Генетика русских

В первой части статьи мы рассмотрели результаты многолетнего труда российских антропологов в деле изучения этнической истории русского народа. Во второй половине XX столетия на помощь физической антропологии пришла генетика, которая в современных условиях превращается в один из основных инструментов исследования этнических процессов. О генетике мы и поговорим.
Становление генетики как академической науки произошло в начале XX столетия, когда были повторно открыты и подтверждены основы учения австрийского монаха-августинца Георга Менделя о наследственности. В 1915 году американец Томас Морган создал хромосомную теорию наследственности, а несколькими годами ранее Вильгельм Иогансен впервые ввел в употребление термин «ген».
Генетика в России
В Российской Империи, подарившей миру двух нобелевских лауреатов по биологии — И.И. Мечникова и И.П. Павлова — молодая наука сразу обрела преданных поклонников. Накануне Первой мировой войны в Петербургском университете начинает читать курс лекцией по генетике Ю.А. Филипченко, а в 1917 году в Москве по инициативе выдающегося русского биолога Николая Константиновича Кольцова был открыт Институт экспериментальной биологии (ИЭБ). Тремя годами позже на базе Института появится Русское евгеническое общество, членом которого был уже упоминавшийся в первой части материала антрополог В.В. Бунак.
Кольцов и Бунак проводят исследования по частоте групп крови ABO у восточных славян, а институт становится центром притяжения для лучших умов отечественной биологии — Н.П. Дубинина, Н.В. Тимофеева-Ресовского, А.С. Серебровского и других. Именно Александр Сергеевич Серебровский в 1928 году впервые ввел в употребление термин «геногеография» — для обозначения дисциплины, изучающей генофонды отдельных человеческих популяций.

Тогда же вошло в обращение само понятие «генофонд» — для обозначения совокупности всех генов, распространенных в той или иной популяции. С этого момента геногеография становится одним из важнейших инструментов, помогающих в изучении процессов этногенеза ныне существующих народов. Сам Серебровский изначально считал генеографию прежде всего разделом исторической науки, с помощью которого ученый может увидеть в генофонде современных народов следы древних миграций, расового и этнического смешения.
Тяжелейший удар по отечественной генетике, нанесенный в период «лысенковщины», затормозил и развитие отечественной генеографии. В то время как в Британии выходит классическая монография Рональда Фишера «Генетическая теория естественного отбора», заложившая основы современной популяционной генетики, в СССР при полном одобрении Сталина начинаются «гонения на генетику», организованные псевдоученым Трофимом Лысенко. Самой известной жертвой этой кампании стал Николай Вавилов, умерший в тюрьме в 1943 году.
Господству Лысенко и его сторонников пришел конец после отставки Хрущева. В 1966 году появляется Институт общей генетики им. Н.И. Вавилова. Там в лаборатории генетики человека проводит свои исследования генетического полиморфизма коренных народов России Ю.Г. Рычков. В 1970-е годы огромный вклад в изучение генетического разнообразия человеческих популяций внесли также итальянский ученый Луиджи Кавалли-Сфорца и американец Ричард Левонтин. Настоящей же научной революцией стала расшифровка в 1953 году структуры ДНК Джеймсом Уотсоном, Френсисом Криком и Морисом Уилкинсоном — спустя десятилетие все трое были удостоены Нобелевской премии.

Это открытие позволило генетикам в перспективе перейти к изучению ДНК маркеров, о которых дальше пойдет речь.
Маркеры и гаплогруппы
Человек наследует ДНК обоих биологических родителей. Гены не наследуются целиком, происходит рекомбинация — фрагменты генов из поколения в поколение перемешиваются и замещают друг друга, образуя новые комбинации. Исключение составляют два участка ДНК, которые мы получаем от матери и отца соответственно — митохондриальная ДНК и не подвергающийся рекомбинации участок (около 95% длины) Y-хромосомы. В свою очередь митохондрии передаются потомкам обоего пола от матери вместе с яйцеклеткой, в то время как Y-хромосома «наследуется» от отца к сыну.
Изучение этих однородительских маркеров позволило генетикам заглянуть далеко в прошлое. Полученная нами в неизменном виде от матери митохондриальная ДНК (мтДНК) может привести нас к далекому предку по материнской линии, жившему тысячи и даже десятки тысяч лет назад. При этом мтДНК подвергается неизбежным точечным мутациям, которые, в свою очередь, наследуются. Это дает возможность увидеть небольшие отличия в генетическом коде представителей разных популяций и определить, когда они возникли. Митохондриальная ДНК была открыта в 1963 году, а в 1987 году коллектив ученых под руководством Ребекки Канн опубликовал в журнале Nature статью о «митохондриальной Еве» — последнем общем для всех людей женском предке, вернее, группе предков.
По первоначальной версии Канн «митохондриальная Ева» могла жить в Восточной Африке около 150–250 тысяч лет назад. Позже эти даты скорректировали — в пределах 100–150 тысяч лет назад. Общее число Homo sapiens было тогда очень невелико — несколько десятков тысяч человек. В дальнейшем пути разных человеческих групп разошлись, а порядка 70–100 тысяч лет назад анатомически современный человек покинул пределы Африки через Баб-эль-Мандебский пролив или Синайский полуостров.

Миграции Homo sapiens.
Изучение накопившихся в мтДНК современных людей мутаций позволило в свою очередь определить, как и какими путями расселялись те или иные «дочери Евы» по планете. Изучали и мутации Y-хромосомы. Эти исследования позволили в 1990-е выделить отдельные гаплогруппы как по мтДНК, так и по У-ДНК, и составить их древо.
Гаплогруппы, как группы схожих гаплотипов, выделялись на основе обнаружения нескольких общих мутаций. Каждой гаплогруппе присвоили буквенное обозначение — L, M, T и так далее. Самые древние — A (У-ДНК) и L (мтДНК) — первоначально возникли на территории Африки — прародине Homo sapiens. Как выяснилось, у каждой гаплогруппы есть определенная географическая привязка. Так, отдельные гаплогруппы распространены прежде всего на территории Восточной Азии, среди монголоидных народов. Другие — преимущественно в Европе, на Ближнем Востоке и в Северной Африке среди европеоидов.
Разумеется, связь между гаплогруппами и расовыми группами не может быть абсолютной. Но теперь изучение миграций носителей тех или иных гаплогрупп позволило генетикам больше узнать о процессах расового и этнического смешения. В особенности это пригодилось при исследовании крайне неоднородного населения молодых государств Южной и Северной Америки. Для примера — изучение однородительских маркеров у населения США позволило ученым сделать вывод о наличии заметной западноевразийской (европейской) примеси у афроамериканцев и практически полном отсутствии субсахарской (африканской) примеси у белых американцев.



Генетика русских
Перейдем к русским и другим народам Восточной Европы. Первые актуальные с методологической точки зрения исследования однородительских маркеров в русских популяциях появляются в конце 1990-х. Помимо довольно узких научных проблем популяционные генетики пытались получить ответы и на некоторые вопросы, интересные обывателю. Это, во-первых, определение степени генетической близости русских и соседних народов, в особенности восточнославянских, а во-вторых, роль восточноевразийского (читай — монголоидного) компонента в сложении русского генофонда. То самое пресловутое наследие «трехсотлетнего монгольского ига».
За последний десяток лет самый большой вклад в изучение генетической изменчивости русских популяций в «историческом ареале» внесли Е.В. Балановская, и ее сын О.П. Балановский (оба — доктора биологических наук). Исследования генофонда русского населения Сибири и Дальнего Востока проводятся на базе Института биологических проблем севера (руководитель лаборатории генетики — Б.А. Малярчук). Исследователи обычно стараются изучать население небольших городов и деревень. У самих обследованных все бабушки и дедушки должны принадлежать к одному этносу и региональной популяции. В других случаях может изучаться население крупных городов, а требования к происхождению выдвигаются менее строгие.
Стоит оговориться: популяционные генетики в своей работе изучают не народы, а отдельные популяции. У генов нет ни языка, ни этнической идентичности. Задача ученых — определить, в какой степени генетические границы совпадают с этническими и какой набор генов характерен для генофонда тех или иных народов. Нет русских, украинских или татарских генов, есть гены в большей или меньшей степени характерные для тех или иных популяций. И есть генофонд отдельных народов как совокупность генов на определенной территории.
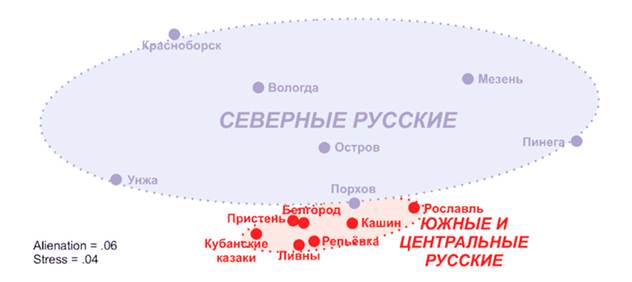
Первое, о чем стоит сказать — это внутренние различия между отдельными группами русских. На сегодняшний день изучено несколько десятков популяций на огромной территории от Смоленска до Владивостока. Наиболее полно обследованы русские в своем «историческом ареале» — в границах российского государства эпохи Ивана Грозного. В классической монографии «Русский генофонд на русской равнине» Балановские отметили гетерогенность (разнородность) русского генофонда по данным однородительских маркеров. При этом большинство центральных и южных русских популяций образуют единый кластер, демонстрируя близость между собой. Исключение составляют северные популяции (выборки из Архангельской и Вологодской областей), заметно отличающиеся как от центральных и южных групп, так и друг от друга. Именно это своеобразие северных русских — причина большей по сравнению с другими европейскими народами генетической гетерогенности русских в целом.
Соседи
Говоря о женских однородительских маркерах (мтДНК), О.П. Балановский очертил круг самых близких популяций:
По степени сходства с русскими генофондом его соседи располагаются в следующем порядке: восточные славяне, волго-финские народы, западно-финские народы, западные славяне.
В статье коллектива российских, эстонских и британских специалистов исследован полиморфизм Y-хромосомы (отцовские линии) в 14 русских популяциях (1228 мужчин) — от кубанских казаков на юге до жителей Мезени и Пинеги на севере. Результаты работы генетиков подтвердили вывод о значительных отличиях северных великорусов — жителей Вологодской и Архангельской областей — от остальных русских популяций. В южных и центральных группах самой распространенной оказалась характерная для других восточных славян и поляков гаплогруппа R1a, обнаруженная в 55,4% и 46,5% случаев соответственно. На севере же R1a выявили лишь у 34,2% обследованных, первенство досталось гаплогруппе N3 (по новой номенклатуре N1C), типичной для прибалтийско-финских и балтских популяций.

Карта генетических расстояний между европейскими народами (Y-хромосома). Русские: R1 — Мезень, R2 — Пинега, R3 — Красноборск, R4 — Вологда, R5-12 — южные и центральные русские. Источник: Two Sources of the Russian Patrilineal Heritage in Their Eurasian Context
В результате южные и центральные русские популяции образовывают компактный кластер с украинскими, белорусскими и польскими группами.
Северные великорусы на картах генетических расстояний оказались заметно смещены в сторону североевропейских популяций — финной и шведов. Своеобразие жителей Архангельской и Вологодской областей не могло оставить генетиков равнодушным. В статье «Генофонд Русского Севера: Славяне? Финны? Палеоевропейцы?» коллектив российских и европейских ученых попытался определить место северных русских на генетической карте Европы. Были изучены данные по однородительским маркерам и распространенность СПИД-протекторной мутации del32 в аутосомном гене CCR5. Обнаружилась близость упомянутых групп с целым рядом североевропейских народов — литовцами, латышами, шведами и прибалтийскими финнами.
Сходство может указывать на общий для всех северных европейцев генетический пласт, восходящий корнями к охотникам и собирателям эпохи мезолита. Это в свою очередь может свидетельствовать о роли финно-угорского субстрата, а также о заметных различиях между ильменскими словенами, потомками которых являются русские Вологодской и Архангельской областей, и другими средневековыми восточнославянскими племенами.
Нужно упомянуть и такую своеобразную группу русского служилого населения как казачество. Живя на окраинах русского государства, эти представители военно-служилого сословия неизбежно вступали в контакты, в том числе и брачные, с соседними народами. В последние годы генетики из Москвы, Харькова, Ростова-на-Дону и Майкопа провели исследования кубанских, донских и терских казаков.
Анализ показал своеобразие каждой из трех групп. Генофонд кубанских казаков оказался «слепком с русского», не несущим никаких следов неславянского субстрата. У терцев же наблюдается увеличение кавказского генетического компонента. Результаты прекрасно совпадают со сведениями исторических источников. Если кубанское казачье войско формировалось на основе донских казаков (линейцев) и запорожцев, то в состав терских казаков помимо русских записывали чеченцев, осетин и других кавказцев. Сами донские казаки вполне ожидаемо оказались неотличимы от южнорусских популяций и близки к украинцам.

Особый интерес представляет, разумеется, сравнение русских с другими восточнославянскими народами. Генофонд украинцев и белорусов был подробно изучен как российскими, так и местными генетиками. Кандидатская диссертация Андрея Пшеничнова рассматривает структуру генофонда украинцев по данным полиморфизма как мтДНК, так и Y-хромосомы. Все украинские популяции вошли в компактный кластер вместе с кубанскими казаками, юго-западными русскими, белорусами и поляками. В 2013 году коллектив белорусских, эстонских, израильских и итальянских генетиков изучил 6 региональных групп белорусов. На уровне внутриэтнических различий обнаружились различия между северными и южными белорусами. Белорусы оказались очень близки к соседним славянским народам — русским, украинцам и полякам. Вопреки стереотипам о балтском происхождении, генетическое сходство с литовцами и латышами как по «мужским», так и по «женским» линиям оказалось незначительным.

Аутосомные маркеры
В последние годы наряду с изучением полиморфизма мтДНК и Y-хромосомы становятся все более востребованы данные по аутосомным маркерам. Речь идет о генетических маркерах, расположенных на аутосомах — неполовых хромосомах. Ученых интересуют так называемые точки генетического разнообразия (однонуклеотидный полиморфизм или SNP — снип).
Изучение десятков, а в последние годы и сотен тысяч подобных снипов дает определенную информацию о геноме конкретного человека. В то время как однородительские маркеры позволяют судить о предках лишь по одной мужской и одной женской линии, изучение однонуклеотидных полиморфизмов позволяет получить своеобразный генетический паспорт. Эта особенность активно используется в персональном генетическом тестировании коммерческими организациями — для определения отдаленного родства, принадлежности к определенным популяциям, склонности к тем или иным заболеваниям. К недостаткам можно отнести малый объем выборок — проблема, безусловно, временная. Сейчас число изученных русских популяций сравнительно невелико, но полученный массив информации уже позволяет сделать определенные выводы.
В работе, посвященной генетическому анализу популяций Европейской части России, исследованы четыре выборки этнических русских (из Твери, Курска, Мурома и Мезени). Результаты оказались схожи с уже полученными ранее путем изучения полиморфизма мтДНК и Y-хромосомы. Жители Твери, Курска и Мурома оказались в одном кластере с поляками, чехами и немцами. Северные русские вполне ожидаемо сместились в сторону вепсов и коми-зырян. Весьма примечательно, что результаты, полученные при использовании всех трех генетических систем, совпадают с данными антропологии. Именно в северной вологдо-вятской зоне антропологи обнаружили заметный финно-угорский субстрат.

Изученные популяции
Сопоставление трех генетических систем позволило сделать ряд общих выводов:
1) Восточные славяне (русские, украинцы и белорусы) образуют свой кластер, при этом не «перекрываясь» полностью друг с другом. Исключение составили северные русские, тяготеющие к финно-угорским популяциям. Наиболее близки к восточным славянам поляки, в то время как чехи смещены в сторону немцев.
2) Вновь подтверждено сходство балтских народов (латышей и литовцев) с эстонцами.

Генетическая структура балто-славянских популяций в сравнении с другими народами Европы по трем генетическим системам: А) по аутосомным SNP-маркерам, В) по Y-хромосоме, С) по митохондриальной ДНК.
3) Использование метода Admixture показало наличие в генофонде восточных европейцев трех основных предковых компонентов. Первый, однозначно преобладающий у восточных славян и балтов, видимо, связан с европейскими охотниками-собирателями. Второй восходит к эпохе неолитической революции и появился в Европе вместе с земледельцами из Северной Африки и с Ближнего Востока. Третий указывает на связь с Сибирью и ее монголоидными популяциями. Он практически отсутствует у балтов и славян, и более заметен у северных русских.

Генетика русских
В первой части статьи мы рассмотрели результаты многолетнего труда российских антропологов в деле изучения этнической истории русского народа. Во второй половине XX столетия на помощь физической антропологии пришла генетика, которая в современных условиях превращается в один из основных инструментов исследования этнических процессов. О генетике мы и поговорим.
Становление генетики как академической науки произошло в начале XX столетия, когда были повторно открыты и подтверждены основы учения австрийского монаха-августинца Георга Менделя о наследственности. В 1915 году американец Томас Морган создал хромосомную теорию наследственности, а несколькими годами ранее Вильгельм Иогансен впервые ввел в употребление термин «ген».
Генетика в России
В Российской Империи, подарившей миру двух нобелевских лауреатов по биологии — И.И. Мечникова и И.П. Павлова — молодая наука сразу обрела преданных поклонников. Накануне Первой мировой войны в Петербургском университете начинает читать курс лекцией по генетике Ю.А. Филипченко, а в 1917 году в Москве по инициативе выдающегося русского биолога Николая Константиновича Кольцова был открыт Институт экспериментальной биологии (ИЭБ). Тремя годами позже на базе Института появится Русское евгеническое общество, членом которого был уже упоминавшийся в первой части материала антрополог В.В. Бунак.
Кольцов и Бунак проводят исследования по частоте групп крови ABO у восточных славян, а институт становится центром притяжения для лучших умов отечественной биологии — Н.П. Дубинина, Н.В. Тимофеева-Ресовского, А.С. Серебровского и других. Именно Александр Сергеевич Серебровский в 1928 году впервые ввел в употребление термин «геногеография» — для обозначения дисциплины, изучающей генофонды отдельных человеческих популяций.
Тогда же вошло в обращение само понятие «генофонд» — для обозначения совокупности всех генов, распространенных в той или иной популяции. С этого момента геногеография становится одним из важнейших инструментов, помогающих в изучении процессов этногенеза ныне существующих народов. Сам Серебровский изначально считал генеографию прежде всего разделом исторической науки, с помощью которого ученый может увидеть в генофонде современных народов следы древних миграций, расового и этнического смешения.
Тяжелейший удар по отечественной генетике, нанесенный в период «лысенковщины», затормозил и развитие отечественной генеографии. В то время как в Британии выходит классическая монография Рональда Фишера «Генетическая теория естественного отбора», заложившая основы современной популяционной генетики, в СССР при полном одобрении Сталина начинаются «гонения на генетику», организованные псевдоученым Трофимом Лысенко. Самой известной жертвой этой кампании стал Николай Вавилов, умерший в тюрьме в 1943 году.
Господству Лысенко и его сторонников пришел конец после отставки Хрущева. В 1966 году появляется Институт общей генетики им. Н.И. Вавилова. Там в лаборатории генетики человека проводит свои исследования генетического полиморфизма коренных народов России Ю.Г. Рычков. В 1970-е годы огромный вклад в изучение генетического разнообразия человеческих популяций внесли также итальянский ученый Луиджи Кавалли-Сфорца и американец Ричард Левонтин. Настоящей же научной революцией стала расшифровка в 1953 году структуры ДНК Джеймсом Уотсоном, Френсисом Криком и Морисом Уилкинсоном — спустя десятилетие все трое были удостоены Нобелевской премии.
Это открытие позволило генетикам в перспективе перейти к изучению ДНК маркеров, о которых дальше пойдет речь.
Маркеры и гаплогруппы
Человек наследует ДНК обоих биологических родителей. Гены не наследуются целиком, происходит рекомбинация — фрагменты генов из поколения в поколение перемешиваются и замещают друг друга, образуя новые комбинации. Исключение составляют два участка ДНК, которые мы получаем от матери и отца соответственно — митохондриальная ДНК и не подвергающийся рекомбинации участок (около 95% длины) Y-хромосомы. В свою очередь митохондрии передаются потомкам обоего пола от матери вместе с яйцеклеткой, в то время как Y-хромосома «наследуется» от отца к сыну.
Изучение этих однородительских маркеров позволило генетикам заглянуть далеко в прошлое. Полученная нами в неизменном виде от матери митохондриальная ДНК (мтДНК) может привести нас к далекому предку по материнской линии, жившему тысячи и даже десятки тысяч лет назад. При этом мтДНК подвергается неизбежным точечным мутациям, которые, в свою очередь, наследуются. Это дает возможность увидеть небольшие отличия в генетическом коде представителей разных популяций и определить, когда они возникли. Митохондриальная ДНК была открыта в 1963 году, а в 1987 году коллектив ученых под руководством Ребекки Канн опубликовал в журнале Nature статью о «митохондриальной Еве» — последнем общем для всех людей женском предке, вернее, группе предков.
По первоначальной версии Канн «митохондриальная Ева» могла жить в Восточной Африке около 150–250 тысяч лет назад. Позже эти даты скорректировали — в пределах 100–150 тысяч лет назад. Общее число Homo sapiens было тогда очень невелико — несколько десятков тысяч человек. В дальнейшем пути разных человеческих групп разошлись, а порядка 70–100 тысяч лет назад анатомически современный человек покинул пределы Африки через Баб-эль-Мандебский пролив или Синайский полуостров.
Миграции Homo sapiens.
Изучение накопившихся в мтДНК современных людей мутаций позволило в свою очередь определить, как и какими путями расселялись те или иные «дочери Евы» по планете. Изучали и мутации Y-хромосомы. Эти исследования позволили в 1990-е выделить отдельные гаплогруппы как по мтДНК, так и по У-ДНК, и составить их древо.
Гаплогруппы, как группы схожих гаплотипов, выделялись на основе обнаружения нескольких общих мутаций. Каждой гаплогруппе присвоили буквенное обозначение — L, M, T и так далее. Самые древние — A (У-ДНК) и L (мтДНК) — первоначально возникли на территории Африки — прародине Homo sapiens. Как выяснилось, у каждой гаплогруппы есть определенная географическая привязка. Так, отдельные гаплогруппы распространены прежде всего на территории Восточной Азии, среди монголоидных народов. Другие — преимущественно в Европе, на Ближнем Востоке и в Северной Африке среди европеоидов.
Разумеется, связь между гаплогруппами и расовыми группами не может быть абсолютной. Но теперь изучение миграций носителей тех или иных гаплогрупп позволило генетикам больше узнать о процессах расового и этнического смешения. В особенности это пригодилось при исследовании крайне неоднородного населения молодых государств Южной и Северной Америки. Для примера — изучение однородительских маркеров у населения США позволило ученым сделать вывод о наличии заметной западноевразийской (европейской) примеси у афроамериканцев и практически полном отсутствии субсахарской (африканской) примеси у белых американцев.
Карты распространения гаплогрупп (мтДНК и Y-ДНК) и их возраст. Данные FamilyTreeDNA.
Генетика русских
Перейдем к русским и другим народам Восточной Европы. Первые актуальные с методологической точки зрения исследования однородительских маркеров в русских популяциях появляются в конце 1990-х. Помимо довольно узких научных проблем популяционные генетики пытались получить ответы и на некоторые вопросы, интересные обывателю. Это, во-первых, определение степени генетической близости русских и соседних народов, в особенности восточнославянских, а во-вторых, роль восточноевразийского (читай — монголоидного) компонента в сложении русского генофонда. То самое пресловутое наследие «трехсотлетнего монгольского ига».
За последний десяток лет самый большой вклад в изучение генетической изменчивости русских популяций в «историческом ареале» внесли Е.В. Балановская, и ее сын О.П. Балановский (оба — доктора биологических наук). Исследования генофонда русского населения Сибири и Дальнего Востока проводятся на базе Института биологических проблем севера (руководитель лаборатории генетики — Б.А. Малярчук). Исследователи обычно стараются изучать население небольших городов и деревень. У самих обследованных все бабушки и дедушки должны принадлежать к одному этносу и региональной популяции. В других случаях может изучаться население крупных городов, а требования к происхождению выдвигаются менее строгие.
Стоит оговориться: популяционные генетики в своей работе изучают не народы, а отдельные популяции. У генов нет ни языка, ни этнической идентичности. Задача ученых — определить, в какой степени генетические границы совпадают с этническими и какой набор генов характерен для генофонда тех или иных народов. Нет русских, украинских или татарских генов, есть гены в большей или меньшей степени характерные для тех или иных популяций. И есть генофонд отдельных народов как совокупность генов на определенной территории.
Первое, о чем стоит сказать — это внутренние различия между отдельными группами русских. На сегодняшний день изучено несколько десятков популяций на огромной территории от Смоленска до Владивостока. Наиболее полно обследованы русские в своем «историческом ареале» — в границах российского государства эпохи Ивана Грозного. В классической монографии «Русский генофонд на русской равнине» Балановские отметили гетерогенность (разнородность) русского генофонда по данным однородительских маркеров. При этом большинство центральных и южных русских популяций образуют единый кластер, демонстрируя близость между собой. Исключение составляют северные популяции (выборки из Архангельской и Вологодской областей), заметно отличающиеся как от центральных и южных групп, так и друг от друга. Именно это своеобразие северных русских — причина большей по сравнению с другими европейскими народами генетической гетерогенности русских в целом.





