Расчет разветвленных цепей упрощается, если пользоваться правилами Кирхгофа. Первое правило относится к узлам цепи. Узлом называется точка, в которой сходится более чем два тока. Токи, текущие к узлу, считается, имеют один знак (плюс или минус), от узла - имеют другой знак (минус или плюс).
𝐼1
𝐼2
Рис.3 𝐼3
Первое правило Кирхгофа является выражением того факта, что в случае установившегося постоянного тока ни в одной точке проводника и ни на одном его участке не должны накапливаться электрические заряды и формулируется в следующем виде: алгебраическая сумма токов, сходящихся в узле, равна нулю.

Второе правило Кирхгофа является обобщением закона Ома на разветвленные электрические цепи.
Рассмотрим произвольный замкнутый контур в разветвленной цепи (контур 1 – 2 – 3 – 4 – 1) (рис.2). Зададим обход контура по часовой стрелке и применим к каждому из неразветвленных участков контура закон Ома.

 +
+ 


В любом замкнутом контуре произвольной разветвленной электрической цепи, алгебраическая сумма падений напряжений (произведений сил токов на сопротивление) соответствующих участков этого контура равна алгебраической сумме эдс, входящих в контур.

Лекция 7. УГЛЕВОДЫ.
Вопросы:
1. Общие сведения.
2. Моносахариды.
3. Олигосахариды.
4. Полисахариды.
a. Углеводы – вещества состава СnН2nОn, имеющие первостепенное биохимическое значение, широко распространены в живой природе. К ним относятся различные сахаристые вещества, крахмал, целлюлоза (клетчатка).
Название углеводы сохранилось еще тогда, когда строение этих соединений еще не было известно, но установлен состав, имеющий общую формулу (СnН2О)n, т.е. углеводы.
Класс углеводов объединяют:
моносахариды – соединения, имеющие химическую природу гидроксиальдегидов или гидроксикетонов. Они существуют преимущественно в таутомерных циклических формах;
олигосахариды – продукты конденсации нескольких молекул моносахаридов друг с другом с выделением воды (по типу простых эфиров); важнейшими олигосахаридами являются дисахариды, молекулы которых построены из двух остатков моносахаридов;
полисахариды – высокополимерные вещества, продукты конденсации большого числа остатков моносахаридов.
Моносахариды в зависимости от числа входящих в их состав атомов кислорода (обычно это число равно числу атомов углерода) разделяют: на группы тетроз, пентоз и гексоз.
В зависимости от того имеется ли в молекуле моносахаридов альдегидная или кетонная группы, их разделяют на альдозы и кетозы.
Классификация углеводов такова:
| Моносахариды
| Дисахариды С12Н22О11
|
| Тетрозы С4Н8О4
| сахароза
|
| эритроза
| лактоза
|
| треоза
| мальтоза
|
| Пентозы С5Н10О5
| Полисахариды
|
| арабиноза
| целлюлоза (С5Н10О5)n
|
| ксилоза
| крахмал
|
| рибоза
| гликоген
|
| Гектозы С6Н12О6
|
|
| глюкоза
|
|
| галактоза
|
|
| фруктоза
|
|
Из всего разнообразия моносахаридов наиболее распространены альдопентозы и альдогектозы. Самой распространенной альдогектозой является глюкоза. Глюкоза – виноградный сахар, т.к. она содержится в виноградном соке, не менее распространена глюкоза и в животном мире: около 0,1 % ее содержится в крови. Глюкоза распространяется кровью по всему организму и служит источником энергии для организма.
Другой моносахарид – фруктоза широко распространен в природе. Фруктоза – кетогектоз или плодовый сахар.
Другие моносахариды почти не встречаются в свободном виде, но входят в состав важных олиго – и полисахаридов. Например, рибоза имеет очень большое биологическое значение. Из-за своей связи с нуклеиновыми кислотами.
Галактоза входит в состав дисахарида лактозы, т.е. молочного сахара. Моносахариды способны существовать в ряде таутомерных форм, что приводит к появлению большого числа оптически активных стереоизомеров. Например, вещества состава С6Н12О6 может образовывать около 100 структурно и пространственно изомерных форм таутомерных модификаций.
Моно- и дисахариды – бесцветные кристаллические вещества, хорошо растворимые в воде, но не растворимые в неполярных органических растворителях. При нагревании до температур порядка 120-1500С они не разлагаются, а плавятся, образуя карамели. Оптически активны, обладают сладким вкусом. Тростниковый или свекловичный сахар – типичный представитель дисахаридов – сахароза.
Полисахариды – высокомолекулярные соединения. Они не плавятся, образуют коллоидные растворы. Главная химическая способность омно – и полисахаридов – их способность к гидролизу с образованием моносахаридов. В природе углеводы образуются в процессе фотосинтеза: так называют превращение углекислоты воздуха за счет солнечной энергии в сахара. Этот процесс идет в зеленых листьях на солнечном свету. Катализатором служит хлорофилл – зеленый пигмент листьев. Синтетические способы получения углеводов не имеют практического значения, хотя способы их получения давно известно.
b. Моносахариды, которые содержат альдегидную группу (рибоза, дезоксирибоза, глюкоза), восстанавливают аммиачный раствор Ag2O (реакция «серебряного зеркала»). Качественная реакция на глюкозу
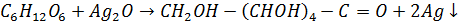
глюконовая кислота
ОН
Взаимодействие глюкозы с гидратом меди (II) в щелочном растворе с образованием алкоголятом.
Качественная реакция на глюкозу
O
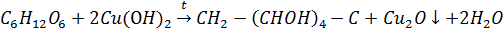
ОН OН
морковный цвет
При этом альдегидная группа, как обычно превращается в карбоксильную группу. Все углеводы без нагревания образуют синие растворимые комплексные соединения с  в щелочной среде.
в щелочной среде.
Постепенное окисление глюкозы с медленным выделением энергии:
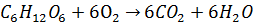 (
(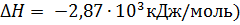
Брожение глюкозы:
дрожжи
1) спиртовое: 
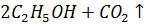
фермент
2) молочнокислое: 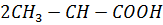
OH
молочная кислота
фермент
3) маслянокислое: 
воздух масляная кислота
Рибоза – моносахарид  - бесцветное кристаллическое вещество, растворима в воде, спирте, нерастворима в эфире. Рибоза входит в состав рибонуклеиновой кислоты в циклической форме, строение которой таково:
- бесцветное кристаллическое вещество, растворима в воде, спирте, нерастворима в эфире. Рибоза входит в состав рибонуклеиновой кислоты в циклической форме, строение которой таково:

Дезоксирибоза – моносахарид  – бесцветное кристаллическое вещество. Растворима в воде и спирте, нерастворима в эфире. Входит в состав дезоксирибонуклеиновой кислоты (ДНК) в циклической форме.
– бесцветное кристаллическое вещество. Растворима в воде и спирте, нерастворима в эфире. Входит в состав дезоксирибонуклеиновой кислоты (ДНК) в циклической форме.
Глюкоза (виноградный сахар) – наиболее широко распространенный моносахарид. Может существовать в открытой и двух циклических формах. Обычная форма глюкозы 

Обе циклические формы можно получить в кристаллическом виде, при этом α – форма более устойчива и обычна. В водяном растворе при комнатной температуре содержится 36 % α – формы, около 0,02 % открытой формы и 64 % β – модификации. Крахмал образован исключительно остатками α – формы глюкозы, а целлюлоза - β – формы.
Фруктоза (фруктовый сахар) – моносахарид, содержащий кетонную группу (кетогексоза). Формула в обычном виде  , в растворе существует в виде смеси двух циклических и одной линейной форме.
, в растворе существует в виде смеси двух циклических и одной линейной форме.
Пятичленные циклические формы фруктозы называют фуранозами:

Фруктоза – бесцветное кристаллическое вещество. Хороша растворима в воде. По сладости в 1,5 раза превышает обычный сахар (сахарозу).
Галактоза – альдогексоза, один из пространственных изомеров глюкозы. Входит в состав дисахарида лактозы.

3. Дисахариды – соединения, получаемые конденсацией двух молекул моносахаридов. Моносахариды, образующие дисахарид, входят в него в одной из циклических форм. Если у одного из них альдегидная группа остается не затронутой при конденсации, как у лактозы, дисахарид называется восстанавливающим, поскольку вступает в реакции «серебряного зеркала» и с Cu(OH)2 в щелочном растворе. В противном случае дисахарид называется невосстанавливающим (сахароза).
В состав дисахарида лактозы (молочный сахар) входят остатки глюкозы и галактозы.

лактоза
Сахароза (свекловичный и тростниковый сахар) – невосстанавливающий дисахарид, образованный остатками глюкозы и фруктозы в циклической форме.

остаток остаток
α – глюкозы β – фруктозы
Сахароза – бесцветное кристаллическое вещество, растворима в воде, спирте малорастворима и нерастворима в эфире. В пределах 190 – 2000С разлагается с образованием «жженного сахара».
Под действием кислот подвергается гидролизу до смеси глюкозы и фруктозы, та же реакция протекает при образовании меда пчелами. Эта реакция получила название инверсии, поскольку направление вращения плоскости поляризованного света у исходной сахарозы и получаемой смеси глюкозы и фруктозы противоположно, а ее продукт получил название инвертного сахара – продукта гидролиза сахарозы:

глюкоза фруктоза
Лактоза – восстанавливала дисахарид, образованный формами глюкозы и галактозы.
Полисахариды – высокомолекулярные соединения, при полном гидролизе образуют молекулы моносахаридов 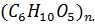 Полисахариды – продукты поликонденсации моносахаридов. Обычно состоят из остатков циклической формы одного моносахарида, чаще всего глюкозы. Степень полимеризации составляет примерно 100 – 1400 – это менее разветвленная форма, иногда называемая амилазой или от 1000 – 6000 форма с более разветвленными цепями, называемая амилопектином.
Полисахариды – продукты поликонденсации моносахаридов. Обычно состоят из остатков циклической формы одного моносахарида, чаще всего глюкозы. Степень полимеризации составляет примерно 100 – 1400 – это менее разветвленная форма, иногда называемая амилазой или от 1000 – 6000 форма с более разветвленными цепями, называемая амилопектином.
Крахмал – количество изомеров – несколько тысяч, молекулы бывают линейными и разветвленными. Крахмал аморфный нерастворимый в холодной воде и образующий коллоидный раствор в горячей воде. При нагревании с разбавленной кислотой или под действием ферментов постепенно гидролизуется с образованием сначала декстрина – смеси олигомеров с небольшой длиной цепи и, в конце концов, мономера – глюкозы.
Характерным для крахмала является взаимодействие с йодом с образование синего соединения неопределенного состава.
Гликоген (животный крахмал) – полимер глюкозы, близкий к крахмалу, имеющему большую степень полимеризации (25000- 90000 остатков глюкозы) и менее разветвленные цепи. В организме человека и животных используется для регулирования содержания в крови глюкозы: при избытке последней она связывается в гликоген, при недостатке – выделяется из последнего в кровь.
Лекция 8. АМИНОКИСЛОТЫ, ПЕПТИДЫ И БЕЛКИ.
Вопросы:
5. Основные сведения.
6. Свойства аминокислот.
7. Пептидная связь. Пептиды.
8. Белки.
1. Аминокислоты – соединения, в молекулы которых входят аминогруппы и карбоксильные группы.
Кроме обычной системы обозначений, часто используется другая, в которой положение аминогруппы относительно карбоксильной группы обозначается греческими буквами:  - аминокислоты (аминогруппа у соседнего с карбоксилом атома углерода),
- аминокислоты (аминогруппа у соседнего с карбоксилом атома углерода),  - аминокислоты (через один атом углерода),
- аминокислоты (через один атом углерода), 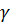 - аминокислоты (через два) и т.д.
- аминокислоты (через два) и т.д.
Соединения, в которых аминогруппа и карбоксильная группа находятся на противоположных концах цепи, обычно называют 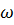 - аминокислотами.
- аминокислотами.
Около 150 аминокислот были обнаружены в живых организмах, около 20 из них входят в состав белков. Половина этих аминокислот незаменимы (не синтезируются в организме человека), они поступают с пищей.
Синтетические аминокислоты получают кислотным гидролизом белков, либо из карбоновых кислот, воздействуя галогеном и, долее, аммиаком.
Наибольшее практическое значение имеют - и некоторые - аминокислоты: первые как составные части пептидов и белков; вторые как сырьё для получения полимерных материалов.
2. Физические свойства аминокислот:
· белые кристаллические вещества;
· хорошо растворяются в воде, а в органических растворителях (даже в спирте) плохо растворимы;
· довольно высокая температура плавления (220  230)0С, иногда связана с разложением.
230)0С, иногда связана с разложением.
Большинство - аминокислот содержат в молекуле ассиметричный атом углерода и образуют оптические изомеры.
Химические свойства
Поскольку молекулы аминокислот содержат кислотную (- СООН) и основную (- NН)2 группы, они ведут себя как амфотерные соединения. Как в твердом состоянии, так и в растворах они существуют в виде внутренних солей, или биополярных ионов  , поскольку протон карбоксильной группы переходит к атому азота аминогруппы. Такая структура объясняет высокую температуру плавления аминокислот и нерастворимость их в некоторых неполярных растворителях.
, поскольку протон карбоксильной группы переходит к атому азота аминогруппы. Такая структура объясняет высокую температуру плавления аминокислот и нерастворимость их в некоторых неполярных растворителях.
Аминокислоты, как амфотерные соединения, могут проявляться как кислотные свойства, обусловленные наличием в их молекулах карбоксильной группы – СООН, так и основные свойства, обусловленные аминогруппой – NН2
Аминокислоты взаимодействуют с кислотами и щелочами:
1. Реакции аминогруппы – реакции образования солей с кислотами
СН2  СН2
СН2  СООН Cl-
СООН Cl-
(хлороводородная соль глицина)
NH2 N+H3
2. Реакция карбоксильной группы – реакции образования солей с основаниями
СН2  СН2 COONa + H2O
СН2 COONa + H2O
NH2 NH2 (натриевая соль глицина)
3. Реакции этерификации – реакции образования сложных эфиров
О
СН2  СН2 C + H2O
СН2 C + H2O
NH2 NH2 О 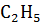
(этиловый эфир глицина)
Получение аминокислот Хотя для получения - аминокислот разработаны синтетические методы, на практике используется почти исключительно гидролиз белков. Получаемая смесь аминокислот разделяется методами хроматографии, как способе разделения газообразных и растворенных веществ, основанном на последовательной многократной адсорбции и десорбции, происходящей при медленном протекании потока газа или раствора сквозь твердый сорбент. Таким сорбентом может быть целлюлоза, входящая в состав фильтрованной бумаги, активированный уголь, специально подготовленный силикагель и оксид алюминия. Аминокислоты могут быть получены из карбоновых кислот по следующим реакциям:
СН3СООН  CH2
CH2  CН2 СООН
CН2 СООН
Cl NH2
При отсутствии в молекуле дополнительных кислотных или основных функциональных групп и кислотные, и основные свойства выражены слабо (К  поскольку в обычной структуре молекулы карбоксильной группы и аминогруппы взаимно нейтрализованы.
поскольку в обычной структуре молекулы карбоксильной группы и аминогруппы взаимно нейтрализованы.
3. Пептидная связь – результат реакции конденсации молекул - аминокислот (одинаковых или различных). Например,

За счет пептидной связи – аминокислоты образуют пептиды и белки.
Аминокислоты, являющиеся составными частями белков, имеют общую структуру 
Строение радикала R двадцати – аминокислот, входящих в природные белки и пептиды указанны в таблице 1. Эти двадцать аминокислот составляют основу жизни.
Таблица 1
| № п/п
| Название кислоты
| R- радикал
|
|
| Глицин
| - Н
|
|
| Аланин
| - СН3
|
|
| Серин
| - СН2ОН
|
|
| Треонин
| - СН(ОН)СН3
|
|
| Метионин
| - (СН2)2SСН3
|
|
| Валин
| - СН(СН3)2
|
|
| Лейцин
| - СН2СН(СН3)2
|
|
| Изолейцин
| - СН(СН3)С2Н5
|
|
| Фенилаланин
| - СН2С6Н5
|
|
| Тирозин
| - СН2С6Н4ОН (пара)
|
|
| Цистеин
| - СН2SН
|
|
| Аспарагиновая кислота
| - СН2СООН
|
|
| Глютаминовая кислота
| - (СН2)2 СООН
|
|
| Аргенин
| - (СН2)3NHС(NH)NH2
|
|
| Лизин
| - (СН2)4NH2
|
|
| Гистидин
| - СН2С3N2Н3
|
|
| Пролин (полная структура)
| 
|
|
| Триптофан
| - СН2С8NН6
|
|
| Аспаргин
| - СН2СОNН2
|
|
| Глутамин
| - (СН2 )2СОNН2
|
Организм человека в состоянии синтезировать все аминокислоты, входящие в состав своих белков, за исключением следующих девяти: гистидина, лизина, триптофана, фенилаланина, лейцина, изолейцина, треонина, метионина и валина.
Эти аминокислоты должны получаться с пищей (суточная потребность колеблется в пределах от 1 до 4-5 г). Белки, не содержащие одной или более из указанных аминокислот, например, желатин, не содержащий триптофана, валина и треонина, или многие растительные белки, не содержащие лизина, являются неполноценными по своим питательным свойствам.
Пептиды – продукт конденсации аминокислот с образованием пептидной связи. Низшие пептиды по свойствам близки к аминокислотам. При увеличении длины цепи у пептидов появляется вторичная структура, и пептиды достаточно длинной цепью по свойствам приближаются к белкам, к которым они находятся примерно в таком же отношении, как омпомеры к полимерам.
Белки – высокомолекулярные (с молекулярной массой от 10 000до миллионов) природные полипептиды.Структура белков, как полипептидов:
1) первичная структура – конкретная последовательность остатков – аминокислот в молекуле, соединенных пептидными связями.
2) вторичная структура – упаковка спиральной полипептидной цепи в пространстве. Пространственное строение определяется водородными связями, образующимися между группами NH и С = 0, входящими в пептидную связь. Обычно это спираль.
3) третичная структура – трехмерная конфигурация благодаря дополнительного свертывания спиральной боковой цепи в результате разнообразных взаимодействий боковых цепей аминокислот. Разрушение вторичной и третичной структур (денатурация), происходящее нагревании, радиации, сильных кислот и щелочей и т.д., приводит к потере характерных свойств белка, несмотря на сохранение первичной структуры;
4) четвертичная структура – это объединение нескольких полипептидных цепей, не связанных химически, для выполнения биохимической роли белков (например, гемоглобин состоит из четырех полипептидных цепей).
Характерные химические реакции на белки:
Цветные реакции:
1) биуретовая реакция: белок + CuSO4 + NaOH  фиолетовое окрашивание;
фиолетовое окрашивание;
2) ксантопротеиновая реакция: белок + HNO3(конц.) желтое окрашивание, если белок содержит бензольные кольца.
Применение. Человек получает необходимые аминокислоты в составе белков пищевых продуктов. В медицине: для питания тяжелобольных и в качестве лекарства. Белки находят также применение для производства синтетических волокон.
Гетероциклические соединения
Гетероциклические соединения – циклические соединения, в которых один или более из атомов, образующих кольцо, не является углеродным.
Пиримидиновые и пуриновые основания – производные более сложных азотосодержащих гетероциклических соединений пиримидина и пурина, входящих в состав нуклеиновых кислот
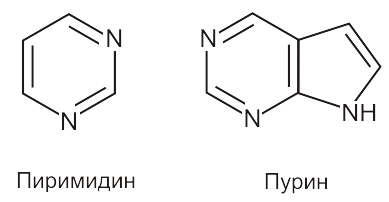
Пиримидиновые основания:
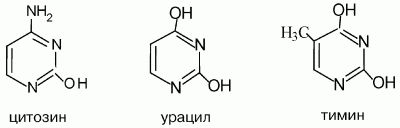
Пуриновые основания:

В молекулах нуклеиновых кислот (ДНК и РНК) каждое из пиримидиновых оснований благодаря комплементарности (взаимной дополнительности) геометрического строения и относительного расположения атомов водорода, кислорода и азота может образовывать специфические конфигурации водородных связей с определенным пуриновым основанием

Водородные связи показаны точками, урацил ведет себя аналогично тимину.
Лекция 11. ЖИВЫЕ СИСТЕМЫ
1. Происхождение жизни
2. Эволюция живых систем
3. История жизни на Земле и методы исследования эволюции
4. Генетика и эволюция
1. С античных времен до наших дней идут споры о том, как возникла жизнь на Земле. Просто и радикально решают эту проблему креационисты, считающие, что жизнь является результатом божественного творения.
Альтернативой такой концепции является представление о том, что жизнь во Вселенной существовала вечно – концепция стационарного состояния, а на Землю живая материя была занесена из космоса – концепция панспермии. Долгое время в естествознании господствовала гипотеза самопроизвольного (или спонтанного) зарождения жизни. Суть концепции такова, что жизнь возникла и продолжает постоянно и многократно возникать из неживого (косного) вещества. В противовес этой концепции выступил итальянский биолог и врач Франческо Реди, который сформулировал концепцию биогенеза, в соответствии с которой жизнь может возникнуть только из предшествующей жизни («все живое от живого»).
В XIX веке общепринятой стала концепция абиогенного происхождения жизни на Земле, основанная на гипотезе происхождения жизни путем биохимической эволюции, выдвинутая независимо российским академиком Опариным и английским ученым Холдейном. Академик Опарин полагал, что органические вещества могли создаваться в океане из более простых неорганических соединений. Энергия этих реакций синтеза могла быть получена от интенсивной солнечной радиации. Другим вариантом является синтез содержащих углерод и азот веществ в расплавленных глубинах Земли с последующим выносом их в океан вулканами.
Концепция биохимической эволюции основана на том, что первичная атмосфера Земли носила восстановительный, а не окислительный характер, в ней не было кислорода. Атмосфера безжизненной Земли в основном состояла из водородных соединений: паров воды (Н2О), аммиака (NH3),сероводорода (H2S) и т.д. Из смеси этих паров, в присутствии электрических разрядов молний, образовались первые аминокислоты и пиримидиновые основания, которые являются кирпичиками для построения белков и нуклеиновых кислот. Эти соединения около 3,5 млрд лет назад превратили гидросферу в раствор разнообразных органических соединений, образовался «питательный бульон». В этом бульоне постепенно выделялись и отделялись от окружающего раствора молекулярные образования в форме видимых под микроскопом коацерватных капель – предшественников клетки.
Коацерваты – жидкие коллоидные скопления в растворе высокомолекулярных соединений, обогащенные растворенным веществом.
Коацерваты, при всей сложности их организации, не были живыми организмами прежде всего потому, что у них нет стабильного самовоспроизведения, жесткой структурной организации, функционального взаимодействия между белками и нуклеиновыми кислотами.
Появление истинно живых систем – протобионтов – происходило около 3,0 млрд лет назад. У протобионтов уже появляется корреляция между нуклеиновыми кислотами и белками; способность синтезировать белки определенного строения в соответствии с информацией, заключенной в нуклеиновый кислоте. Одновременно у них совершенствуется мембранный аппарат, обеспечивающий упорядоченность обмена веществ, поддержание стабильности системы. И самое главное они приобретают способность к самовоспроизведению.
Структурное усложнение и функциональное совершенствование пробионтов привело к появлению организмов, имеющих клеточную организацию – первичных прокариотных (безъядерных) организмов – бактерий. С этого момента начинает осуществляться биологическая эволюция организмов.
Единственным источником питания для первичных организмов могли вначале служить только органические вещества, которые раньше возникли чисто абиогенным путем (гетеротрофное питание), т.к. способность к органическому питанию заложена в самой основе жизни, присуще всем без исключения живым существам. Отсутствие свободного О2 в первичной земной атмосфере и гидросфере обусловило анаэробный характер энергетического обмена первичных организмов.
В процессе развития жизни запас абиогенно образовавшихся органических веществ на земной поверхности постепенно истощался, т.к. развитие жизни шло быстрее образования веществ. Это изменение условий существования выдвинуло на первый план развития такие организмы, которые благодаря приобретенной или способности поглощать свет получили возможность строить заново органические вещества из неорганических соединений атмосферы – СО2 и водяных паров, возникает новый биологический метод синтеза* этих веществ – фотосинтез, который осуществляется на основе обмена веществ очень совершенными путями и поэтому в дальнейшем приобрел главенствующее, монопольное значение, которое сохранил до наших дней.
Возникновение фотосинтеза изменило всю обстановку жизни на Земле. Часть организмов сама стала строить им необходимые органические соединения (автотрофы), др. часть сохранила прежние формы питания, используя органические вещества (гетеротрофы).
Возникновение фотосинтеза не только создало изобилие органических веществ, но и привело к появлению свободного кислорода, до этого отсутствовавшего на земной поверхности. Это изменило весь характер происходивших здесь химических процессов и позволило большинству живых существ значительно свой энергетический обмен, надстроив над прежним анаэробным обменом новые системы кислородного дыхания целиком используя скрытую в органических веществах энергию.
2. Под биологической эволюцией понимают необратимый процесс исторического развития живой природы начиная с момента абиогенного возникновения первых живых организмов на Земле до настоящего времени.
В XVII – XVIII в.в. в биологии и философии был распространен трансформизм: представление об изменяемости видов организмов под воздействием окружающей среды, к которым организмы приспосабливаются благодаря заложенной в них изначально способности целесообразно реагировать на внешние воздействия, а приобретенные этим путем изменения наследуются.
В начале XIX в. появляются первые эволюционные теории. Особое значение для дальнейшего развития эволюционной идеи имела теория Ж.–Б. Ламарка. Своеобразие его концепции заключается в объединении идеи изменяемости органического мира (трансформизм) с представлением о градации – постепенном повышении уровня организации от самых простых до наиболее сложных и совершенных организмов. Отсюда вывод, что изменения организмов имеют не случайный, а закономерный характер, в направлении постепенного совершенствования и усложнения организации. Движущей силой градации Ламарк считал «стремление природы к прогрессу», которое заложено Творцом.
Крупнейшим событием в науке явилось появление научной теории исторического развития живой природы – эволюционной теории Ч. Дарвина (1859 г.). Заслуга Дарвина заключается в том, что он вскрыл ее сущность и выстроил систему доказательств эволюции.
Основные положения эволюционной теории Дарвина:
1. Всем живым организмам окружающего нас мира свойственна наследственная изменчивость. Стимулом для возникновения изменчивости служат изменения внешней среды, но характер изменчивости определяется спецификой самого организма, а не направлением внешних условий.
2. В центре внимания эволюционной теории находятся не отдельные организмы, а биологические виды и внутривидовые группировки - популяции.
3. Все виды организмов в природе вынуждены вести жесткую борьбу за свое существование. Эта борьба для особей данного вида складывается из их взаимодействия с неблагоприятными биотическими и абиотическими факторами внешней среды, а также из их конкуренции между собой.
4. Естественный отбор – это особый механизм выживания и воспроизведения организмов в природе, отбор в ходе эволюции наиболее приспособленных к условиям среды и гибель неприспособленных, следствие борьбы за существование.
5. Следствиями борьбы за существование и естественного отбора являются: 1) развитие приспособлений видов к условиям из существования; 2) дивергенция (развитие от общего предка несколько дочерних видов, все большее расхождение их признаков в эволюции) и 3) прогрессивная эволюция (усложнение и усовершенствование организации).
6. Частным случаем Е.О. (естественный отбор) является половой отбор, который обеспечивает развитие признаков, связанных с функцией размножения.
7. Породы домашних животных и сорта с/х растений созданы посредством искусственного отбора, аналогичного естественному отбору, но ведущегося человеком в своих интересах.
Развитие дарвинская теория эволюции Дарвина получила в синтетической эволюции (СТЭ), сложившейся к 1940 – м годам.
Основные положения СТЭ
1. Материалом для эволюции служат дискретные изменения наследственности – мутации – это стойкие изменения наследственных структур живой материи, ответственных за хранение и передачу генетической информации. Мутационная изменчивость носит случайный характер.
2. Ведущим движущим фактором эволюции является естественный отбор, основанный на селекции случайных и мелких мутаций.
3. Наименьшая эволюционная единица – популяция.
4. Эволюция в основном носит дивергентный характер, т.е. один вид может стать предком нескольких дочерних видов, но каждый вид имеет один единственный предковый вид, единственную предковую популяцию.
5. *Эволюция носит постепенный и длительный характер.
6. Вид состоит из множества соподчиненных морфологических, физиологических и генетических отличных, но репродуктивно не изолированных единиц – подвидов, популяций.
7. Обмен генами возможен лишь внутри вида. Если мутация имеет положительную селективную ценность на территории всего ареала вида, то она может распространяться по всем его подвидам и популяциям.
8. Макроэволюция, или эволюция на уровне выше вида, идет лишь путем микроэволюции, под которой понимают видообразование.
9. Каждая систематическая единица (вид, род, и т.д.) должна иметь единственный корень.
10. * Эволюция не предсказуема: она не направлена к некой конечной цели.
Мутационный процесс и другие факторы (миграция особей, изоляция и случайный дрейф генов), создают генетическую неоднородность внутри вида.
Существует лишь один направленный фактор эволюционного процесса – естественный отбор, который действует в пределах популяции. Естественный отбор - единственный эволюционный процесс, приводящий к повышению приспособленности организмов. Действие естественного отбора основана на статических закономерностях. Оно сводится к дифференциальному размножению особей, а это означает, что носители одних генотипов имеют больше шансов выжить и оставить потомство, чем носители других генотипов. При этом революционную роль играет не выживание особей, а вклад каждой особи в генофонд популяции.
СТЭ выделяет три основные формы Е.О. имеющие разный эволюционный результат: движущий, дизруптивный и стабилизирующий.
Популяционные волны (их иногда называют волнами жизни) – это количественные колебания численности популяций под воздействием различных проявлений внешней среды – сезонных изменений климата, различных природных или техногенных катастроф и т.д. Колебания численности популяций имеют место и между компонентами биоценоза, когда один из компонентов – жертва – служит пищей для другого – хищника.
Изоляция, или обособленность, группы организмов означает ограничение свободы скрещиваний. Изоляция определенной группы организмов необходима для того, чтобы воспрепятствовать скрещиванию с другими видами и передачи им, как и получено от них генетической информации. Причины изоляции могут быть любыми: географические границы (неопредолимая водная среда, болота, высокие горы), экологические условия (предпочтения в выборе экологической ниши или места обитания), разные периоды спаривания, особенности поведения разных групп организмов. Изоляция препятствует снижению межпопуляционных генотипических различий и является условием сохранения, закрепления и распространения в популяциях повышенной жизнеспособности. В п






