обратное влияние. Их называют модифицирующими факторами, так как они вызывают изменения численности популяций, но не могут её регулировать. Эффект воздействия абиотических факторов на уровень численности реализуется через изменение рождаемости и смертности. Благоприятные условия могут послужить причиной массовой вспышки размножения вида; а в случае стадных саранчовых неблагоприятные климатические погодные условия, наоборот, снижают численность популяции иногда до полного её исчезновения. Например, в малоснежные холодные зимы в лесной зоне из-за промерзания почвы в массе гибнут кроты, полёвки, мыши и др. Заморозки в начале лета могут повлиять на выживаемость многих организмов.
Факторы, зависящие от плотности популяции, не просто изменяют её численность, а сглаживают колебания, приводя после отклонения от оптимума к прежнему уровню. Это происходит потому, что эффект их воздействия тем сильнее, чем выше плотность популяции. Их называют регулирующими факторами, так как они действуют по принципу обратной связи. В качестве регулирующих факторов выступают межвидовые и внутривидовые отношения организмов: возбудители болезней, конкуренция, хищничество, паразитизм, собирательство и др.
Одна из важных форм взаимоотношений организмов, влияющая на периодические изменения численности популяций, – отношение потребителя и его пищи. Примером такого рода взаимоотношений могут служить «лемминговые циклы», сопровождающиеся катастрофическими нарушениями тундровой растительности, массовыми миграциями и гибелью животных. Но циклические изменения их численности связаны не только с нехваткой пищи и её последующим восстановлением. Выяснилось, что в период пика численности наблюдаются увеличение агрессии особей, возникновение стресса, что ведёт к снижению рождаемости через механизмы популяционного гомеостаза. Наиболее изучены циклические изменения численности популяций хищников и их жертв. В лабораторных условиях при совместном содержании хищника и его жертвы наблюдаются периодические изменения обоих видов, имеющие довольно закономерный характер (рис. 15).

Рис. 15. Взаимосвязанные колебания численности в системе хищник – жертва по модели Лотки – Вольтерра: N1 – жертва, N2 – хищник
Прежде всего это относится к специализированным хищникам, которые не могут переключиться на другие виды пищи при снижении численности их основной жертвы. Увеличение численности жертв идёт до тех пор, пока темп их выедания нарастающим количеством хищников не превысит скорость их размножения. После этого дальнейшая деятельность хищников подрывает кормовую базу, что ведёт к снижению их собственной численности. Однако в природных условиях такие закономерные циклические изменения численности в системе хищник – жертва выявить трудно, так как реальная динамика численности идёт под влиянием многих факторов. Иногда такие колебания обнаруживаются в бедных сообществах, где межвидовые связи не столь разнообразны: в лесах с господством одной древесной породы, в тундре, арктических пустынях и др.
Примерно по такой же схеме происходит динамика численности популяций при взаимодействии паразитов и возбудителей болезней с их хозяевами. При повышении плотности популяций возрастает вероятность заражения возбудителями болезней. В результате вспышки заболевания происходит вымирание животных, что ведёт к снижению численности возбудителя, который в выживших животных интенсивно размножается, увеличивая свою численность, и вновь вызывает вспышку заболевания.
Однако в природе такие закономерные колебания численности в рассмотренных системах паразит – хозяин и возбудитель болезни – хозяин часто осуществляются на протяжении короткого промежутка времени, после чего следует отклонение от классической схемы. В связи с этим ряд учёных (Н.Н. Наумов, А.А. Максимов) полагают, что биоценотические взаимоотношения организмов лишь сдерживают чрезмерный рост численности популяций. Процесс динамики численности популяций очень сложный, в нём невозможно выделить один ведущий фактор или даже небольшое их число. Роль отдельных факторов, определяющих общий уровень численности и тенденции к её изменению, меняется в зависимости от условий местности и состояния популяции в разные сезоны и годы.
Гомеостаз популяций
Принцип гомеостаза заключается в поддержании динамического равновесия популяции со средой как биологической системой. Одна из функциональных категорий внутрипопуляционного гомеостаза – поддержание оптимальной плотности популяции.
В основе способностей популяций к гомеостазу лежат изменения физиологических особенностей роста, поведения каждой особи в ответ на увеличение или уменьшение числа членов популяции.
В основе внутрипопуляционного гомеостаза лежит внутривидовая конкуренция. У одних видов она может проявляться в жёсткой форме, приводя к гибели избытка особей, у других – в смягчённой, через понижение рождаемости. У растений жёсткие формы конкуренции проявляются в самоизреживании всходов. При их большой густоте часть растений погибает в результате угнетения более сильными соседями. У животных жёсткие формы регуляции плотности популяции проявляются в форме каннибализма. Такое явление наблюдается при недостатке ресурсов среди как беспозвоночных, так и позвоночных животных. Так, у многих птиц более старшие птенцы при недостатке корма съедают своих собратьев. В небольших водоёмах взрослые окуни при высокой плотности популяции поедают своих мальков.
Среди механизмов, задерживающих рост популяций, у многих видов большое распространение имеет химическая регуляция (у растений, микроорганизмов, кишечнополостных, моллюсков, ракообразных и др.). Например, головастики выделяют в воду вещества, которые задерживают рост других головастиков. Чем крупнее особь, тем сильнее она воздействует на более мелких. Цианобактерии способны выделять вещества, которые влияют на рост и деление клеток. В лесу молодое поколение не появляется под материнским пологом, или находится в угнетённом состоянии из-за корневых выделений взрослых растений.
Территориальное поведение животных также является эффективным механизмом сдерживания роста численности популяции. Закрепление индивидуальных участков, их охрана, не допускающая размножения на ней других особей, приводят к рациональному использованию территории. Избыточная часть особей популяции при этом не размножается и вынуждена выселяться за пределы занимаемой популяцией территории. Выселение как ответная реакция на растущую плотность популяции свойственно многим видам птиц и млекопитающих. Для некоторых видов с резкими колебаниями численности характерны массовые перемещения – нашествия, которые наблюдаются в годы вспышек размножения (у леммингов, крыс, белок и др.). У некоторых насекомых повышение плотности ведёт к изменению развития и появлению специфических расселяющихся форм: крылатых особей у тлей, стадных перелётных форм у саранчовых, длиннокрылых у цикад и у некоторых других видов. Гибель животных среди расселяющихся животных выше, чем среди оседлых. У некоторых полёвок потери популяции при расселении составляют 40–70%. У животных со сложной возрастной структурой популяций аналогом расселения является разделение стад на самостоятельно существующие группы.
При отклонении плотности популяции от оптимума, а также при любых отрицательных воздействиях среды часть особей может испытывать состояние напряжения – стресс. В состоянии стресса в организме животных происходят изменения в функционировании нервной системы и в гормональном равновесии организма. У самок в популяции учащаются нарушения овуляции, рано прекращается лактация, угасают инстинкты размножения и заботы о потомстве. Всё это приводит к снижению роста популяции. В состоянии стресса у животных понижается устойчивость к воздействию неблагоприятных факторов среды, что увеличивает смертность. Большую роль в стимуляции стрессового состояния играет агрессивное поведение животных. У многих видов в условиях высокой плотности популяции повышается уровень агрессивности, меняется реакция на особей противоположного пола и т.д. В некоторых случаях агрессия может выступать как прямой фактор ограничения численности. Так, в экспериментах с домовыми мышами было показано, что рост численности и плотности популяции всегда сопровождается увеличением числа драк. У некоторых видов мелких грызунов (полёвок, леммингов и др.) наблюдается прерывание беременности самок в присутствии «чужого» самца, т.е. в ситуации, которая учащается при повышении плотности популяции. Влияние агрессии на размножение и смертность опосредуется через структуру популяции. Например, исследования показали, что в популяции рыжих полёвок в Подмосковье уровень стресса зависит не только от численности, но и от соотношения оседлых зверьков и мигрантов. В составе внутрипопуляционных групп наблюдается неодинаковое проявление агрессии и стресса у разных категорий особей. Агрессия чаще свойственна взрослым и доминантам, а стресс всегда более выражен у низкоранговых и молодых особей.
Таким образом, физиологические механизмы регуляции плотности популяции функционируют на базе разнокачественности особей в составе популяции и отдельных внутрипопуляционных групп. В результате действие стресса регулирует размножение и смертность в зависимости от остроты внутрипопуляционных отношений и степени неблагоприятности внешних условий.
Контрольные вопросы и задания
1. Дайте определение популяции. Каковы особенности элементарных, экологических и географических популяций?
2. Что понимается под половой структурой популяции? Назовите типы половой структуры популяций и их экологическое значение.
3. Каково экологическое значение возрастной структуры популяции и какие факторы влияют на её формирование?
4. Охарактеризуйте основные типы возрастной структуры популяций.
5. У какой популяции растений больше шансов на выживание: у той, которая состоит из одних проростков, или из проростков, молодых и взрослых особей? Ответ обоснуйте.
6. Назовите периоды и возрастные состояния в жизненном цикле растений.
7. Назовите основные типы пространственной структуры популяций. Чем объясняется преобладание в природе неравномерного распределения особей популяции на занимаемом пространстве? Объясните экологическое значение пространственной структуры популяции.
8. Каковы особенности территориального поведения оседлых и кочевых животных? Какой экологический смысл заключается в территориальном поведении оседлых животных?
9. Назовите основные формы совместного существования особей в популяции. Укажите особенности взаимоотношений животных в семьях, колониях, стаях и стадах.
10. Что такое репродуктивный (биотический) потенциал популяции (вида)? Почему он не реализуется полностью в природных условиях?
11. Каковы экологические причины, вызывающие рост численности популяции по экспоненциальной и по логистической кривым? Приведите примеры роста численности популяций по экспоненциальной и по логистической моделям.
12. Назовите типы динамики численности популяций у животных. Укажите, для популяций каких видов они характерны.
13. Какие факторы среды ответственны за изменения численности популяций? Каковы особенности действия модифицирующих и регулирующих факторов? Приведите примеры их действия на численность популяций.
14. Что такое гомеостаз популяции? Назовите основные внутрипопуляционные механизмы регуляции численности популяции.
15. Что нужно знать о виде, чтобы с достаточной вероятностью прогнозировать его численность?
16. Почему в сообществах, созданных человеком (посевы, парки), чаще наблюдается рост численности насекомых-вредителей, чем в природных сообществах?
ГЛАВА 5. Биоценозы
Понятие о биоценозе
Каждая популяция в природе существует в окружении множества других, вступая с ними в разнообразные отношения. Совокупность популяций всех видов организмов, совместно населяющих относительно однородное по абиотическим факторам среды пространство и взаимодействующих между собой, называют биоценозом. Термин «биоценоз» (от лат. биос – жизнь, ценоз – общий) предложен в 1877 г. немецким учёным К. Мёбиусом, изучавшим места обитания устриц в Северном море. Согласно Мёбиусу, возможность разных видов длительно существовать друг с другом в одном биоценозе, сложившаяся в процессе исторического развития видов, представляет результат естественного отбора. Приспособленность членов биоценоза к совместной жизни выражается в сходстве требований к важнейшим абиотическим условиям среды и закономерных отношениях друг с другом. Масштабы биоценозов различны: от сообщества лишайников на стволах деревьев до населения целых ландшафтов (лесов, степей, пустынь и т.д.). На суше биоценозы обычно выделяют по границам растительных ассоциаций, например биоценозы сосняка-черничника, ельника-кисличника, злаково-разнотравный и т.д. В водной среде различают биоценозы, соответствующие экологическим подразделениям частей водоёмов, например биоценозы прибрежных песчаных грунтов, абиссальных глубин и др. По отношению к более мелким сообществам (населению стволов или листвы деревьев, моховых кочек на болоте, муравейников и т. д.) применяют такие термины, как «микросообщества», «биоценотические группировки» и др. Мелкие группировки входят в состав более крупных, а те, в свою очередь, являются частями сообществ ещё больших масштабов. То есть организация жизни на биоценотическом уровне иерархична. С увеличением масштабов сообществ увеличиваются их сложность и доля непрямых (косвенных) связей между видами.
Биоценозу как любой надорганизменной биологической системе характерны определённые закономерности сложения, функционирования и развития. Важнейшие из них следующие:
1. Сообщества всегда возникают из готовых частей (представителей различных видов или целых комплексов видов), имеющихся в окружающей среде.
2. Части сообщества заменимы. Один вид (или комплекс видов) может занять место другого со сходными экологическими требованиями без ущерба для всей системы.
3. Надорганизменная система существует в основном за счёт уравновешивания противоположно направленных сил. Интересы многих видов в биоценозе прямо противоположны. Например, хищники – антагонисты своих жертв, но тем не менее они живут вместе в рамках единого сообщества.
4. Сообщества основаны на количественной регуляции численности одних видов другими.
5. Размеры надорганизменных систем определяются внешними причинами.
Структура биоценоза
Структура любой системы – это закономерности в соотношении и связях её частей. Структура биоценоза многопланова, поэтому при её изучении выделяют различные аспекты.
Видовая структура биоценоза характеризует разнообразие в нём видов и соотношение их численности или массы. Каждый биоценоз имеет определённый видовой состав, который зависит от условий среды, длительности его существования и истории формирования. Различают бедные и богатые видами биоценозы. Там, где условия абиотической среды приближаются к оптимальным для жизни, формируются богатые видами сообщества. Примерами являются влажные тропические леса, коралловые рифы, хвойно-широколиственные леса в лесной зоне и др. Где условия среды неблагоприятны для жизни большинства видов организмов, сообщества имеют бедный видовой состав, как, например, в арктических пустынях, в северной тундре, в глинистых пустынях и т. д.
Молодые, формирующиеся биоценозы имеют обычно меньший набор видов, чем давно сложившиеся. Сообщества, созданные человеком (поля, сады, парки), также беднее видами, чем сходные с ними природные биоценозы (луга, леса и др.). Однако даже самые бедные сообщества включают сотни видов организмов, принадлежащих к разным систематическим и экологическим группам. Богатые же биоценозы могут включать десятки тысяч видов.
В составе почти всех наземных и многих водных сообществ присутствуют растения, животные и микроорганизмы. Однако в некоторых условиях формируются биоценозы, в которых нет растений (например, в пещерах), или состоящие только из микроорганизмов (в анаэробной зоне водоёмов, иле на дне водоёмов и др.).
Видовое богатство биоценоза в значительной мере зависит от разнородности среды обитания. Чем разнообразнее условия среды, в которых формируется сообщество, тем более богатый их видовой состав, так как большее число видов может найти условия, соответствующие их экологическим требованиям. Например, на опушках леса формируются более богатые по видовому составу биоценозы, чем в глубине леса, так как здесь разнообразнее условия освещённости, температуры, влажности. Разнородность среды создаётся как абиотическими факторами, так и самими живыми организмами. Чем разнообразнее видовой состав и структура сообщества (фитоценоза), тем более разнообразны условия микроклимата в биоценозе, тем больше видов животных, грибов и микроорганизмов в нём может поселиться.
Численность организмов в биоценозах зависит от их размеров. Чем мельче особи видов, тем выше их численность в биоценозах. Так, например, обилие бактерий в почвах исчисляет






