Аэробный гликолиз.
В клетках глюкоза может превращаться в жирные кислоты, аминокислоты, гликоген и окисляться в различных катаболических путях.
Окисление глюкозы называют гликолизом. Глюкоза может окисляться до лактата и до пирувата. В аэробных условиях главным продуктом является пируват, такой путь называется аэробным гликолизом. При недостатке кислорода преобладает продукт - лактат. Этот путь окисления называется анаэробным гликолизом.
Процесс аэробного распада глюкозы можно разделить на три части: специфические для глюкоза превращения, завершающиеся образованием пирувата (аэробный гликолиз); общий путь катаболизма (окислительное декарбоксилирование и ЦЛК); дыхательная цепь.
В результате этих процессов глюкоза распадается до CO2 и H2O, а освобождающаяся энергия используется для синтеза АТФ.
Ферментативные реакции.
Распад глюкозы до пирувата также можно разделить на два этапа. Первый этап (глюкоза ® глицеральдегидфосфат) требует энергии в форме АТФ (2 АТФ).
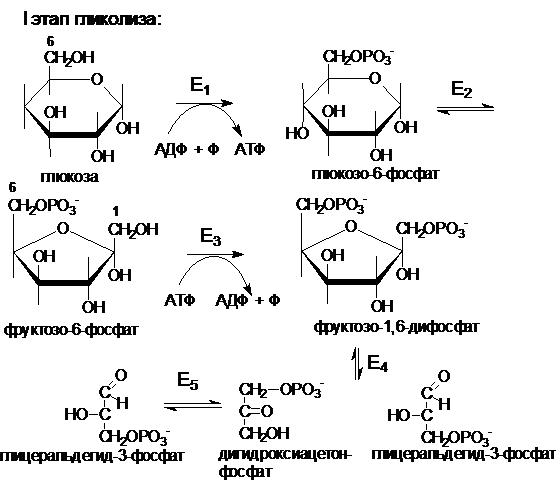
Е1 - гексокиназа или глюкокиназа
Е2 - глюкозофосфатизомераза
Е3 - фосфофруктокиназа
Е4 - фруктозодифосфатальдолаза
Е5 - триозфосфатизомераза
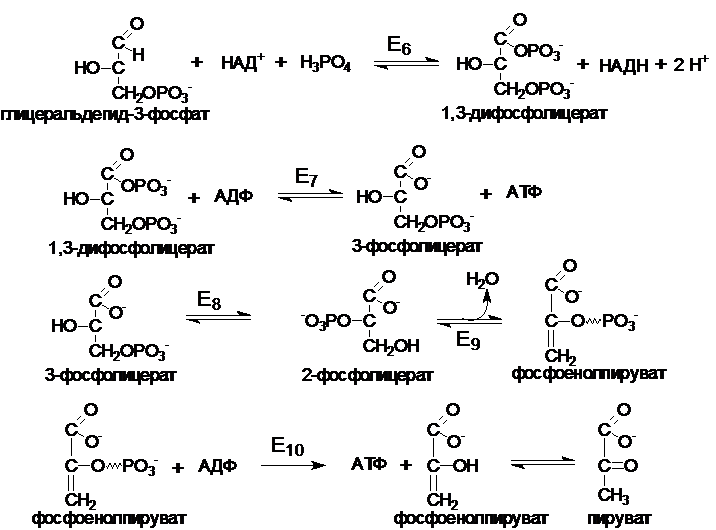
Второй этап (глицеральдегид ® пируват) протекает с выходом энергии в виде АТФ и НАДН (4 АТФ и 2 НАДН).
Е6 - глицеральдегид-3-фофатдегидрогеназа
Е7 - фосфоглицераткиназа
Е8 - фосфоглицератфосфомутаза
Е9 - енолгидратаза
Е10 - прируваткиназа
Суммарное уравнение гликолиза
Глюкоза + 2 АДФ + 2 НАД+ + 2 H3PO4 ® 2 Пируват + 2АТФ + 2 НАДН + 2 Н+
Челночные механизмы.
Перенос водорода с цитозольного НАДН в митохондрии происходит при участии специальных механизмов, называющихся челночными. Суть этих механизмов сводится к тому, что НАДН в цитозоле восстанавливает некоторое соединение, способное проникать в митохондрию; в митохондрии это соединение окисляется, восстанавливая внутримитохондриальный НАД+, и вновь переходит в цитозоль. Самая активная малат-аспартатная система, действующая в митохондриях печени, почек и сердца. На каждую пару электронов цитозольной НАДН, переданную на кислород по этой системе, образуется 3 молекулы АТФ.
В скелетных мышцах и мозге перенос восстановительных эквивалентов от цитозольной НАДН осуществляет глицеролфосфатная система. При этом восстановительные эквиваленты передаются в цепь переноса электронов через комплекс II, и поэтому синтезируется только 2 молекулы АТФ.
Значение ПФП.
Большая часть глюкозы расщепляется по гликолитическому пути. Наряду с гликолизом существуют второстепенные пути катаболизма глюкозы, одним из которых является пентозофосфатный путь (ПФП). Пентозофосфатный путь скорее даже - анаболический путь, который использует 6 атомов С глюкозы для синтеза пентоз и восстановительных эквивалентов в виде НАДФН, необходимых для образования липидов в организме. Однако, этот путь - окисление глюкозы, и при определенных условиях может завершиться полным окислением глюкозы до СО2 и Н2О. Главные функции этого цикла следующие:
Генерирование восстановительных эквивалентов в виде НАДФН для восстановительных реакций биосинтеза. Ферменты, которые катализируют реакции восстановления, часто используют в качестве кофактора НАДФ+/НАДФН, а окислительные ферменты - НАД+/НАДН. Восстановительные реакции биосинтеза жирных кислот и стероидных гормонов требуют НАДФН, поэтому клетки печени, жировой ткани и др., где протекает синтез липидов, имеют высокий уровень ферментов ПФП. В печени около 30% глюкозы подвергаются превращениям в ПФП.
НАДФН используется при обезвреживании лекарств и чужеродных веществ в монооксигеназной цепи окисления в печени.
НАДФН предохраняет ненасыщенные жирные кислоты эритроцитарной мембраны от аномальных взаимодействий с активным кислородом. Кроме того, эритроциты используют реакции ПФП для генерирования больших количеств НАДФН, который необходим для восстановления глутатиона, участвующего в системе обезвреживания активного кислорода, что позволяет поддерживать нормальную степень окисления Fe2+ в гемоглобине.
Снабжение клеток рибозо-5-фосфатом для синтеза нуклеотидов, нуклеотидных коферментов и нуклеиновых кислот. Превращение рибонуклеотидов в дезоксирибонуклеотиды также требует НАДФН, поэтому многие быстро пролиферирующие клетки требуют больших количеств НАДФН.
Хотя это не значительная функция ПФП, он принимает участие в метаболизме пищевых пентоз, входящих в состав нуклеиновых кислот пищи и может превращать их углеродный скелет в интермедиаты гликолиза и глюконеогенеза.
Реакции ПФП.
Реакции ПФП протекают в цитоплазме. ПФП можно разделить на два этапа: окислительный и неокислительный. Субстратом является глюкозо-6-фосфат. Первая и третья реакции катализируются НАДФ-зависимыми дегидрогеназами. На этом этапе происходит окисление глюкозы с образованием 2 НАФН, поэтому реакции (1 - 3) - окислительная фаза ПФП. Фермент Е1 (глюкозо-6-фосфат-дегидрогеназа) - ключевой фермент ПФП, катализирует образование 6-глюконолактона. Лактон является неустойчивым соединением, которое легко гидролизуется спонтанно и под действием фермента Е2(специфической лактоназы). Фермент Е3 (6-фосфоглюконат-дегидрогеназа) катализирует реакцию окислительного декарбоксилирования.
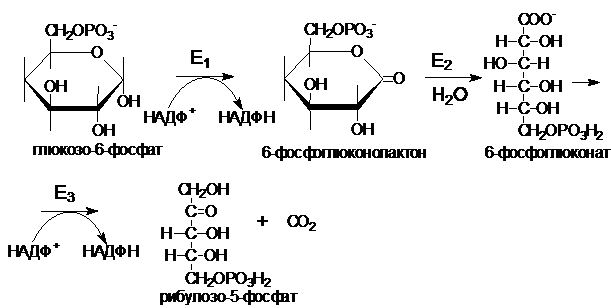
Рибулозо-5-фосфат под действием ферментов рибозофосфатизомеразы и рибулозо-5-фосфат-3-эпимеразы может обратимо изомеризоваться в другие пентозы: рибозо-5-фосфат, ксилулозо-5-фосфат.
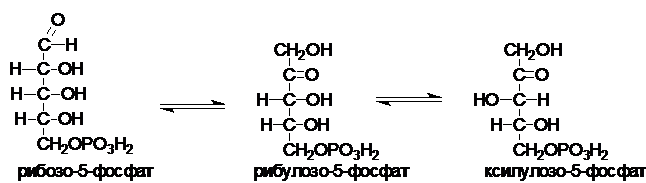
В некоторых тканях ПФП на этом заканчивается Суммарное уравнение ПФП:
глюкозо-6-фосфат + Н2О + НАДФ+ ® рибозо-5-фосфат + СО2 + НАДФН×Н+
Некоторые клетки нуждаются в большем количестве НАДФН, по сравнению с рибозо-5-фосфатом, т.е. НАФДН утилизируется, а рибозо-5-фосфат остается в избытке. В таких случаях рибозо-5-фосфат превращается в глицеральдегид-3-фосфат и фруктозо-6-фосфат (второй этап). Этот этап связывает ПФП с гликолизом.
Эти превращения катализируются двумя ферментами: транскетолазой (Е4) и трансальдолазой (Е5).
I
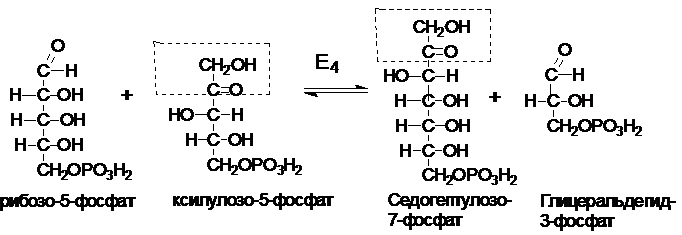
II

III
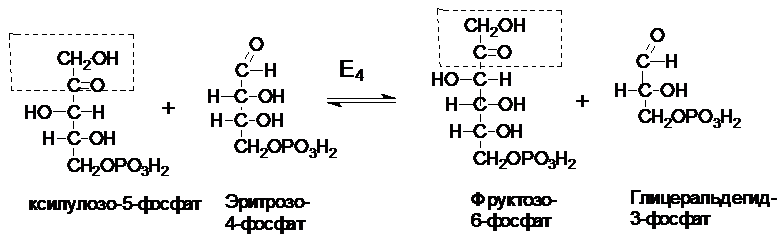
Фруктозо-6-фосфат и глицеральдегид-3-фосфат вступают на путь гликолиза.
Суммарное уравнение 2-го этапа:
рибозо-5-фосфат + 2 ксилулоза-5-фосфат ® 2 фруктозо-6-фосфат + глицеральдегид-3-фосфат
Общая характеристика метаболизм гликогена.
Так как глюкоза свободно диффундирует через клеточные мембраны, она не может служить резервным источником энергии в клетках тканей. Фосфорилирование глюкозы задерживает ее в клетке, но накапливаться глюкозо-6-фосфат не может, т.к. при этом увеличилось бы количество проникающей в клетку воды за счет осмоса. Создание запаса недиффундирующей глюкозы без осмотического набухания возможно благодаря способности некоторых тканей (печени и мышц) превращать избыток углеводов в нерастворимый полимер - гликоген.
Аэробный гликолиз.
В клетках глюкоза может превращаться в жирные кислоты, аминокислоты, гликоген и окисляться в различных катаболических путях.
Окисление глюкозы называют гликолизом. Глюкоза может окисляться до лактата и до пирувата. В аэробных условиях главным продуктом является пируват, такой путь называется аэробным гликолизом. При недостатке кислорода преобладает продукт - лактат. Этот путь окисления называется анаэробным гликолизом.
Процесс аэробного распада глюкозы можно разделить на три части: специфические для глюкоза превращения, завершающиеся образованием пирувата (аэробный гликолиз); общий путь катаболизма (окислительное декарбоксилирование и ЦЛК); дыхательная цепь.
В результате этих процессов глюкоза распадается до CO2 и H2O, а освобождающаяся энергия используется для синтеза АТФ.
Ферментативные реакции.
Распад глюкозы до пирувата также можно разделить на два этапа. Первый этап (глюкоза ® глицеральдегидфосфат) требует энергии в форме АТФ (2 АТФ).
Е1 - гексокиназа или глюкокиназа
Е2 - глюкозофосфатизомераза
Е3 - фосфофруктокиназа
Е4 - фруктозодифосфатальдолаза
Е5 - триозфосфатизомераза
Второй этап (глицеральдегид ® пируват) протекает с выходом энергии в виде АТФ и НАДН (4 АТФ и 2 НАДН).
Е6 - глицеральдегид-3-фофатдегидрогеназа
Е7 - фосфоглицераткиназа
Е8 - фосфоглицератфосфомутаза
Е9 - енолгидратаза
Е10 - прируваткиназа
Особенности ферментов гликолиза.
На пути гликолиза три реакции являются необратимыми (реакция 1 - глюкокиназная, реакция 3 - фофофруктокиназная, реакция 10 - пируваткиназная). Они катализируются регуляторными ферментами и определяют скорость всего процесса гликолиза. Кроме того, именно эти реакции отличаются от реакций обратного пути - синтеза глюкозы (глюконеогенеза).
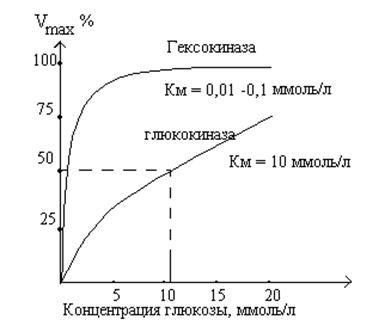
Гексокиназа и глюкокиназа
Глюкокиназная реакция - первая АТФ-зависимая реакция гликолиза. Она катализируется тканеспецифическими ферментами - гексокиназами. У человека известно 4 изомера гексокиназ (типы I - IV). Изофермент IV типа - глюкокиназа. Глюкокиназа находится только в печени и имеет высокое значение Км к глюкозе. Это приводит к тому, что фермент насыщается субстратом лишь при очень высоких концентрациях глюкозы. Гексокиназа катализирует фосфорилирование глюкозы при любых (в том числе низких) концентрациях глюкозы и ингибируется продуктом глюкозо-6-фосфатом. Глюкокиназа не ингибируется глюкозо-6-фосфатом. При увеличении концентрации глюкозы после приема пищи увеличивается скорость глюкокиназной реакции. Глюкозо-6-фосфат не проходит через клеточные мембраны и задерживается в клетке, поэтому больше глюкозы задерживается в печени. Таким образом, глюкокиназа является буфером глюкозы в крови. В тоже время, в тканях, энергетический обмен которых зависит от глюкозы, локализован изофермент с низким значением Км.
Глюкозофосфатизомераза
Фермент имеет почти равное значение Км для глюкозо-6-фосфата и фруктозо-6-фосфата. Этот фермент по-другому называют гексозофосфатизомераза. Катализирует реакции гликолиза и глюконеогенеза.
Фосфофруктокиназа
Этот фермент катализирует только прямую реакцию, т.е. эта реакция гликолиза является необратимой и определяет скорость всего процесса.
Фруктозодифосфатальдолаза катализирует реакции гликолиза и глюконеогенеза.
Триофосфатизомераза катализирует равновесную реакцию, и равновесие смещается в сторону гликолиза или глюконеогенеза по принципу действия масс.
Глицеральдегид-3-фосфатдегидрогеназа катализирует реакции гликолиза и глюконеогенеза.
Фосфоглицераткиназа катализирует обратимую реакцию (гликолиза и глюконеогенеза). Эта реакция имеет большое значение в эритроцитах, т.к. образующийся 1,3-дифосфоглицерат под действием фермента дифосфоглицератмутазы превращается в 2,3-дифосфоглицерат (ДФГ) - регулятор сродства Hb к кислороду.
Фосфоглицератфосфомутаза и енолгидратаза катализируют превращение относительно низкоэнергетической связи в 3-фосфоглицерате в высокоэнергетическую форму, а затем в АТФ.
Пируваткиназа - регуляторный фермент, который катализирует необратимую реакцию, в которой высокоэнергетический фосфат фосфоенолпирувата превращается в АТФ.
Пируват далее окисляется в митохондриях. Распад глюкозы до пирувата протекает в цитоплазме, поэтому существует специальный переносчик пирувата в митохондрии по механизму симпорта с Н+. Образующийся НАДН также должен быть транспортирован в митоходрии для окисления в цепи переноса электронов.






