ЭНЕРГЕТИЧЕСКОЕ ОБЕСПЕЧЕНИЕ МЫШЕЧНОГО СОКРАЩЕНИЯ.
Непосредственным источником энергии для мышечной деятельности служит реакция расщепления АТФ. При ферментативном гидролизе АТФ происходит освобождение энергии, преобразуемой в процессе мышечного сокращения в механическую работу.
Запас АТФ в мышцах невелик. Для поддержания активности тканей на определенном уровне неоходим быстры ресинтез АТФ. Последний происходит в процессе рефосфорилирования при соединении АДФ и фосфатов. Наиболее доступным веществом является креатин-фосфат: КФ + АДФ = Креатин +АТФ
Концентрация КФ в мышцах в 2-4 раза больше в
Аденозинтрифосфат (АТФ) является нуклеотидом. В состав молекулы АТФ входят азотистое основание - аденин, углевод - рибоза и три остатка фосфорной кислоты (аденин, связанный с рибозой, называется аденозином).
Особенностью молекулы АТФ является то, что второй и третий остатки фосфорной кислоты присоединяются связью, богатой энергией. Такая связь называется высокоэнергетической, или макроэргической, и обозначается знаком Соединения, имеющие макроэргические связи, обозначаются термином «макроэрги».
Структурная формула АТФ имеет следующий вид:
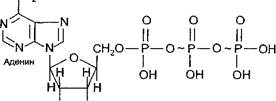
В упрощенном виде строение АТФ можно отразить схемой: Аденин — рибоза.— Ф.К. ~ Ф.К. ~ Ф.К.
Y
аденозин
При использовании АТФ в качестве источника энергии обычно происходит отщепление путем гидролиза последнего остатка фосфорной кислоты:
АТФ + Н20 -> АДФ + Н3Р04 + Q (энергия)
В физиологических условиях, т. е. при тех условиях, которые имеются в живой клетке (температура, рН, осмотическое давление, концентрация реагирующих веществ и пр.), расщепление моля АТФ (506 г) сопровождается выделением 12 ккал, или 50 кДж* энергии.
Основным поставщиком АТФ является тканевое дыхание - завершающий этап катаболизма, протекающий в митохондриях всех клеток, кроме красных клеток крови (эритроцитов).
Содержание АТФ в мышце относительно постоянно. Концентрация АТФ составляет около 5 мм на 1 кг сырого веса мышц (около 25%). Накапливать большое количество АТФ мышца не может, т.к. при этом возникает субстратное угнетение миозиновой АТФ-азн, препятствующее образованию спаек между актиновыми и миозановыми нитями в миофибриллах и ведущее к утрачиванию сократительной способности мышцы. Вместе с тем концентрация АТФ в мышце не может снижаться ниже 2 мм на 1 кг сырого веса ткани, поскольку при этом перестанет действовать "кальциевый насос" в пузырьках, и мышца будет сокращаться вплоть до полного исчерпания всех запасов АТФ и развития тригора (состояния стойкого непроходящего сокращения).
Запасов АТФ в мышце обычно хватает на 3-4 одиночных сокращения максимальной силы. В то же время, как показывают исследования с использованием микробиопсии мышц, в процессе мышечной работы не наблюдается значительного снижения концентрации АТФ. Это объясняется тем, что по ходу мышечной деятельности АТФ восстанавливается из продуктов распада (ресинтезируется) с той же скоростью, с какой она расщепляется в процессе мышечных сокращений. Для обращения реакции гидролиза АТФ конечные продукты распада –АДФ и Н3РО4 – должны получить из реакционной среды энергию, равную той, что выделилась при распаде АТФ. Следовательно, синтез АТФ должен быть сопряжен с каталитическими реакциями, при которых освобождается значительное количество потенциальной химической энергии.
Ресинтез АТФ при мышечной деятельности может осуществляться
как в ходе реакции, идущих без кислорода, так и за счет окислительных превращений в клетках, связанных с потреблением кислорода. В обычных условиях ресинтез АТФ происходит в основном путем аэробных превращений, но при напряженной мышечной деятельности, когда доставка кислорода к мышцам затруднена, в тканях одновременно усиливаются анаэробные процессы ресинтеза АТФ.
Однако есть и недостатки.
1. Обязательное потребление кислорода, что ограничено скоростью доставки кислорода в мышцы и скоростью проникновения кислорода через мембрану митохондрий.
2. Большое время развертывания.
3. Небольшую по максимальной величине мощность.
Поэтому мышечная деятельность, свойственная большинству видов спорта, не может быть полностью получена этим путем ресинтеза АТФ.
В спортивной практике для оценки аэробного ресинтеза используются следующие показатели: максимальное потребление кислорода (МПК), порог аэробного обмена (ПАО), порог анаэробного обмена (ПАНО) и кислородный приход.
МПК – это максимально возможная скорость потребления кислорода организмом при выполнение физической работы. Чем выше МПК, тем выше скорость тканевого дыхания. Чем тренированнее человек, тем выше МПК. МПК рассчитывают обычно на 1кг массы тела. У людей, не занимающихся спортом МПК 50 мл/мин-кг, а у тренированных людей он достигает 90 мл/мин-кг. В спортивной практике МПК также используется для характеристики относительной мощности аэробной работы, которая выражается в процентах от МПК. Например, относительная мощность работы, выполняемая с потреблением кислорода 3 л/мин спортсменом, имеющим МПК 6 л/мин, будет составлять 50% от уровня МПК.
ПАО – это наибольшая относительная мощность работы, измеряемая по потреблению кислорода в процентах по отношению к МПК. Большие величины ПАО говорят о лучшем развитии аэробного ресинтеза. ПАНО – это минимальная относительная мощность работы, также измеренная по потреблению кислорода в процентах по отношению к МПК. Высокое ПАНО говорит о том, что аэробный ресинтез выше в единицу времени, поэтому гликолиз включается при гораздо больших нагрузках.
Кислородный приход – это количество кислорода (сверх дорабочего уровня), использованное во время выполнения данной нагрузки для обеспечения аэробного ресинтеза АТФ. Кислородный приход характеризует вклад тканевого дыхания в энергообеспечение всей проделанной работы. Кислородный приход часто используют для оценки всей проделанной аэробной работы.
Под влиянием систематических тренировок в мышечных клетках возрастает количество митохондрий, совершенствуется кислородно-транспортная функция организма, возрастет количество миоглобина в мышцах и гемоглобина в крови.
Стимуляция мышечных волокон
Высвобождение ацетилхолина вблизи двигательной концевой пластинки скелетной мышцы ведет к возникновению тока концевой пластинки, который распространяется электротонически и активирует быстрые потенциалзависимые Na+-каналы в сарколемме. Это ведет к возникновению потенциала действия (ПД), который проводится со скоростью 2 м/с вдоль сарколеммы всего мышечного волокна и быстро проникает в глубь волокна по Т-системе (А).
Генетические дефекты в структуре натриевых каналов замедляют их дезактивацию, что приводит к повышенной возбудимости с увеличением длительности сокращения и задержкой расслабления скелетной мышцы (миотония). Увеличение мышечной активности сопровождается массированным выходом ионов калия из волокна. Это приводит к гиперкалиемии, вследствие чего мышечный потенциал покоя достигает значений, при которых натриевые каналы более не могут быть активированы, и мышца оказывается временно парализована: семейный гилеркалиемический периодический паралич.
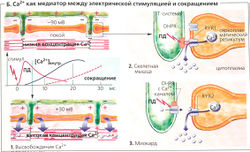

Б. Са2+ как медиатор между электрической стимуляцией и сокращением
Переход от возбуждения к сокращению поперечно-полосатой мышцы называется электромеханическим сопряжением (Б). В скелетной мышце этот процесс начинается с потенциала действия, возбуждающего зависимые от напряжения дигидропиридиновые рецепторы (DHPR) в сарколемме в районе триад. Эти рецепторы организованы в ряды, а напротив них в соседней мембране саркоплазматического ретикулума находятся ряды Са2+-каналов, называемых рианодиновыми рецепторами (RYR; в скелетных мышцах рианодиновый рецептор типа 1 - сокращенно RYR1). Каждый второй RYR1 ассоциирован с DHPR (Б2). RYR1 открываются, когда механически «ощущают» изменение конформации DHPR под действием потенциала действия. В миокарде каждый DHPR является частью потенциалзависимого Са2+-канала сарколеммы, который открывается в ответ на потенциал действия. Малые количества внеклеточного Са2+ входят в клетку через этот канал, приводя тем самым к открыванию миокардиального канала RYR2 (так называемый индуцирующий эффект Са2+, или «Са2+-вспышка», БЗ). Ионы Са2+, хранящиеся в СР, выходят через открытый RYR1 или RYR2 в цитоплазму, увеличивая цитоплазматическую концентрацию Са2+ до более чем 1 мкмоль/л по сравнению с концентрацией в покое —0,01 мкмоль/л (Б1). В скелетной мышце стимуляции DHPR на одном участке достаточно, чтобы запустить «дружное» открывание всей группы RYR1. Таким образом увеличивается надежность проведения импульса. Повышенная концентрация Са2+ в цитоплазме насыщает Са2+-связывающие сайты тропонина С, отменяя ингибиторный эффект тропомиозина на скольжение филаментов (Г), которое препятствует прочному (высокоаффинному) связыванию актина и миозина II.
У пациентов с генетическими дефектами RYR1 общая анестезия может приводить к массированному выбросу кальция, что вызывает сильные мышечные сокращения, сопровождающиеся стремительным и опасным для жизни повышением температуры тела: злокачественная гипертермия (=молниеносная гиперпирексия).

В. Скользящие филаменты
Молекулы АТФ необходимы для скольжения филаментов и, следовательно, для мышечного сокращения. Благодаря своей АТФазной активности миозиновые головки действуют как моторы (моторные белки) этого процесса. Миозиновые-ll и актиновые филаменты в саркомере организованы таким образом, что могут скользить друг по другу. Миозиновые головки соединяются с актиновыми филаментами под особым углом, образуя так называемые поперечные мостики (В1). Благодаря конформационным изменениям в районе нуклеотид-связывающего сайта миозина-ll, пространственные размеры которого увеличиваются согласованным движением участка шейки, миозиновая головка наклоняется, смещая за два последовательных «шага» тонкие филаменты в общей сложности на 4-12 нм (рабочий такт). Вторая миозиновая головка может также действует на соседний актиновый филамент, вызывая его сокращение. Затем головка отсоединяется и «напрягается», готовясь к следующему «гребку», когда свяжется с актином снова (ВЗ).
Кинезин, другой моторный белок, независимо двигается по микротрубочке посредством «шагания» двух своих головок (на 8 нм за цикл), как при перетягивании каната. В этом случае 50% цикла - «рабочее время» (коэффициент полезного действия 0,5). В скелетной мышце между двумя последовательными взаимодействиями с актином сам миозин-ll совершает «прыжок» на 36 нм (или на число нм, кратное 36, например, при быстром сокращении на 396 нм или более), чтобы достичь следующего (или 11-го) удобно расположенного актинсвязывающего сайта (ВЗ, прыжок от а к б). В то же время другие миозиновые головки, работающие на данном актиновом филаменте, должны сделать по крайней мере от 10 до 100 гребков примерно по 4 нм каждый. Коэффициент полезного действия головки миозина-ll, таким образом, от 0,1 до 0,01. Такое «разделение труда» между миозиновыми головками гарантирует, что некоторая доля миозиновых головок всегда готова совершить быстрое сокращение.
При скольжении филаментов Z-диски сближаются, и участки наложения тонких и толстых становятся шире, но их общая длина остается неизменной. Это приводит к укорачиванию 1-полосы и Н-зоны. Когда концы толстых филаментов «надвигаются» на Z-диск, происходит максимальное укорачивание мышцы и концы тонких филаментов перекрываются. Укорачивание саркомера, таким образом, происходит с обоих концов миозиновых пучков, но в противоположных направлениях.
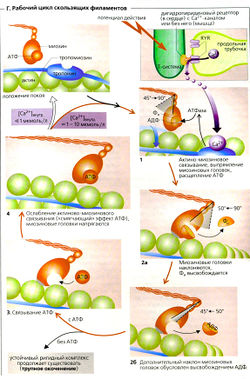
Г. Рабочий цикл скользящих филаментов
Механика скелетной мышцы
Потенциалы действия, генерируемые в мышечном волокне, увеличивают внутриклеточную концентрацию Са2+, [Са2+]внугр, инициируя сокращение (скелетная мышца; миокард). В скелетных мышцах регуляция силы сокращения достигается участием разного числа двигательных единиц и изменением частоты потенциала действия. Однократный стимул, если он выше порогового уровня, всегда ведет к максимальному высвобождению Са2+ и, таким образом, к максимально интенсивному однократному сокращению (реакция «все или ничего»). Тем не менее такой однократный стимул не индуцирует максимального укорочения мышечного волокна, поскольку он слишком короткий для поддержания скользящих филаментов в движении до достижения финальной позиции. Укорочение мышцы продолжается только в том случае, если второй стимул поступит до того, как мышца полностью расслабится после первого стимула. Такое повторение стимула ведет к нарастающей механической суммации, или суперпозиции, отдельных сокращений (А). Если частота стимуляции становится настолько высокой, что мышца уже не может расслабиться между стимулами, то наступает длительное максимальное сокращение двигательных единиц, или тетанус (А). Это происходит, например, при 20 Гц в медленно сокращающихся мышцах, и при 60-100 Гц в быстро сокращающихся мышцах. Мышечная сила во время тетануса может быть в четыре раза больше, чем при однократном сокращении скелетных мышц. Концентрация Са2+ несколько уменьшается между суммирующимися стимулами, а при тетанусе остается высокой.
Окоченение, как и контрактура, характеризуются стойким укорачиванием мышц. Это состояние нужно отличать от тетануса. Контрактура вызывается не потенциалом действия, а стойкой локальной деполяризацией, например, вследствие увеличенной внутриклеточной концентрации К+ (К+-контрактура) или индуцированного высвобождения Са2+, например, в ответ на кофеин. Сокращение так называемых изотонических волокон (особых волокон внешних мышц глаза и мышечных веретен; с. 326) также одна из форм контрактуры. Изотонические волокна не отвечают на стимулы по закону «все или ничего», а сокращаются пропорционально величине деполяризации. Степень сокращения изотонических волокон регулируется изменением концентрации Са2+ в цитоплазме (не потенциалом действия!).
И напротив, общий мышечный тонус (рефлекторный тонус), или устойчивое напряжение скелетной мышцы в покое, относится к развитию нормального потенциала действия в отдельной двигательной единице. Единичные сокращения не могут быть зарегистрированы, поскольку двигательные единицы работают асинхронно. Например, лозные мышцы (поддерживающие осанку) при видимом покое находятся в непроизвольном напряжении. Тонус покоящейся мышцы регулируется рефлексами и увеличивается при повышении внимания.
Типы сокращений
]

Б. Типы сокращений
Существуют разные типы мышечных сокращений. При изометрическом сокращении мышечная сила (напряжение) изменяется, а длина мышцы остается постоянной. (В сердечной мышце этот тип представлен изоволюметрическим (изообъемным) сокращением, ведь длина мышцы определяет объем предсердия и желудочков.) При изотоническом сокращении длина мышцы меняется под действием постоянной мышечной силы. (В сердечной мышце этот тип представлен изобарным сокращением (при постоянном давлении) - мышечная сила определяет давление в предсердии или в желудочке.) При ауксотоническом сокращении мышечная длина и сила изменяются одновременно. Изотоническое или ауксотоническое сокращение, которое формируется на основе изометрического, называется сокращением с постнагрузкой.
Растяжимость мышцы

В. Изометрическая мышечная сила при разной длине саркомера
Покоящаяся мышца, содержащая АТФ, может быть растянута, как будто она резиновая. Сила, необходимая для начала расслабления мышцы (Г, Д, сила покоя), очень мала, но увеличивается экспотенциально в случае эластичной мышцы (см. кривую покоя, Г). Мышечное сопротивление растяжению, которое удерживает скользящие филаменты в саркомере от разделения, в какой-то степени зависит от фасций (фиброзной ткани). Основной фактор, однако, это гигантская нитевидная эластичная молекула, называемая титином (или коннектином; длиной 1000 нм, массой от 3 до 3,7 МДа), которая включена в саркомер (6 молекул титина на миозиновый филамент). В районе полосы А каждого саркомера титин расположен около миозинового филамента и помогает удерживать его в центре саркомера. Молекулы титина в районе полосы I гибкие и функционируют в качестве «эластичных тяжей», которые противодействуют пассивному сокращению мышцы и влияют на скорость ее укорачивания.
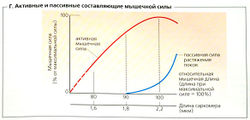
Г. Активные и пассивные составляющие мышечной силы
Растяжимость молекулы титина (титин может вытягиваться до примерно десятикратного размера по сравнению с первоначальной длиной в скелетной мышце и немного меньше в сердечной мышце) определяется частым повторением последовательности PEVK (пролин-глутамат-валин-лизин). При очень сильном растяжении мышцы, которое представлено самым крутым участком кривой покоя (Г), также разворачиваются элементы глобулярной цепи, называемые доменами иммуноглобулина С2. Чем быстрее сокращается мышца, тем более неожиданным и резким будет действие этого «поглотителя шока».

Д. Кривые «длина-сила» для скелетной и сердечной мышц
Длина (L) и сила (F), или «напряжение», мышцы тесно взаимосвязаны (В, Д). Общая сила мышцы является суммой ее активной силы и ее напряжения в покое, как было описано выше. Поскольку активная сила определяется величиной всех потенциальных актино-миозиновых взаимодействий, она варьирует в соответствии с начальной длиной саркомера (В, Г). Скелетная мышца может развить максимальную активную (изометрическую) силу (F0) от своей длины покоя (Lmax; длина саркомера примерно от 2 до 2,2 мкм; В). Когда саркомеры укорачиваются, (L < Lmax), часть тонких филаментов перекрывается - развиваемая сила меньше Fq (В). При L —0,7 /тах (длина саркомера 1,65 мкм) толстые филаменты контактируют с Z-диском - F еще меньше. Кроме того, способность предварительно растянутой мышцы [L > развивать силу ограничена, поскольку ограничено количество потенциально доступных актино-миозиновых мостиков (В). При растяжении до 130% или более величины Lmax сила растяжения в покое становится основной составляющей общей мышечной силы (Д).
Кривая «длина-сила» соответствует сердечной диаграмме «давление-объем», где длине соответствует объем наполнения желудочка, а силе -давление в желудочке. Концентрация Са2+ в цитоплазме может влиять на соотношение давление/объем из-за изменения сократимости.
Другие важные функциональные различия между сердечной и скелетной мышцами перечислены ниже.
Поскольку скелетная мышца, по сравнению с сердечной, более эластична, пассивная сила растяжения сердечной мышцы в покое больше, чем таковая скелетной мышцы (Д1, 2)
В норме скелетная мышца функционирует в области плато кривой «длина-сила», тогда как сердечная мышца в норме работает на восходящем участке (ниже Lmax) кривой (которая не имеет плато) (В, Д1, 2). Следовательно, желудочек отвечает на увеличение диастолического наполнения увеличением развиваемой силы (механизм Франка-Старлинга). В сердечной мышце растяжение также воздействует на чувствительность тропонина к Са2+, что дает более крутую кривую (Д2).
Потенциал действия в сердечной мышце гораздо продолжительнее, чем в скелетной мышце, потому что gK< понижается со временем, а gCа увеличивается за 200-500 мс после быстрой инактивации Na+-каналов. Это вызывает медленный приток Са2+, в результате чего потенциал действия достигает плато. Таким образом, период рефрактерности не заканчивается почти до конца сокращения. Следовательно, тетанус в сердечной мышце невозможен.
В отличие от скелетной мышцы, в сердечной мышце нет двигательных единиц. Вместо этого стимул распространяется по всем волокнам предсердий, а потом желудочков, вызывая сокращение по принципу «все или ничего» обоих предсердий и желудочков.

Е. Мышечная сила (или нагрузка) и скорость укорачивания
В сердечной мышце, но не в скелетных, продолжительность потенциала действия может менять силу сокращения, которая контролируется варьированием притока Са2+ в клетку.
Чем больше сила (нагрузка), тем ниже скорость изотонического сокращения (см. диаграмму «скорость-сила», E1). Максимальная сила с небольшим количеством тепла - в отсутствие укорочения. Максимальная скорость (примерно 7 м/с в бицепсах) и много тепла - в отсутствие нагрузки. Сокращения с незначительной нагрузкой, таким образом, могут быть совершены гораздо быстрее, чем с тяжелой нагрузкой (Е2). Общее количество энергии, потребляемое для работы и теплообмена, больше при изотоническом сокращении, чем при изометрическом. Мощность мышцы - это произведение силы на скорость укорочения: Н • м • с-1 = Вт (Е1, закрашенные области диаграммы).
ЭНЕРГЕТИЧЕСКОЕ ОБЕСПЕЧЕНИЕ МЫШЕЧНОГО СОКРАЩЕНИЯ.
Непосредственным источником энергии для мышечной деятельности служит реакция расщепления АТФ. При ферментативном гидролизе АТФ происходит освобождение энергии, преобразуемой в процессе мышечного сокращения в механическую работу.
Запас АТФ в мышцах невелик. Для поддержания активности тканей на определенном уровне неоходим быстры ресинтез АТФ. Последний происходит в процессе рефосфорилирования при соединении АДФ и фосфатов. Наиболее доступным веществом является креатин-фосфат: КФ + АДФ = Креатин +АТФ
Концентрация КФ в мышцах в 2-4 раза больше в
Аденозинтрифосфат (АТФ) является нуклеотидом. В состав молекулы АТФ входят азотистое основание - аденин, углевод - рибоза и три остатка фосфорной кислоты (аденин, связанный с рибозой, называется аденозином).
Особенностью молекулы АТФ является то, что второй и третий остатки фосфорной кислоты присоединяются связью, богатой энергией. Такая связь называется высокоэнергетической, или макроэргической, и обозначается знаком Соединения, имеющие макроэргические связи, обозначаются термином «макроэрги».
Структурная формула АТФ имеет следующий вид:
В упрощенном виде строение АТФ можно отразить схемой: Аденин — рибоза.— Ф.К. ~ Ф.К. ~ Ф.К.
Y
аденозин
При использовании АТФ в качестве источника энергии обычно происходит отщепление путем гидролиза последнего остатка фосфорной кислоты:
АТФ + Н20 -> АДФ + Н3Р04 + Q (энергия)
В физиологических условиях, т. е. при тех условиях, которые имеются в живой клетке (температура, рН, осмотическое давление, концентрация реагирующих веществ и пр.), расщепление моля АТФ (506 г) сопровождается выделением 12 ккал, или 50 кДж* энергии.
Основным поставщиком АТФ является тканевое дыхание - завершающий этап катаболизма, протекающий в митохондриях всех клеток, кроме красных клеток крови (эритроцитов).
Содержание АТФ в мышце относительно постоянно. Концентрация АТФ составляет около 5 мм на 1 кг сырого веса мышц (около 25%). Накапливать большое количество АТФ мышца не может, т.к. при этом возникает субстратное угнетение миозиновой АТФ-азн, препятствующее образованию спаек между актиновыми и миозановыми нитями в миофибриллах и ведущее к утрачиванию сократительной способности мышцы. Вместе с тем концентрация АТФ в мышце не может снижаться ниже 2 мм на 1 кг сырого веса ткани, поскольку при этом перестанет действовать "кальциевый насос" в пузырьках, и мышца будет сокращаться вплоть до полного исчерпания всех запасов АТФ и развития тригора (состояния стойкого непроходящего сокращения).
Запасов АТФ в мышце обычно хватает на 3-4 одиночных сокращения максимальной силы. В то же время, как показывают исследования с использованием микробиопсии мышц, в процессе мышечной работы не наблюдается значительного снижения концентрации АТФ. Это объясняется тем, что по ходу мышечной деятельности АТФ восстанавливается из продуктов распада (ресинтезируется) с той же скоростью, с какой она расщепляется в процессе мышечных сокращений. Для обращения реакции гидролиза АТФ конечные продукты распада –АДФ и Н3РО4 – должны получить из реакционной среды энергию, равную той, что выделилась при распаде АТФ. Следовательно, синтез АТФ должен быть сопряжен с каталитическими реакциями, при которых освобождается значительное количество потенциальной химической энергии.
Ресинтез АТФ при мышечной деятельности может осуществляться
как в ходе реакции, идущих без кислорода, так и за счет окислительных превращений в клетках, связанных с потреблением кислорода. В обычных условиях ресинтез АТФ происходит в основном путем аэробных превращений, но при напряженной мышечной деятельности, когда доставка кислорода к мышцам затруднена, в тканях одновременно усиливаются анаэробные процессы ресинтеза АТФ.





