Эволюция нейрона и филогенетические типы
Нервных систем
1.1. Филогенез нервной клетки
1.2. Филогенез нервной системы
1.2.1. Диффузный тип нервной системы
1.2.2. Ганглионарный тип нервной системы
1.2.3. Трубчатый тип нервной системы
Термин филогенез означает историческое развитие. Когда мы говорим о филогенезе нейрона, нервной ткани, нервных систем, мы подразумеваем эволюционные процессы формирования исследуемых структур и изменения, возникающие в ходе этого развития.
Филогенез нервной клетки
На сегодняшний день совершенно очевидным является тот факт, что предшественником нервной клетки является железистая, секреторная клетка. Химический способ передачи информации – это самый древний эволюционный механизм, обеспечивающий как взаимодействие клеток внутри организма, так и перенос информации на значительные расстояния, позволяющий общаться различным организмам на языке химии, например при помощи феромонов. Такой способ передачи сигнала принято считать дистантным. Молекулярная специфика переносимой информации определила эволюционное развитие соответствующего воспринимающего рецепторного аппарата. Дистантный способ передачи информации в нервных сетях сохранился и у самых высокоорганизованных организмов в виде нейроэндокринных (нейросекреторных) клеток. Биологически активные вещества – нейрогормоны выделяются в кровь, а затем депонируются в нейро-гемолимфатических или нейрогемальных органах, таких как кардиальные тела насекомых, синусная железа рака, нейрогипофиз млекопитающих. По мере совершенствования организмов количество нейросекреторных клеток резко сокращается, а их локализация ограничивается строго определенным отделом центральной нервной системы.
Секреторные клетки по своему происхождению в большинстве своем являются эпителиальными структурами, объединенными различными клеточными контактами в единую систему. Нервные клетки в ходе закладки и развития нервных систем развиваются из эктодермального или энтодермального эпителия. Типы клеточных контактов, создаваемых нейронами, усложняются и обеспечивают эволюционное преобразование простых нервных сетей в сложные нервные системы. У кишечнополостных нейроны оказываются связанными в единую сеть при помощи плотных замыкающих контактов – анастомозов. В ней даже присутствуют цитоплазматические мостики между соседними нервными элементами, наподобие синцития. Однако электоронномикроскопические исследования показывают наличие в таких нервных сетях и синаптических контактов. Они могут быть как асимметричными, так и симметричными, электрическими и химическими. Синаптические контакты характеризуются тем, что рецепторные, нервные и эффекторные клетки, принимающие участие в их формировании, специализированы на восприятие и передачу специфической нервной информации. Общим признаком таких клеточных контактов является обязательная электоргенность мембран, а в синапсах химического типа – наличие в пресинаптическом компоненте медиатора (нейротрансмиттера, синаптичеcкого передатчика). Такой способ передачи информации принято называть контактным.
Существует определенная зависимость между синаптическими характеристиками и эволюционной продвинутостью животных. Так, у примитивных беспозвоночных много электрических синапсов. В химических синапсах щель может достигать нескольких сот нанометров. Медиатором чаще всего является ацетилхолин или один из биогенных моноаминов, причем в ацетилхолиновых синапсах нет фермента холинэстеразы, расщепляющего ацетилхолин у позвоночных до холина и уксусной кислоты. В теле беспозвоночных инактивация медиатора происходит за счет ферментов, растворенных в гемолимфе. У позвоночных электрические синапсы встречаются в ограниченном количестве. Химические контакты отличаются разнообразием медиаторов и сложным рецепторным аппаратом постсинаптического компонента. Рецепторы постсинапса приобрели способность распознавать различные нейротрансмиттеры, вступать с ними во взаимодействия, вызывая эффекты деполяризации или гиперполяризации клеточной мембраны.
Таким образом, нейроны, из которых простроены реальные нервные системы, – это эволюционно модифицированные железистые клетки, специализированные для синтеза и выделения физиологически активных веществ. Различие между медиатором и нейрогормоном достаточно условно. Медиатором вещество называют в том случае, когда оно действует контактно, т.е. областью выделения является синаптическая щель, мишенью – постсинапс. Нейрогормоны действуют дистантно. В этом случае нервная клетка образует химический контакт с эндотелием сосуда, т.е. нейросекрет выделяется в кровь (или гемолимфу) и ее потоком доносится до самых отдаленных мишеней. Одно и то же химическое вещество может выступать как в роли нейромедиатора, так и в роли нейрогормона; более того, это может быть даже продукт одного нейрона.
В историческом развитии и усложнении типов нервных сетей у позвоночных стали преобладать нейроны трансмиттерного типа, в то время как у беспозвоночных преобладают нейроны нейросекреторного типа. Важно отметить, что образование нейросекрета в нейронах беспозвоночных ограничивается временем сезонной половой активности. Этот же процесс наблюдается у позвоночных с четко выраженной сезонной половой активностью: нейрокринные клетки синтезируют разнообразные регуляторные пептиды, которые накапливаются в нейро-гемолимфатических и нейрогемальных органах. Под их контролем в частности находится гаметогенез.
Третьим типом клеток, которые также наиболее широко представлены у беспозвоночных, являются нейроны смешанного типа. Они функционируют одновременно как трансмиттерные и нейрокринные клетки. Синапсы этих нейронов могут быть как химическими, так и электрическими. Например, в нервных системах двустворчатых моллюсков часто встречаются нейроны, которым свойственна и нейросекреция, и передача нервного импульса на базе ацетилхолина. Известно, что в так называемых электрических синапсах взаимодействие между клетками идет при посредстве электрического тока. В хорошо исследованных случаях оказывалось, что нейроны, вступающие в электрическую связь, – это все-таки секреторные клетки, однако секреция у них происходит не там, где находятся электрические синапсы. Так, нейроны, управляющие электрическими органами электрических рыб, должны секретировать ацетилхолин синхронно, поэтому их дендриты связаны электрическими синапсами, секрецию при этом ведут аксонные окончания, иннервирующие электроплаксы (электрические пластинки). Очень часто электрически бывают связаны изохимичные (имеющие одинаковый химизм) нейроны. Этот механизм обеспечивает одновременный выброс секрета, запасенного всеми связанными клетками. В результате достигается высокая концентрация нейрогормона в крови или медиаторов в нейропиле. Нельзя, конечно, с полной уверенностью утверждать, что не существуют нейроны, лишенные способности к секреции. Однако это скорее будет исключением из правила.
Филогенез нервной системы
1.2.1. Диффузный тип нервной системы. В ходе исторического развития жизни на Земле первыми животными, у которых появилась нервная система, являются примитивные кишечнополостные: гидроидные полипы, актинии. Их одиночные нейронырасположены в промежутке между экто- и энтодермой. Поскольку нейроны образуют некоторое множество, их довольно условно объединяют в нервную сеть простейшего типа. Однако даже такие примитивные одиночные нервные клетки имеют отросток, оканчивающийся на эффекторе – эпителиомиоците. Существует мнение, что эволюционным предшественником нейрона могла быть миоэпителиальная клетка. Появившись в составе пограничных тканей, миоэпителиальная клетка могла дивергировать в направлении возникновения самостоятельных нервной и мышечной единиц. Возможно, эта дивергенция произошла путем разделения миоэпителиоцита на эпителиальный секреторный предшественник нейрона и миоцит тем же механизмом, который определяет цитотомию соматической клетки в митозе. Тесные связи нервных и мышечных элементов сохранились и у всех более высокоорганизованных животных. В ходе всей дальнейшей эволюции нервных систем соблюдается это элементарное правило – дифференцированный нейрон существует как жизнеспособная клеточная единица только в том случае, если он имеет контакты с другими клетками, в частности с эффекторами.
Более продвинута организация нервной системы у медуз: нейронная сеть расположена в наружных покровах и топографически считается эктодермальной сетью диффузного типа. Каждая нервная клетка имеет длинные отростки (один или несколько) и соединена с другими нервными клетками.
Нейроны кишечнополостных являются изополярными, что означает отсутствие специализации у их отростков, а, следовательно, отростки проводят возбуждение в любую сторону и не образуют длинных проводящих путей. Контакты между нервными клетками в такой сети разнообразны: анастомозы – цитоплазматические контакты, обеспечивающие непрерывность сети, щелевидные контакты – подобны синапсам. Последние также бывают двух видов: симметричные синапсы – содержат синаптические пузырьки по обе стороны контакта, несимметричные синапсы – имеют везикулы только с одной стороны щели.
В примитивных нервных системах нервные импульсы распространяются по поверхности нейронов с одинаковой скоростью. Благодаря своей изополярности униполярные нейроны таких сетей проводят афферентные и эфферентные импульсы поочередно, последовательно сменяя направление распространения волны возбуждения. Это сопровождается довольно протяженным временем ответной реакции.
Диффузный тип нервной системы характеризуется следующими признаками.
1. Нервные клетки равномерно распределены в теле животного. У кишечнополостных имеется два неоформленных скопления нервных клеток – в районе подошвы и вокруг ротового отверстия.
2. Проведение возбуждения во всех направлениях. Отсутствие специализированных отростков (дендритов и нейритов) связано с отсутствием специализированной рецепции. У гидры есть отдельные рецепторные клетки, но они не способны различать специфику разных раздражителей. Следствием этого является отсутствие четко дифференцированной ответной реакции. Кишечнополостные способны формировать реакции побега при воздействии неблагоприятных факторов среды, не дифференцируя сами факторы, а воспринимая лишь их повреждающий эффект.
3. Волну распространяющегося возбуждения сопровождает волна мышечного сокращения.
Вся дальнейшая эволюция нервной системы оказывается тесно связанной с эволюцией рецепторных и двигательных систем.
Диффузный тип нервной системы характерен и для трехслойных животных – плоских червей. Однако в связи с более сложным строением тела – появлением третьего зародышевого листка мезодермы («промежуточной кожи»), развитием двусторонней симметрии, специализацией примитивных органов чувств статоцистов (аналог органа равновесия), «глазков», обонятельных ямок, т.е. с эволюцией рецепторного аппарата, – диффузная сеть усложняется. Из нее обособляются несколько продольных стволов, расположенных вдоль тела. В переднем конце животного эти стволы соединяются поперечными перемычками. Такая решетка нервных структур получила название ортогона.Стволы ортогона принципиально отличаются от нервных стволов высокоорганизованных животных (нервов) тем, что первые на всем своем протяжении содержат как нейроны, так и их отростки, а вторые состоят исключительно из отростков, а нейроны объединяются в ганглии.
Общее направление эволюции нервного аппарата у низших червей – уменьшение числа нервных стволов и комиссур, уход нервного комплекса вглубь тела, возникновение церебрального (головного) ганглия (связанное с развитием органов чувств, в частности статоциста, органов обоняния) – привело к внешнему архитектоническому упрощению нервного аппарата. Все перечисленное максимально выражено у немертин (сколециды – низшие черви), в мозгу которых появляются скопления ассоциативных клеток наподобие высших ассоциативных центров членистых животных.
Развитие диффузной нервной системы в систему ортогонального типа определяет следующие направления эволюции нервного аппарата:
1. Централизация нервной системы.
2. Интеграцияфункций организма – интегрирующая роль нервного аппарата возрастает по мере увеличения степени его собственной централизации.
Для беспозвоночных животных характерно наличие нескольких источников происхождения нервных клеток. Для них возможно одновременное и независимое развитие изополярных нейронов из трех зародышевых листков (впрочем, происхождение нейронов из мезодермы до сих пор оспаривается, однако есть работы, в которых сообщается о развитии нервных элементов из мезодермы у ряда примитивных беспозвоночных). Считается, что такой разнообразный нейрогенез является причиной многочисленности медиаторов нервных систем.
1.2.2. Ганглионарный тип нервной системы. Появление этого типа нервной системы тесно связано с другим эволюционным нововведением – появлением сегментированных животных, пример – кольчатые черви. Тело этих животных состоит из большого числа повторяющихся сегментов, или метамеров.В каждом сегменте имеется ганглий – парное скопление нервных клеток. Именно ганглии становятся основной анатомической структурой нервной системы у большого количества таксономических групп животных. Кроме упомянутых кольчатых червей ганглионарный тип нервной системы наблюдается у двустворчатых, брюхоногих и головоногих моллюсков (рис. 1). У последних органы чувств, в частности глаза, достаточно хорошо развиты. Ганглионарный план строения нервной системы типичен и для членистоногих.
Ганглии представляют собой скопления нервных клеток, окруженные соединительнотканной капсулой. Примитивные ганглии имеют корковое строение: тела нейронов располагаются непосредственно под капсулой, направляя свои отростки внутрь нервного узла. Центральная часть ганглия, состоящая из нервных отростков и глиальных элементов, называется нейропилем. Далее отростки нейронов выходят за пределы ганглия и формируют нервы и нервные стволы: комиссуры и коннективы.Комиссурами называют нервные стволы, объединяющие парные ганглии сегментов у метамерных животных (например у кольчатых червей), или одноименные ганглии у животных в значительной степени утративших метамерность (например у моллюсков). Коннективы соединяют в продольные цепочки ганглии соседних сегментов у метамерных животных или разноименные ганглии у неметамерных.
| |
| | 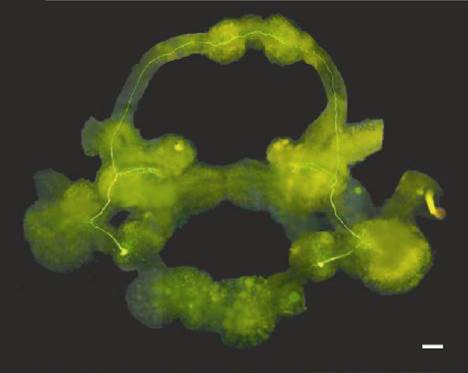 |
Рис. 1. Околопищеводное ганглионарное кольцо брюхоного моллюска Lymnaea stagnalis. Показаны топографические проекции нейронов плевральных ганглиев (По Alania M., Sakharov D.A., Elliott C.J.H., 2000).
Возникновение ганглионарного типа нервной системы связано с замечательным событием филогенеза нейронов – их поляризацией. Первыми в историческом развитии нейронов появляются клетки, которые имеют один довольно толстый отросток. В таком отростке одновременно, но изолированно могут следовать афферентные и эффекторные импульсы. Понятно, что при этом один импульс будет распространяться центробежно, а другой центростремительно. Такие нервные клетки описаны у аннелид, ракообразных, головоногих моллюсков.
Затем в эволюции возникают нейроны, единственный отросток которых дивергирует в соответствии с имеющейся поляризацией нервного импульса. В результате появляются псевдоуниполярные и биполярные нервные клетки. Нервные отростки специализируются как чувствительные, приносящее возбуждение к телу клетки – дендриты, и двигательные, передающие возбуждение от тела к эффектору – аксоны. Скорость ответной реакции в узловых сетях соответственно возрастает, возникают быстрые и генерализованные поведенческие реакции.
В ходе эволюции ганглионарного типа нервной системы отмечаются следующие основные тенденции:
1. Дальнейшая централизация и интеграция нервной системы. Эти процессы проявляются в укорочении коннектив и комиссур, слиянии ганглиев (одноименных и разноименных). Ганглии, образовавшиеся путем слияния, имеют более сложную структуру, чем их предшественники. Исчезает типичный нейропиль. Нейроны в ганглии занимают не только периферическое, но и центральное положение. Наблюдается концентрация ганглиев вокруг жизненно важных центров животного: головной конец, гонада, мышцы ноги (у двустворчатых и брюхоногих моллюсков).
2. Появление специализированных отростков нейронов. Развитие сложных воспринимающих чувствительных структур и моторных (двигательных) элементов животного потребовало более точной и адресной иннервации. С возникновением дендритов и аксонов дальнейшее функционирование нервной системы стало осуществляться по принципу рефлекса.
3. Развитие межнейрональных взаимодействий идет по двум направлениям. Первое связано с усиленной дивергенцией аксона, второе – с увеличением размера перикариона. Самые мелкие размеры тел нейронов зарегистрированы у морских ежей – 6-8 мкм. У брюхоногих моллюсков в связи с распространением полиплоидии – адаптационной эволюционной надстройки, связанной с интенсификацией функции различных систем, в том числе и нервной, – возникают гигантские нейроны. Размер перикариона таких клеток (до 1 мм) сравним разве что с мюллеровкими и маутнеровскими нейронами костистых рыб и хвостатых амфибий (животные с трубчатым типом нервной системы).
4. Цефализация. У высокоорганизованных в эволюционном отношении беспозвоночных животных (насекомые, головоногие моллюски) ганглии сливаются с образованием общей массы наподобие головного мозга позвоночных животных. Однако в таких мозгоподобных структурах нейроны остаются немиелинизированными, скорость распространения нервного импульса остается в прямопропорциональной зависимости от толщины нервного волокна. Альтернативным вариантом полиплоидии в таких сетях становится процесс слияния отдельных аксонов изофункциональных нейронов в единое гигантское нервное волокно. Академическим примером, вошедшим во все учебники по анатомии и физиологии беспозвоночных, является гигантский аксон кальмара, диаметр которого достигает 1мм – очень популярный объект электрофизиологических исследований.
1.2.3. Трубчатый тип нервной системы. У позвоночных животных в основе формирования нервной системы лежит нервная трубка, которая закладывается с дорсальной стороны эмбриона. Передний конец трубки обычно расширен и образует головной мозг. Задняя цилиндрическая часть есть не что иное, как спинной мозг. Существует гипотеза, согласно которой у предков хордовых животных имелась продольная дорсальная полоса первичного чувствительного эпителия. Затем в ходе эволюционного развития она стала погружаться под эктодерму, сформировав сначала открытый желоб, а потом и замкнутую нервную трубку. Эту гипотезу подтверждают картины раннего эмбриогенеза позвоночных животных.
В ходе эволюционного развития позвоночных трубчатая нервная система претерпевает ряд изменений.
Во-первых, все дальнейшее развитие идет по пути цефализации – преимущественного развития головного мозга. Если у примитивно устроенного хордового животного – ланцетника – головной конец практически не развит, то уже у круглоротых имеется заметное утолщение нервной трубки в головном конце. Этот примитивный головной мозг уже состоит из трех отделов: переднего, среднего и заднего. Передний отдел связан с развитием обоняния, средний – зрения и висцерального контроля (соотношение показателей гомеостаза в соответствии с меняющимися показателями внешней среды), а задний – с механорецепцией. У рыб выделяется также промежуточный мозг, достаточное развитие получает мозжечок. У амфибий значительно увеличивается передний мозг за счет развития полушарий, хорошо развит средний мозг, который у данной группы животных является высшим зрительным центром.
Во-вторых, у высокоорганизованных позвоночных возникает новый отдел головного мозга – кора больших полушарий (плащ конечного мозга). Эта структура все больше подчиняет себе рефлексы низших отделов мозга, осуществляет за ними контроль. Данный этап развития мозга обозначается как кортикализация (от лат. cortex – кора). Акуловые рыбы обладают лишь зачатком плаща конечного мозга. У рептилий появляется дифференцируемая кора головного мозга. Развитие головного мозга млекопитающих характеризуется еще большим усложнением новой коры, появляется Варолиев мост, совершенствуются структуры среднего и продолговатого мозга. У высших млекопитающих ассоциативные зоны коры являются высшим центром интегративной деятельности в ЦНС. Процесс развития новой коры сопряжен с увеличением численности нейронов. Так у крота, лишенного зрения число нейронов около 0,7 млн., у мыши – 4,2 млн., у дикой кошки более 35 млн. В процессе антропогенеза у человека разумного по сравнению с человеком умелым за счет роста плаща конечного мозга число нейронов увеличилось примерно вдвое – от 5,0·109 до 9,0·109.
Таким образом, увеличение объема и усложнение структуры отделов головного мозга позвоночных тесно связаны с развитием сенсорных систем и интегративной деятельности. Постепенно, в зависимости от притока сенсорной информации, в существующих отделах мозга появляются филогенетически новые образования, которые берут под свой контроль все большее количество функций.
Человек, как и все позвоночные животные, имеет гетерополярные нервные клетки (рис. 2). Особенно высока доля мультиполярных клеточных элементов – до 90%. Волокна таких клеток подвержены разной степени миелинизации. Это приводит к существенному увеличению скорости проведения нервных импульсов без наращивания объемов перикариона и аксона нейронов. В филогенезе появление многоотростчатых клеток связано, прежде всего, с увеличением объема движений, что, в свою очередь, сопряжено с развитием и совершенствованием мышечных систем и освоением новых жизненных пространств. Мультиполяры в ЦНС человека ответственны за образование коммуникативных связей в распределительных системах мозга и являются основой формирования сложных поведенческих актов, психической деятельности, сознания.
А Б
Рис. 2. Мультиполярные нейроны. А - дифференцировка стволовых стромальных клеток костного мозга человека в нейральном направлении (По Корочкин Л.И. и др., 2009), Б – клетки Пуркинье мозжечка крысы (По Johansson B. et al., 2008).
Следует отметить, что и у позвоночных (в том числе и высших) сохраняются черты предшествующих эволюционных типов нервных систем: ганглионарной и диффузной.
Так, в периферической нервной системе (соматической и вегетативной) чувствительные нейроны образуют ганглии (спинномозговые, симпатические и парасимпатические). При помощи ганглионарых структур в человеческом организме обеспечивается работа эволюционно древних (по сравнению с психикой) структур. Это, прежде всего, афферентация (чувствительность, рецепция) и автономная деятельность внутренних органов.
Также в нашем организме сохранились и признаки диффузной нервной системы. Она образует третий отдел вегетативной нервной системы – метасимпатическую нервную систему (в дополнение к симпатической и парасимпатической). Данный отдел обеспечивает автономную работу полых внутренних органов. Нейроны метасимпатической нервной системы образуют микроганглионарные скопления внутри стенок органов, координируя их деятельность (например, перистальтические движения кишечника, обеспечивающие продвижение пищи). Процессы, протекающие в диффузной нервной системе, могут видоизменяться под влиянием симпатической и парасимпатической нервных систем.








