Проводящие пути ЦНС построены из функционально однородных групп нервных волокон; они представляют собой внутренние связи между ядрами и корковыми центрами, расположенными в разных частях и отделах мозга, и служат для их функционального объединения. Проводящие пути, как правило, проходят в белом веществе спинного и головного мозга, но могут локализоваться и в покрышке ствола мозга, где четких границ между белым и серым веществом нет.
Основным проводящим звеном в системе передачи информации от одних центров мозга к другим являются нервные волокна - аксоны нейронов, передающие информацию в форме нервного импульса в строго определенном направлении, а именно от тела клетки. Среди проводящих путей в зависимости от их строения и функционального значения выделяют различные группы нервных волокон: волокна, пучки, тракты, лучистости, спайки (комиссуры).
Среди внутренних связей в ЦНС различают три группы проводящих путей: ассоциативные, комиссуралъные и проекционные.
Ассоциативные пути достаточно многочисленны и широко представлены в различных отделах ЦНС, но наиболее развиты они в коре мозга. Эти пути образованы ассоциативными нейронами и их волокнами, соединяющими различные области коры мозга одного и того же полушария большого мозга или мозжечка. Ассоциативные пути соединяют также различные участки одной и той же половины (правой или левой) спинного мозга
Ассоциативные волокна в полушарии мозга могут быть короткими и длинными (рис. 28). Короткие волокна соединяют между собой соседние извилины. Длинные волокна соединяют между собой отдаленные друг от друга участки коры или доли большого мозга.
Среди ассоциативных пучков больших полушарий выделяют верхний продольный пучок, соединяющий лобную долю с нижней теменной долькой, затылочной долей и задней частью височной доли. Нижний продольный пучок соединяет затылочную и височную доли, а крючковидный пучок - нижнюю поверхность лобной доли и височный полюс.
Рис. 28. Ассоциативные связи между долями и извилинами в левом полушарии мозга.
В коре мозжечка расположенные рядом листки и дольки как в черве, так и в полушариях, соединены между собой ассоциативными волокнами. К ассоциативным относят также связи, образующиеся между ядрами одной половины ствола мозга, промежуточного мозга и базальными ядрами соответствующего полушария.
В спинном мозге ассоциативные нейроны обеспечивают также межсегментарные связи спинного мозга. Волокна могут быть короткими и соединять между собой соседние спинномозговые сегменты, длинными связывать отдаленные сегменты.
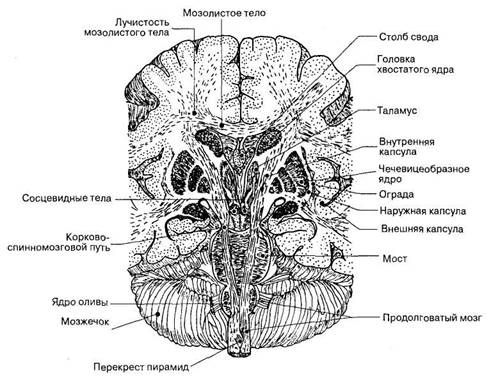
Комиссуральные пути, или спайки, состоят из нейронов и их волокон, обеспечивающих связи между зеркально симметричными участками правой и левой половин головного и спинного мозга. Наиболее мощной спайкой конечного мозга, состоящей из комиссуральных волокон, соединяющих правое и левое полушария, является мозолистое тело (рис. 29).
Менее крупные мозговые спайки - передняя комиссура и комиссура свода - соединяют участки старой и древней коры полушарий (парагиппокампальные извилины, гиппокампы, зубчатые извилины и др.). В отличие от большого мозга существование комиссуральных путей между правым и левым полушариями мозжечка в настоящее время остается недоказанным.
Рис. 29. Комиссуральные волокна и проекционные волокна.
В области спинного мозга и в стволе головного мозга истинные комиссуральные пути отсутствуют. Однако подобием мозговых спаек являются перекресты - части восходящих или нисходящих проводящих путей ЦНС, в которых происходит переход волокон с одной половины мозга на противоположную половину. Например, в передней белой спайке спинного мозга перекрещиваются восходящие волокна правого и левого латеральных спино-таламических трактов, а также нисходящие волокна переднего корково-спинномозгового пути.
Проекционные пути состоят из нейронов и их волокон, обеспечивающих связи между спинным и головным мозгом. Проекционные пути соединяют также ядра ствола с базальными ядрами и корой больших полушарий, а также ядра ствола с корой и ядрами мозжечка. Проекционные пути могут быть восходящими и нисходящими.
Восходящие (сенсорные, чувствительные или афферентные) проекционные пути проводят нервные импульсы от экстеро-, проприо- и интерорецепторов, а также от органов чувств в восходящем направлении к головному мозгу, преимущественно к коре мозга. Помимо коры головного мозга сенсорная информация направляется и в другие отделы нервной системы, а именно в мозжечок, в средний мозг, в ретикулярную формацию.
Сенсорная (чувствительная) информация играет очень важную роль в жизнедеятельности человека. Она поступает в нервную систему различными путями. Через кожный покров и от органов чувств идет поток внешней (экстероцептивной) информации, сигнализирующий о состоянии внешней
Рис. 30. Общий план строения анализатора.
среды (рис. 30)
Сенсорная информация от туловища и конечностей по чувствительным волокнам спинномозговых нервов поступает к спинному мозгу, от которого по восходящим путям направляется в головной мозг. При этом восходящие проекционные связи спинного мозга с головным начинаются либо вне спинного мозга от нейронов спинномозговых узлов, либо от нейронов, расположенных в задних столбах спинного мозга. Сенсорная информация от органов головы и частично шеи поступает непосредственно в головной мозг по чувствительным волокнам черепных нервов; при этом восходящие проекционные волокна начинаются в их сенсорных ядрах.
Общей особенностью сенсорных путей является многоэтапная передача возбуждения через различные ядерные центры, в которых происходит последовательный анализ информации. В стволе мозга сенсорные проводящие пути располагаются в его покрышке и, направляясь к коре мозга, обязательно проходят через промежуточный мозг, через его зрительные бугры (таламусы), в ядрах которых залегают подкорковые центры всех видов чувствительности, кроме слуховой. В них происходит переключение сенсорных путей; при этом сенсорная информация проходит частичную обработку (анализ и синтез) перед тем, как будет направлена к коре больших полушарий.
Среди нисходящих проводящих путей существенное значение имеют пути экстрапирамидной системы, а также ретикулоспинальные пути. Следует отметить, что подразделение нисходящих проводящих путей, участвующих в регуляции и координации движений, на пирамидную и экстрапирамидную системы в значительной мере условно и далеко не полностью отражает сложность иерархических отношений между корковыми и многочисленными подкорковыми моторными центрами.
Пирамидная система представляет собой совокупность двигательных центров коры мозга, моторных центров черепных нервов, залегающих в стволе мозга, и моторных центров в передних рогах спинного мозга, а также эфферентных проекционных нервных волокон, связывающих их между собой. Пирамидные пути обеспечивают проведение импульсов в процессе сознательной регуляции движений (рис. 31).
Пирамидные пути (примерно 40% волокон) начинается в предцентральной извилине, где находится корковый центр двигательного анализатора;около 20% волокон - от постцентральной извилины, а остальные 40% волокон - от задних участков лобных извилин, и от надкраевой извилины нижней теменной дольки, в которой расположен центр праксии, контролирующий сложные координированные целенаправленные движения.
Пирамидные пути подразделяют на корково-спинномозговой и корково-ядерный. Их общей особенностью является то, что они, начинаясь в коре правого и левого полушарий, переходят на противоположную сторону мозга (т. е. перекрещиваются) и в конечном итоге осуществляют регуляцию движений контрлатеральной половины тела.
Корково-спинномозговой путь начинается от новой коры больших полушарий. Его волокна продолжаются в основании среднего мозга и основании моста. В продолговатом мозге на уровне пирамид волокна медиальных частей правого и левого корково-спинномозговых путей переходят на противоположную сторону, образуя перекрест пирамид. Здесь перекрещивается около 80% волокон пирамидных трактов. Перекрещенные волокна спускаются в спинной мозг и идут в составе боковых канатиков. Волокна пирамидных трактов заканчиваются синапсами на мотонейронах двигательных ядер передних рогов спинного мозга. Неперекрещенная часть волокон продолжается в спинной мозг в составе его передних канатиков под названием переднего корково-спинномозгового тракта. Волокна этих трактов переходят на противоположную сторону на уровне соответствующих сегментов спинного мозга и заканчиваются синапсами преимущественно на интернейронах, которые передают центральные нервные влияния на мотонейроны двигательных ядер передних рогов спинного мозга. Только небольшая часть пирамидных волокон (около 8%) непосредственно оканчивается на мотонейронах спинного мозга. Бoльшая часть волокон пирамидных путей оканчивается на интернейронах спинного мозга, а уже от них раздражение передается на мотонейроны.
Нервные волокна, образующие пирамидные пути, неоднородны как по своим структурным, так и функциональным параметрам. Большую часть составляют тонкие и безмиелиновые нервные волокна, а крупные миелиновые волокна, отличающиеся высокой скоростью проведения нервного импульса, составляют значительно меньшую часть. С этими структурными особенностями связано наличие быстро- и медленнопроводящих частей пирамидной системы.
Рис. 31. Проводящий путь пирамидной системы.
Полагают, что по пирамидным путям происходит активация преимущественно мускулатуры сгибателей и торможение разгибателей. Возможно, что одно из функциональных значений пирамидной системы состоит в тонизирующем воздействии корковых центров на работу мотонейронов спинного мозга.
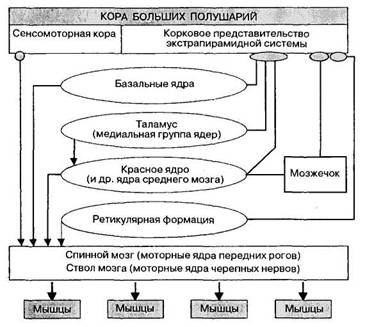
Экстрапирамидная система объединяет филогенетически более древние механизмы управления движениями человека, чем пирамидная система. Она осуществляет преимущественно непроизвольную, автоматическую регуляцию сложных двигательных актов, таких как ходьба, бег, поддержание позы, жевание и т.п.; она регулирует тонус мышц, а также участвует в регуляции двигательных проявлений эмоций. Отличительной особенностью экстрапирамидной системы является многоэтапная, с множеством переключений, передача нервных влияний от различных отделов головного мозга к исполнительным центрам - моторным ядрам спинного мозга и черепных нервов (рис. 32).
По экстрапирамидным путям происходит передача двигательных команд при защитных двигательных рефлексах, протекающих бессознательно. Например, благодаря экстрапирамидным путям передается информация при восстановлении вертикального положения тела в результате потери равновесия (вестибулярные рефлексы) или при двигательных реакциях на внезапное световое или звуковое воздействие (защитные рефлексы) и т. д.
Экстрапирамидную систему образуют ядерные центры полушарий (базалъные ядра: хвостатое чечевицеобразное), промежуточного мозга и ствола мозга (красное ядро, черное вещество), а также проводящие пути, связывающие ее с корой больших полушарий, с мозжечком, с ретикулярной формацией и, наконец, с исполнительными центрами, лежащими в моторных ядрах черепных нервов и в передних рогах спинного мозга.
Рис. 32. Схема пирамидных и экстрапирамидных связей в системе управления движениями человека.
Существует также и несколько расширенная трактовка состава экстрапирамидной системы, когда к ней причисляют мозжечок, ядра четверохолмия среднего мозга, ядра ретикулярной формации и т. п.
Корковые пути берут начало от предцентральной извилины, а также других отделов коры мозга; эти пути проецируют влияние коры на базальные ядра. Сами базальные ядра (хвостатое и чечевицеобразное) тесно связаны многочисленными внутренними связями между собой, а также с ядрами таламуса и с красным ядром среднего мозга. Формирующиеся здесь двигательные команды передаются на исполнительные двигательные центры спинного мозга преимущественно двумя путями: через красноядерно-спинномозговой тракт и через ядра ретикулярной формации (ретикулоспинальный тракт). Надо также отметить, что через красное ядро осуществляется передача влияний мозжечка на работу спинномозговых моторных центров.
Красноядерно-спионномозговой тракт образован отростками нейронов, расположенных в красном ядре. Само красное ядро получает проекции от базальных ядер, субталамического ядра, зубчатого ядра мозжечка и других подкорковых структур головного мозга. Аксоны нейронов красного ядра образуют перекрест на уровне среднего мозга в его покрышке и далее, спускаясь в покрышке ствола, достигают спинного мозга. Здесь волокна красноядерно-спинномозгового тракта продолжаются в боковых канатиках спинного мозга. Заканчиваются волокна красноядерно-спинномозгового тракта синапсами на мотонейронах в двигательных ядрах спинного мозга.
На мотонейронах спинного мозга заканчиваются синапсами также волокна ретикулоспиналъного тракта, передающего влияния экстрапирамидной системы. С помощью этих трактов поддерживается активность спинномозговых двигательных центров и обеспечивается вестибулярный контроль в процессе регуляции движений. Особенностью этих трактов является отсутствие перекрестов их волокон, поскольку каждый из них обеспечивает связи правой и левой половин ствола головного мозга только со своей половиной спинного мозга.
Автономная нервная система составляет ту часть нервной системы, которая регулирует висцеральные функции организма. К ним относятся кровообращение, дыхание, пищеварение, обмен веществ, т. е. обеспечение трофических функций организма и поддержание состава внутренней среды.
Автономная нервная система регулирует функции внутренних органов не связанные непосредственно с передвижением в пространстве. Автономную нервную систему называют еще вегетативной нервной системой в отличие от соматической, которая контролирует работу опорно-двигательного аппарата и устанавливает связь с окружающей средой через покровы тела и органы чувств.
В автономной нервной системе различают центральный и периферический отделы. К центральному отделу относятся вегетативные ядра, расположенные в головном и спинном мозге, к периферическому - вегетативные узлы, вегетативные нервы, сплетения и внутриорганные сплетения (рис. 33).
Наряду с вегетативными нейронами, расположенными в ядрах ствола мозга и спинного мозга, в головном мозге имеются еще и другие вегетативные нервные центры, занимающие более высокое иерархическое положение. Это многочисленные ядра в области гипоталамуса, оказывающие нисходящие регуляторные влияния на стволовые и спинномозговые вегетативные центры или через гипофиз и другие эндокринные железы - непосредственно на работу внутренних органов. Высшие центры, управляющие вегетативными функциями, находятся в коре мозга, а именно в лобной, теменной и височной областях. Здесь происходит синтез вегетативных и анимальных функций всего организма человека, а также их согласование с его психоэмоциональной деятельностью.
Управляя работой внутренних органов и регулируя обмен веществ, автономная нервная система оказывает на них разное, подчас противоположное действие. Под ее влиянием возможно как сужение, так и
Рис. 33. Очаги локализации вегетативных ядер и мест выхода вегетативных нервов из центральной нервной системы (римскими цифрами обозначены пары черепных нервов).
расширение просвета кровеносных сосудов, повышение или снижение частоты сокращений сердца и т. д. По характеру влияния на иннервируемые органы автономная нервная система подразделяется на симпатическуюи парасимпатическую части.
Функциональные проявления жизнедеятельности внутренних органов под влиянием симпатической нервной системы характерны для состояний повышенной активности организма (физического и психического напряжения), а парасимпатической - для его относительного покоя. Так, например, раздражение симпатических нервных элементов вызывает повышение частоты сокращений сердца, сужение кровеносных сосудов и повышение артериального давления, учащение дыхательных движений, ослабление перистальтики кишечника, расширение зрачков.
Возбуждение парасимпатических нервных элементов, напротив, приводит к снижению частоты сокращений сердца, понижению артериального давления и т. д. Однако эти функциональные различия нельзя рассматривать как прямой антагонизм. Обе части автономной нервной системы работают согласованно, функционально дополняя друг друга.
К симпатической части автономной нервной системе относится симпатический ствол с его узлами и отходящие от них вегетативные нервы и ветви.
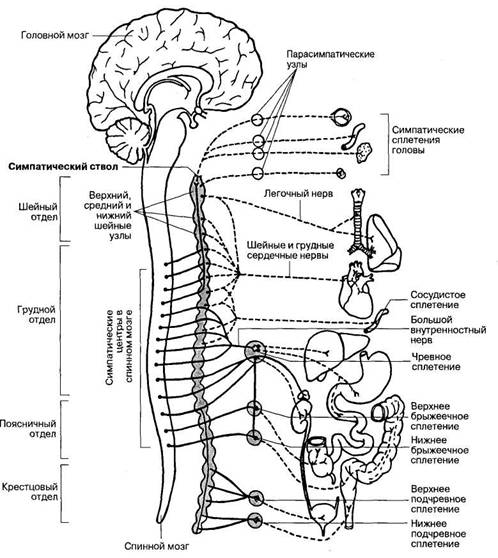
Симпатический ствол парное образование; он состоит из цепочки нервных узлов, располагающихся по бокам от позвоночного столба на всем его протяжении (рис. 34). Все узлы симпатического ствола связаны между собой межузловыми ветвями. В симпатическом стволе выделяют: шейный, грудной, поясничный, крестцовый и копчиковый отделы, представленные вегетативными узлами.
В симпатической части автономной нервной системы преганглионарные нейроны отходят от спинного мозга только в его грудном и частично поясничном отделах. Тела преганглионарных нейронов залегают в ядрах боковых рогов спинного мозга. Эти ядра прослеживаются на уровне всех грудных и верхних двух поясничных сегментов спинного мозга.
Преганглионарные симпатические волокна выходят из спинного мозга в составе передних корешков спинномозговых нервов. Их аксоны покрыты миелиновой оболочкой и в силу этого образуют белые соединительные ветви, отходящие от спинномозгового нерва. Каждая такая ветвь соединяет спинномозговой нерв с одним из узлов симпатического ствола своей стороны. Эти симпатические узлы называют также околопозвоночными или паравертебральными ганглиями.
Часть аксонов преганглионарных нейронов заканчивается синапсами на телах ганглионарных нейронов соответствующих симпатических узлов. Остальные следуют транзитно через узел и продолжаются до других, более удаленных от позвоночника узлов, где происходит их переключение на эфферентные нейроны. Нейроны, расположенные в узлах симпатического ствола, дают начало постганглионарным безмиелиновым нервным волокнам, которые в составе серых соединительных ветвей направляются к каждому спинномозговому узлу. Часть постганглионарных нервных волокон в виде самостоятельных нервов идут либо к висцеральным сплетениям, либо к органам. В составе спинномозговых нервов постганглионарные вегетативные волокна направляются к сосудам и железам кожи, к скелетной мускулатуре, кровеносным сосудам. Постганглионарные волокна идут к иннервируемым органам тремя путями: в составе спинномозговых нервов, по ходу кровеносных сосудов или же в виде обособленных нервов, отходящих от симпатического ствола.
Шейный отдел симпатического стволадает вегетативные нервы, направляющиеся к кровеносным сосудам и органам головы и шеи - слезные, потовые и слюнные железы, к мышцы волос кожи, к мышце, расширяющей зрачок, органы шеи (гортань, глотка, сосуды).
Грудной отдел симпатического стволаучаствует вместе с шейными сердечными нервами в формировании сердечного сплетения, иннервирует пищевод, легкие, желудок, кишечник, почки и другие органы брюшной полости.
Поясничный и крестцовый отдел симпатического ствола несут иннервацию к органам малого таза.
Параганглии - это мелкие скопления в форме узелков хромаффинных
Рис. 34. Строение и области иннервации симпатической части вегетативной нервной системы.
клеток по ходу кровеносных сосудов и висцеральных сплетений. Они относятся к добавочным органам симпатической части автономной нервной системы; как и мозговое вещество надпочечников, параганглии выделяют в кровь катехоламины (адреналин и норадреналин). Катехоламины являются также основными нейромедиаторами, обеспечивающими симпатическую передачу нервных импульсов в симпатической части автономной нервной системы.
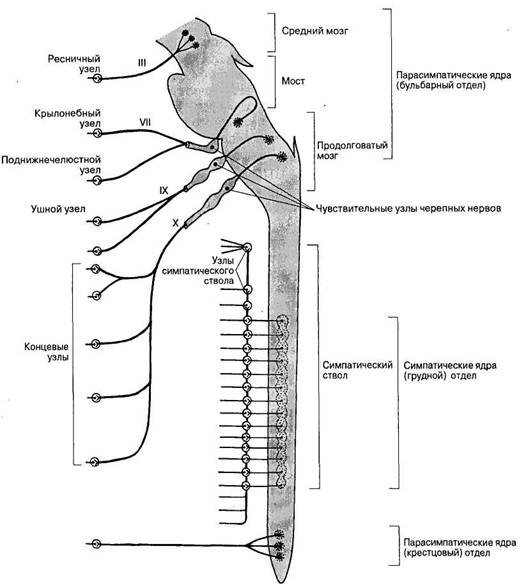
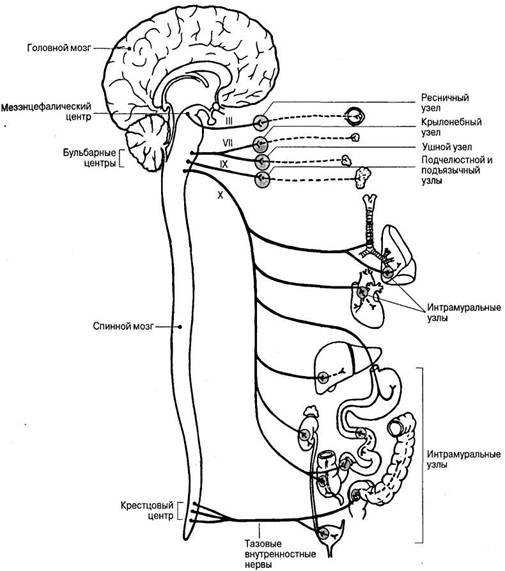
В парасимпатической части автономной нервной системы (рис. 35) существуют два основных очага выхода парасимпатических нервных волокон из ЦНС: краниальный (в области ствола мозга) и тазовый (в области крестцового отдела спинного мозга).
Краниальная часть включает парасимпатические ядра, расположенные в среднем мозге, а также в области моста и продолговатого мозга. Парасимпатические ганглии в отличие от симпатических, располагаются в непосредственной близости от иннервируемых органов или в самих этих органах.
Рис. 35. Строение и области иннервации парасимпатической части вегетативной нервной системы. Римскими цифрами обозначены черепные нервы.
Висцеральные сплетения и висцеральные узлы относятся к терминальной части автономной нервной системы и локализуются вдоль крупных кровеносных сосудов и около органов грудной и брюшно-тазовой полостей. Эти сплетения и ганглии образованы преимущественно нервными клетками и нервными волокнами симпатической нервной системы, однако в них присутствуют и парасимпатические нервные волокна, а также чувствительные нервные волокна из спинномозговых нервов, несущие интероцептивную сенсорную информацию от внутренних органов. В составе сплетений вегетативные нервные проводники непосредственно достигают иннервируемых органов. Различают висцеральные сплетения черепно-шейного, грудного, брюшного и тазового отделов.
В черепно-шейном отделе выделяют сплетение вокруг общей сонной артерии, которое подразделяется на внутреннее сонное и наружное сонное сплетения. Эти сплетения распространяются по ветвям сонных артерий. Вокруг подключичной артерии образуется подключичное сплетение, переходящее без резкой границы в плечевое автономное сплетение, дающее вегетативную иннервацию верхней конечности.
В грудном отделе выделяют сплетение грудной аорты, сердечное сплетение, легочное сплетение и пищеводное сплетение.
Сердечное сплетение устроено наиболее сложно. В иннервации сердца принимают участие симпатические и парасимпатические волокна.
В брюшном отделе выделяют: сплетение брюшной аорты, чревное сплетение, верхнее и нижнее брыжеечные сплетения, почечное сплетение, кишечное сплетение, подвздошное сплетение. Все эти сплетения располагаются по ходу одноименных артерий.
Наиболее крупное из них чревное, или солнечное, сплетение; оно является главным источником иннервации органов брюшной полости. Чревное сплетение располагается на передней стенке брюшной аорты в месте отхождения от нее чревного ствола и позади поджелудочной железы. В этих узлах происходит переключение большей части преганглионарных симпатических волокон на постганглионарные. волокна. Постганглионарные волокна от чревного сплетения образуют симпатические сплетения вокруг артерий, вместе с которыми они направляются ко всем органам брюшной полости.
В тазовом отделе выделяют: верхнее и нижнее подчревные сплетения, которые иннервируют органы малого таза: мочевой пузырь, прямая кишка, половые органы у мужчин и женщин.