

Общие условия выбора системы дренажа: Система дренажа выбирается в зависимости от характера защищаемого...

Особенности сооружения опор в сложных условиях: Сооружение ВЛ в районах с суровыми климатическими и тяжелыми геологическими условиями...
Общие условия выбора системы дренажа: Система дренажа выбирается в зависимости от характера защищаемого...
Особенности сооружения опор в сложных условиях: Сооружение ВЛ в районах с суровыми климатическими и тяжелыми геологическими условиями...
Топ:
Определение места расположения распределительного центра: Фирма реализует продукцию на рынках сбыта и имеет постоянных поставщиков в разных регионах. Увеличение объема продаж...
Эволюция кровеносной системы позвоночных животных: Биологическая эволюция – необратимый процесс исторического развития живой природы...
Генеалогическое древо Султанов Османской империи: Османские правители, вначале, будучи еще бейлербеями Анатолии, женились на дочерях византийских императоров...
Интересное:
Как мы говорим и как мы слушаем: общение можно сравнить с огромным зонтиком, под которым скрыто все...
Берегоукрепление оползневых склонов: На прибрежных склонах основной причиной развития оползневых процессов является подмыв водами рек естественных склонов...
Национальное богатство страны и его составляющие: для оценки элементов национального богатства используются...
Дисциплины:
|
из
5.00
|
Заказать работу |
|
|
|
|
ТЕМА 3.1. СТРОЕНИЕ И ФУНКЦИИ ДНК И РНК
1. ДНК и РНК представляют собой линейные полимеры, построенные из нуклеотидов. Каждый нуклеотид состоит из трех компонентов: азотистого основания, являющегося производным пурина или пиримидина, пентозы (рибозы или дезоксирибозы) и остатка фосфорной кислоты. В состав любой из нуклеиновых кислот входят два производных пурина (аденин, гуанин) и два производных пиримидина: в РНК - цитозин и урацил, а в ДНК - цитозин и тимин (табл. 3.1 и 3.2).
Таблица 3.1. Строение нуклеотидов РНК
| Азотистое основание | Пентоза | Нуклеозид | Нуклеотид | Однобуквенный код |
| Аденин | Рибоза | Аденозин | Аденозинмонофосфат (АМФ) | А |
| Гуанин | Рибоза | Гуанозин | Гуанозинмонофосфат (ГМФ) | G |
| Урацил | Рибоза | Уридин | Уридинмонофосфат (УМФ) | U |
| Цитозин | Рибоза | Цитидин | Цитидинмонофосфат (ЦМФ) | С |
Таблица 3.2. Строение нуклеотидов ДНК
| Азотистое основание | Пентоза | Нуклеозид | Нуклеотид | Однобуквенный код |
| Аденин | Дезоксирибоза | д-Аденозин | Дезоксиаденозинмонофосфат (дАМФ) | А |
| Гуанин | Дезоксирибоза | д-Гуанозин | Дезоксигуанозинмонофосфат (дГМФ) | G |
| Цитозин | Дезоксирибоза | д-Цитидин | Дезоксицитидинмонофосфат (дЦМФ) | С |
| Тимин | Дезоксирибоза | д-Тимидин | Дезокситимидинмонофосфат (дТМФ) | Т |
Первичная структура нуклеиновых кислот (НК) - это порядок чередования нуклеотидов в полинуклеотидной цепи, связанных между собой 3',5'-фосфодиэфирной связью. Образующиеся полимеры имеют фосфатный остаток на 5'-конце и свободную -ОН-группу пентозы на 3'-конце (рис. 3.1). Штрихами обозначают углеродные атомы пентозы для того, чтобы отличать их от атомов, входящих в азотистые основания.
Для краткого изображения последовательности нуклеотидов в нуклеиновых кислотах пользуются однобуквенным кодом. При этом запись осуществляют слева направо таким образом, что первый нуклеотид имеет свободный 5'-фосфатный конец, а последний -ОН-группу в 3'-положении рибозы или дезоксирибозы.
|
|
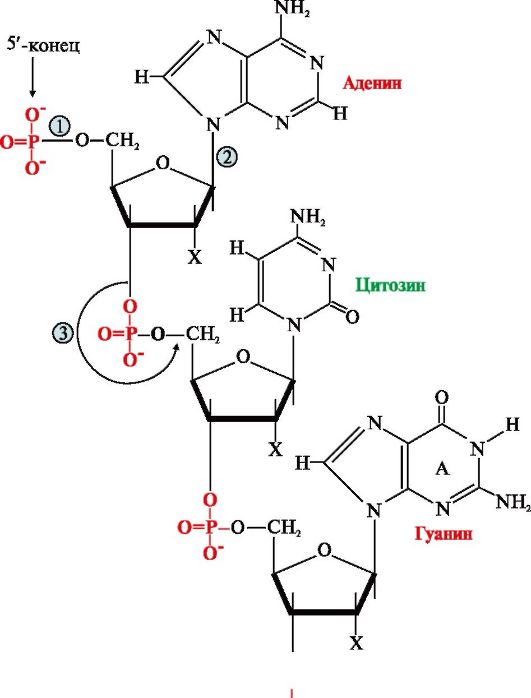
Рис. 3.1. Первичная структура нуклеиновых кислот.
Х = Н для ДНК, Х = ОН для РНК Связи в молекуле нуклеиновых кислот:
1 - 5'-фосфоэфирная; 2 - N-гликозидная; 3 - 3',5'-фосфодиэфирная
2. Пространственная структура ДНК. Вторичная структура представляет собой правозакрученную спираль (рис. 3.2), в которой две полинуклеотидные цепи расположены антипараллельно и удерживаются относительно друг друга за счет водородных связей между комплементарными азотистыми основаниями: А = Т и G = С.
Цепи молекулы ДНК не идентичны, но комплементарны друг другу: если известна первичная структура одной цепи, то последовательность нуклеотидов другой цепи задается правилом комплементарности оснований: Т одной цепи соответствует А, а С - G в другой цепи. Поэтому в молекуле ДНК количество адениловых нуклеотидов равно количеству тимидиловых нуклеотидов (А = Т), а количество гуаниловых равно количеству цитидиловых нуклеотидов (G = С). Соотношение А + Т / G + С - величина постоянная и является видоспецифической характеристикой организма. Основания нуклеотидов обращены внутрь молекулы и лежат в одной плоскости, которая практически перпендикулярна оси спирали. Между основаниями,
расположенными друг под другом, возникают гидрофобные взаимодействия. Дезоксирибозофосфатные остатки образуют остов спирали. На один виток спирали приходится 10 нуклеотидных пар.
Третичная структура ДНК формируется в результате ее взаимодействия с белками. Каждая молекула ДНК упакована в отдельную хромосому, в составе которой разнообразные белки связываются с отдельными участками ДНК и обеспечивают суперспирализацию и компактизацию молекулы. В период покоя комплексы ДНК с белками распределены равномерно по объему ядра, образуя хроматин. Белки хроматина включают две группы: гистоны и негистоновые белки.
|
|
Гистоны - небольшие белки с молекулярной массой от 11 000 до 22 000 Д и высоким содержанием лизина и аргинина. Четыре типа гистонов в количестве восьми молекул (по две каждого вида) образуют комплекс - нуклеосомный кор. Этот комплекс за счет ионных связей взаимодействует с отрицательно заряженными фосфатными группами участка ДНК длиной около 146 нуклеотидных пар (примерно 1,75 витка вокруг кора) и образует структуру, называемую нуклеосомой. Между нуклеосома-
ми находятся участки ДНК длиной около 30 нуклеотидных пар - линкерные участки, к которым присоединяются молекулы гистона Н1 (рис. 3.3).
Негистоновые белки представлены множеством ферментов и белков, участвующих в синтезе ДНК, РНК, регуляции этих процессов и компактизации
ДНК.
3. Пространственная структура РНК. В клетке существует три вида РНК: рибосомная (рРНК), транспортная (тРНК) и матричная (мРНК), каждая из которых выполняет свою особую функцию в синтезе белка.
Вторичная структура РНК формируется в результате спирализации отдельных участков одноцепочечной РНК. В спирализованных участках, или шпильках, между комплементарными парами азотистых оснований А и U, G и С возникают водородные связи. Двуцепочечные фрагменты чередуются с неспирализованными участками молекулы, образующими петли.
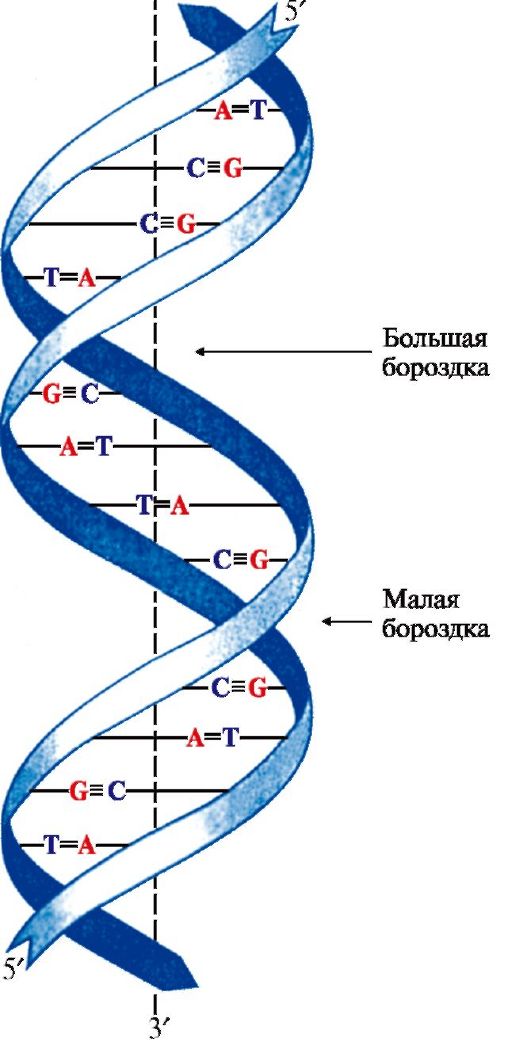 Рис. 3.2. Двойная спираль ДНК.
Рис. 3.2. Двойная спираль ДНК.
Аденин и тимин связывают две, а гуанин и цитозин - три водородные связи
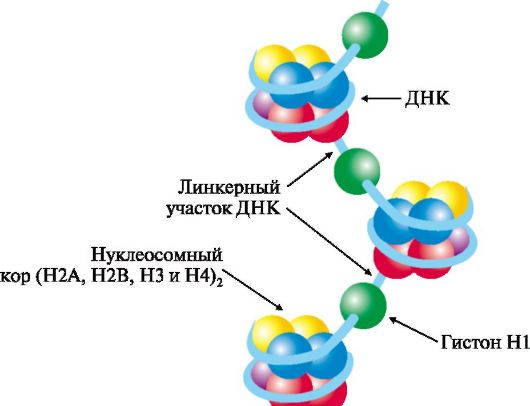 Рис. 3.3. Структура нуклеосом.
Рис. 3.3. Структура нуклеосом.
Восемь молекул гистонов четырех видов (Н2А, Н2В, Н3 и Н4) составляют нуклеосомный кор (ядро), на который наматывается ДНК, образуя примерно два витка
Третичная структура РНК образуется за счет дополнительных водородных связей между нуклеотидами неспирализованных участков, полинуклеотидной цепью рРНК или мРНК и белками, обеспечивает дополнительную компактизацию и стабилизацию пространственной структуры молекулы.
4. Структура молекул ДНК и РНК является видоспецифической характеристикой организмов. Об этом свидетельствуют эксперименты по гибридизации ДНК-ДНК и ДНК-РНК. При нагревании до 80-90°С нуклеиновые кислоты денатурируют с разрушением пространственной структуры, образуя одноцепочечные молекулы. При медленном охлаждении такие молекулы способны восстанавливать двойную спираль и либо приобретать исходную структуру, либо образовывать гибриды: совершенные или несовершенные в зависимости от комплементарности нитей друг другу по всей длине цепей. На этой способности двух образцов ДНК или ДНК и РНК образовывать гибриды в процессе охлаждения после денатурации основан метод молекулярной гибридизации.
|
|
Этот метод позволил установить следующие закономерности:
• ДНК всех клеток одного организма идентична, а ДНК разных организмов одного вида обнаруживает очень высокое сходство, обеспечивая образование «совершенных гибридов»;
• ДНК специфична для каждого вида и чем больше филогенетическая дистанция между видами, тем больше различий в строении принадлежащих им ДНК;
• ДНК, выделенная из тканей определенного организма, содержит информацию о структуре всех видов РНК данного организма.
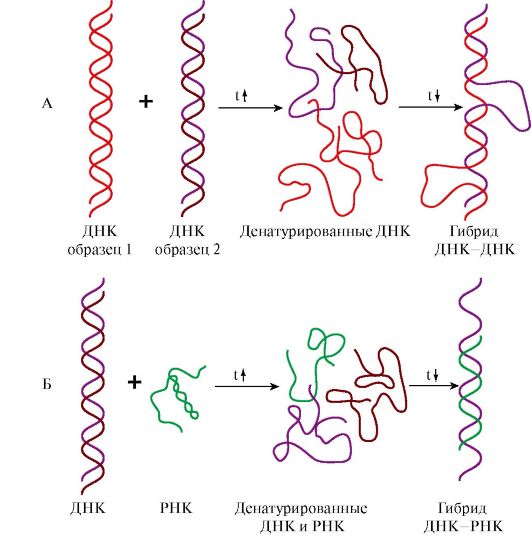 Рис. 3.4. Гибридизация нуклеиновых кислот:
Рис. 3.4. Гибридизация нуклеиновых кислот:
А - гибридизация ДНК-ДНК; Б - гибридизация ДНК-РНК
II. Синтез новых цепей ДНК.
ДНК.
I. Формирование репликативной вилки (рис. 3.5) идет при участии:
ДНК-топоизомеразы, которая является «обратимой нуклеазой». Сначала фермент разрывает 3'-,5'-фосфодиэфирную связь в одной из цепей ДНК
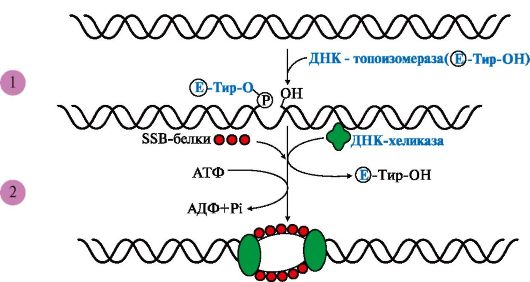 Рис. 3.5 Участие ДНК-топоизомеразы в образовании репликативной вилки.
Рис. 3.5 Участие ДНК-топоизомеразы в образовании репликативной вилки.
1 - фермент расщепляет 3',5'-фосфодиэфирную связь в одной из цепей ДНК и присоединяется к 5'-концу в точке разрыва; 2 - в область разрыва цепи присоединяются две молекулы ДНК-хеликаз и обеспечивают локальное разделение двойной спирали ДНК. ДНК-топоизомераза восстанавливает расщепленную 3',5'-фосфодиэфирную связь и отделяется, а к одноцепочечным участкам присоединяются SSB белки
и присоединяется к 5'-концу в точке разрыва, вызывая сброс суперспиралей ДНК-хроматина. Это облегчает присоединение в область разрыва цепи двух молекул ДНК-хеликаз и образование репликативной вилки. По окончании формирования репликативной вилки ДНК-топоизомераза восстанавливает целостность молекулы ДНК и отделяется;
ДНК-хеликаз - ДНК-зависимых АТФаз, использующих энергию АТФ для расплетения двойной спирали ДНК;
SSB (single strand binding)- белков, связывающихся с одноцепочечными участками ДНК. Эти белки, не закрывая оснований, предотвращают повторное комплементарное скручивание матричных цепей и образование шпилек.
|
|
II. Синтез новых цепей ДНК
На этой стадии дочерние нити ДНК образуются на обеих нитях материнской ДНК. Процесс катализирует несколько ДНК-полимераз, которые синтезируют полинуклеотидные цепи из дНТФ: дАТФ, дГТФ, дТТФ и дЦТФ в направлении от 5'- к 3'-концу антипараллельно матрице, имеющей направление от 3'- к 5'-концу (рис. 3.6).
Новые цепи синтезируются по-разному. На матрице ДНК с направлением от 3'- к 5'-концу цепь растет непрерывно по ходу движения репликативной вилки и называется лидирующей. На матрице с направлением от 3'- к 5'-концу вторая цепь синтезируется против движения репликативной вилки в виде коротких отрезков - фрагментов Оказаки. Рост этой цепи начинается только тогда, когда на матрице ДНК появляется одноцепочечный участок длиной около 200 нуклеотидов, поэтому ее называют запаздывающей или отстающей.
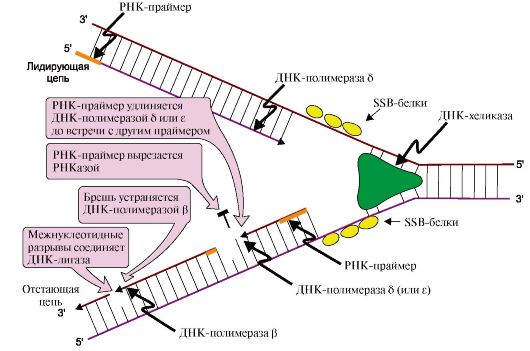 Рис. 3.6. Рост новых цепей в области репликативной вилки.
Рис. 3.6. Рост новых цепей в области репликативной вилки.
Лидирующая нить растет непрерывно, а отстающая - в виде фрагментов Оказаки, каждый из которых включает:
- РНК-праймер (~10 нуклеотидов);
- участок ДНК, примерно равный длине цепи из 150 нуклеотидов
ДНК-полимеразы δ, β и ε не способны инициировать синтез новых цепей ДНК, они могут лишь удлинять имеющуюся нуклеотидную цепь. Синтез лидирующей и отстающей нитей начинается с образования затравки или праймера -олигорибонуклеотида (РНК), включающего около 10 мононуклеотидов. Его образование катализирует праймаза - субъединица ДНК-полимеразы α. Далее этот же фермент, используя в качестве субстратов дНТФ, переключается на образование ДНК и включает во вновь синтезируемую нить 20-50 дезоксирибонуклеотидов, после чего заменяется другими ДНК-полимеразами. Синтез лидирующей цепи продолжает ДНК-полимераза δ, а отстающей - ДНК-полимераза δ или ε. Оба фермента, помимо, полимеразной обладают еще и экзонуклеазной активностью. В ходе синтеза они могут исправлять допущенную ошибку и отщеплять неправильно включенный нуклеотид, что обеспечивает высокую точность синтеза ДНК.
Таблица 3.3. Циклины, регулирующие прохождение клеточного цикла
| Циклин | Функция |
| D | Регулируют переход клетки из Gl-фазы в фазу S |
| Е, А | Активируют синтез ДНК на начальной стадии S-фазы |
| В | Регулируют переход клетки из G2-фазы в М-фазу |
ТЕМА 3.1. СТРОЕНИЕ И ФУНКЦИИ ДНК И РНК
1. ДНК и РНК представляют собой линейные полимеры, построенные из нуклеотидов. Каждый нуклеотид состоит из трех компонентов: азотистого основания, являющегося производным пурина или пиримидина, пентозы (рибозы или дезоксирибозы) и остатка фосфорной кислоты. В состав любой из нуклеиновых кислот входят два производных пурина (аденин, гуанин) и два производных пиримидина: в РНК - цитозин и урацил, а в ДНК - цитозин и тимин (табл. 3.1 и 3.2).
|
|
Таблица 3.1. Строение нуклеотидов РНК
| Азотистое основание | Пентоза | Нуклеозид | Нуклеотид | Однобуквенный код |
| Аденин | Рибоза | Аденозин | Аденозинмонофосфат (АМФ) | А |
| Гуанин | Рибоза | Гуанозин | Гуанозинмонофосфат (ГМФ) | G |
| Урацил | Рибоза | Уридин | Уридинмонофосфат (УМФ) | U |
| Цитозин | Рибоза | Цитидин | Цитидинмонофосфат (ЦМФ) | С |
Таблица 3.2. Строение нуклеотидов ДНК
| Азотистое основание | Пентоза | Нуклеозид | Нуклеотид | Однобуквенный код |
| Аденин | Дезоксирибоза | д-Аденозин | Дезоксиаденозинмонофосфат (дАМФ) | А |
| Гуанин | Дезоксирибоза | д-Гуанозин | Дезоксигуанозинмонофосфат (дГМФ) | G |
| Цитозин | Дезоксирибоза | д-Цитидин | Дезоксицитидинмонофосфат (дЦМФ) | С |
| Тимин | Дезоксирибоза | д-Тимидин | Дезокситимидинмонофосфат (дТМФ) | Т |
Первичная структура нуклеиновых кислот (НК) - это порядок чередования нуклеотидов в полинуклеотидной цепи, связанных между собой 3',5'-фосфодиэфирной связью. Образующиеся полимеры имеют фосфатный остаток на 5'-конце и свободную -ОН-группу пентозы на 3'-конце (рис. 3.1). Штрихами обозначают углеродные атомы пентозы для того, чтобы отличать их от атомов, входящих в азотистые основания.
Для краткого изображения последовательности нуклеотидов в нуклеиновых кислотах пользуются однобуквенным кодом. При этом запись осуществляют слева направо таким образом, что первый нуклеотид имеет свободный 5'-фосфатный конец, а последний -ОН-группу в 3'-положении рибозы или дезоксирибозы.
Рис. 3.1. Первичная структура нуклеиновых кислот.
Х = Н для ДНК, Х = ОН для РНК Связи в молекуле нуклеиновых кислот:
1 - 5'-фосфоэфирная; 2 - N-гликозидная; 3 - 3',5'-фосфодиэфирная
2. Пространственная структура ДНК. Вторичная структура представляет собой правозакрученную спираль (рис. 3.2), в которой две полинуклеотидные цепи расположены антипараллельно и удерживаются относительно друг друга за счет водородных связей между комплементарными азотистыми основаниями: А = Т и G = С.
Цепи молекулы ДНК не идентичны, но комплементарны друг другу: если известна первичная структура одной цепи, то последовательность нуклеотидов другой цепи задается правилом комплементарности оснований: Т одной цепи соответствует А, а С - G в другой цепи. Поэтому в молекуле ДНК количество адениловых нуклеотидов равно количеству тимидиловых нуклеотидов (А = Т), а количество гуаниловых равно количеству цитидиловых нуклеотидов (G = С). Соотношение А + Т / G + С - величина постоянная и является видоспецифической характеристикой организма. Основания нуклеотидов обращены внутрь молекулы и лежат в одной плоскости, которая практически перпендикулярна оси спирали. Между основаниями,
расположенными друг под другом, возникают гидрофобные взаимодействия. Дезоксирибозофосфатные остатки образуют остов спирали. На один виток спирали приходится 10 нуклеотидных пар.
Третичная структура ДНК формируется в результате ее взаимодействия с белками. Каждая молекула ДНК упакована в отдельную хромосому, в составе которой разнообразные белки связываются с отдельными участками ДНК и обеспечивают суперспирализацию и компактизацию молекулы. В период покоя комплексы ДНК с белками распределены равномерно по объему ядра, образуя хроматин. Белки хроматина включают две группы: гистоны и негистоновые белки.
Гистоны - небольшие белки с молекулярной массой от 11 000 до 22 000 Д и высоким содержанием лизина и аргинина. Четыре типа гистонов в количестве восьми молекул (по две каждого вида) образуют комплекс - нуклеосомный кор. Этот комплекс за счет ионных связей взаимодействует с отрицательно заряженными фосфатными группами участка ДНК длиной около 146 нуклеотидных пар (примерно 1,75 витка вокруг кора) и образует структуру, называемую нуклеосомой. Между нуклеосома-
ми находятся участки ДНК длиной около 30 нуклеотидных пар - линкерные участки, к которым присоединяются молекулы гистона Н1 (рис. 3.3).
Негистоновые белки представлены множеством ферментов и белков, участвующих в синтезе ДНК, РНК, регуляции этих процессов и компактизации
ДНК.
3. Пространственная структура РНК. В клетке существует три вида РНК: рибосомная (рРНК), транспортная (тРНК) и матричная (мРНК), каждая из которых выполняет свою особую функцию в синтезе белка.
Вторичная структура РНК формируется в результате спирализации отдельных участков одноцепочечной РНК. В спирализованных участках, или шпильках, между комплементарными парами азотистых оснований А и U, G и С возникают водородные связи. Двуцепочечные фрагменты чередуются с неспирализованными участками молекулы, образующими петли.
Рис. 3.2. Двойная спираль ДНК.
Аденин и тимин связывают две, а гуанин и цитозин - три водородные связи
Рис. 3.3. Структура нуклеосом.
Восемь молекул гистонов четырех видов (Н2А, Н2В, Н3 и Н4) составляют нуклеосомный кор (ядро), на который наматывается ДНК, образуя примерно два витка
Третичная структура РНК образуется за счет дополнительных водородных связей между нуклеотидами неспирализованных участков, полинуклеотидной цепью рРНК или мРНК и белками, обеспечивает дополнительную компактизацию и стабилизацию пространственной структуры молекулы.
4. Структура молекул ДНК и РНК является видоспецифической характеристикой организмов. Об этом свидетельствуют эксперименты по гибридизации ДНК-ДНК и ДНК-РНК. При нагревании до 80-90°С нуклеиновые кислоты денатурируют с разрушением пространственной структуры, образуя одноцепочечные молекулы. При медленном охлаждении такие молекулы способны восстанавливать двойную спираль и либо приобретать исходную структуру, либо образовывать гибриды: совершенные или несовершенные в зависимости от комплементарности нитей друг другу по всей длине цепей. На этой способности двух образцов ДНК или ДНК и РНК образовывать гибриды в процессе охлаждения после денатурации основан метод молекулярной гибридизации.
Этот метод позволил установить следующие закономерности:
• ДНК всех клеток одного организма идентична, а ДНК разных организмов одного вида обнаруживает очень высокое сходство, обеспечивая образование «совершенных гибридов»;
• ДНК специфична для каждого вида и чем больше филогенетическая дистанция между видами, тем больше различий в строении принадлежащих им ДНК;
• ДНК, выделенная из тканей определенного организма, содержит информацию о структуре всех видов РНК данного организма.
Рис. 3.4. Гибридизация нуклеиновых кислот:
А - гибридизация ДНК-ДНК; Б - гибридизация ДНК-РНК
|
|
|

Типы оградительных сооружений в морском порту: По расположению оградительных сооружений в плане различают волноломы, обе оконечности...

Папиллярные узоры пальцев рук - маркер спортивных способностей: дерматоглифические признаки формируются на 3-5 месяце беременности, не изменяются в течение жизни...

Индивидуальные очистные сооружения: К классу индивидуальных очистных сооружений относят сооружения, пропускная способность которых...

Типы сооружений для обработки осадков: Септиками называются сооружения, в которых одновременно происходят осветление сточной жидкости...
© cyberpedia.su 2017-2024 - Не является автором материалов. Исключительное право сохранено за автором текста.
Если вы не хотите, чтобы данный материал был у нас на сайте, перейдите по ссылке: Нарушение авторских прав. Мы поможем в написании вашей работы!