Классическим объектом эмбриональных исследований является ланцетник – представитель класса хордовых.
Яйцеклетка ланцетника олигоизолецитальная, ядро располагается ближе к анимальному полюсу. Оплодотворение наружное. Место проникновения сперматозоида в яйцеклетку определяет вентральную сторону будущего зародыша. На дорсальной стороне зиготы возникает серый серп, что связано с перераспределением содержимого цитоплазмы в яйце. При этом белково-липидные включения скапливаются у одного из краев зиготы, определяя вегетативный полюс зародыша.
Дробление зиготы ланцетника полное равномерное и синхронное. При этом образуется однослойная целобластула с полостью, заполненной жидкостью – бластоцелем.
Начиная со стадии восьми бластомеров появляются неравномерные по величине клетки – более мелкие на анимальном полюсе и более крупные на вегетативном. К концу дробления сохраняется полярность бластулы, в которой определяется дно – вегетативный полюс и крыша – анимальный полюс.
Начало гаструляции, протекающей путем инвагинации, отмечается формированием впячивания на вегетативном полюсе бластулы внутрь бластоцеля.
В результате инвагинации образуется двухслойная чаша, в которой наружный слой представлен эктодермой, а внутренний – энтодермой. При этом формируется новая полость между ними – гастроцель (первичная кишка), а бластоцель принимает форму щели. Гастроцель сообщается со внешней средой посредством бластопора или первичного рта (рис. 6). В бластопоре различают дорсальную, вентральную и две латеральные губы. Бластопор является важнейшим ориентиром в ходе гаструляции, поскольку одни и те же его участки у различных видов позвоночных формируют одинаковые структуры. Каждая губа занимает положение, соответствующее будущей спинной, брюшной и боковым частям зародыша, и имеет в своем составе по два зародышевых листка. В ходе поздней гаструляции формируется третий зародышевый листок – мезодерма и осевой зачаток – хорда. Источником развития хорды являются клетки из состава энтодермы, мигрирующие через область дорсальной губы бластопора. Мезодерма образуется путем выселения клеток через область латеральных губ, оставшиеся клетки являются составной частью энтодермы. Образовавшаяся хорда индуцирует процесс нейруляции и формирование нервной трубки из материала дорсальной эктодермы. Таким образом, в ходе гаструляции у ланцетника формируется три зародышевых листка – мезодерма, эктодерма, энтодерма и комплекс осевых зачатков – хорда, нервная и кишечная трубки.

Рис. 6. Поперечные разрезы зародышей ланцетника [7].
А – карта презумптивных зачатков яйцеклетки ланцетника; Б – поперечный срез бластулы ланцетника; В, Г – поздняя гаструла (продольный и поперечный разрезы); Д – превращение нервной пластинки, погрузившейся под кожную эктодерму, в нервный желобок и начало обособления мезодермальных карманов от первичной кишки; Е – обособление замкнутых целомических мешков, замыкание энтодермы в кишечную трубку и формирование хордального тяжа. Б; 1 – крыша бластулы, 2 – дно бластулы, 3 – краевая зона бластулы; В: 4 – дорсальная губа бластопора, 5 – мелкоклеточный материал вентрального серпа
В последующем происходит разделение мезодермы на спинные сегменты – сомиты, число которых у ланцетника достигает 60–65 пар. Часть латерально расположенной мезодермы не сегментируется и расщепляется на наружный (париетальный) и внутренний (висцеральный) листки спланхнотома. В последующем по мере роста обоих листков между экто- и энтодермой формируется вторичная полость – целом. На переднем конце зародыша возникает углубление – ротовая бухта, растущая по направлению к переднему отделу кишечной трубки. При контакте эктодермы ротовой бухты и слепого конца кишечной трубки под действием процессов апоптоза клеток возникает сообщение кишки с внешней средой. Аналогичные процессы, протекающие на заднем конце зародыша, формируют анальное отверстие. По бокам головного отдела зародыша в результате прорыва контактирующих экто- и энтодермы полость передней кишки сообщается с внешней средой путем сформировавшегося жаберного аппарата.
Таким образом, на примере развития ланцетника можно наблюдать закономерное перемещение и выселение материала будущих эмбриональных зачатков тканей с формированием зародышевых листков и в последующем – осевого комплекса.
Развитие амфибий. Яйцеклетка амфибии мезотелолецитальная, имеет ядро, расположенное на анимальном полюсе, и умеренное количество желтка – на вегетативном. Оплодотворение внутреннее (хвостатые амфибии) или наружное (бесхвостые). Сперматозоид проникает в яйцеклетку в анимальной части. В ходе перераспределения материала цитоплазмы на стороне, противоположной проникновению сперматозоида, возникает область серого серпа. Дробление у амфибий голобластическое, неравномерное. Первая полоска дробления, возникшая на анимальном полюсе, пересекает серый серп и замыкается на вегетативном полюсе. Последующие борозды дробления проходят ближе к вегетативному полюсу, поэтому бластомеры вегетативного полюса значительно крупнее бластомеров анимального.
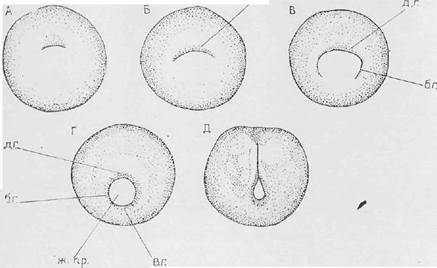
Рис. 7. Последовательные стадии (А—Д) гаструляции у амфибий, внешний вид: б.г – боковые губы; в.г – вентральная губа; д.г — дорсальная губа бластопора; ж.пр. — желточная пробка [8].
В результате дробления возникает многослойная бластула – амфибластула. Более мелкие бластомеры располагаются в несколько слоев в области крыши, крупные формируют дно бластулы. Серый серп намечает место будущей дорсальной губы бластопора, образование которой происходит в начале гаструляции в ходе перемещения клеток внутрь зародыша. Гаструляция у амфибий осуществляется путем эпиболии – обрастания вегетативного полюса подвижными мелкими клетками анимального полюса. В процессе миграции бластомеров формируется дорсальная губа бластопора, которая распространяется билатерально и замыкается в брюшной части зародыша. Область формирования бластопора является важным морфологическим маркером процесса гаструляции амфибий, где происходят наиболее активные и закономерные клеточные перемещения (рис. 7). Клетки, мигрирующие через дорсальную губу бластопора, формируют прехордальную пластинку и хорду. Затем мигрирует материал хорды, который располагается по центральной части зародыша под будущей нервной трубкой. Клетки, прошедшие через область латеральных губ, образуют туловищную мезодерму, располагающуюся по бокам от хорды (рис. 8).
Таким образом, в процессе гаструляции у амфибий клетки крыши бластулы подворачиваются в области бластопора, мигрируют внутрь зародыша и формируют его многослойное строение. В конце гаструляции бластопор постепенно уменьшается, и от него остается небольшое углубление – проктодеум – место, где в последующем сформируется клоакальное отверстие.

Рис. 8. Схема перемещений клеток, происходящих при гаструляции у амфибий [6]:
А, Б, В – последовательные стадии перемещения клеток в ходе гаструляции: 7 – эктодерма; 2 – материал будущей мезодермы; 3 – богатые желтком клетки энтодермы; 4 – клетки в составе эктодермы, образующие презумптивную нервную пластинку; 5 – бластопор; 6 – материал хорды; 7 – энтодерма; 8 – кожная эктодерма; 9 – бластоцель; 10 – гастроцель, или первичная кишка; 11 – материал нейроэктодермы; 12 – дорсальная губа бластопора; 13 – вентральная губа бластопора; 14 – материал мезодермы
Процесс нейруляции протекает аналогично нейруляции у ланцетника. Сформировавшаяся нервная трубка погружается внутрь зародыша и располагается между хордой и кожной эктодермой. Под индуцирующим влиянием хорды одновременно происходит образование сомитов, которые подразделяются на дорсальные и вентральные. Так возникает комплекс эмбриональных зачатков и тканей. Дальнейшее развитие зародыша заключается в формировании тканей и органов.
Развитие рыб. Полителолецитальная яйцеклетка рыб содержит много желтка, поэтому дробление зиготы меробластическое, неравномерное, дискоидальное. Процесс дробления происходит только на анимальном полюсе зиготы. В ходе дробления образуется многослойная дискобластула (рис. 9). Бластоцель представляет собой небольшую полость между бластодиском и желтком. Процесс гаструляции протекает путем деламинации – расщепления бластулы на два листка – наружный эпибласт и внутренний гипобласт. При этом не происходит образование типичного бластопора. В дальнейшем наблюдается инвагинация края эпибласта и распространение клеток будущей хорды и мезодермы между эпибластом и гипобластом. Таким образом, формируется трехслойный зародыш. В ходе гаструляции у рыб происходит формирование первого внезародышевого органа – желточного мешка. Этому предшествует обособление тела зародыша от внезародышевой части бластодиска за счет образования так называемой туловищной складки – кольцеобразного перехвата.

Рис. 9. Дробление костистых рыб [5].
Клетки внезародышевого материала всех трех зародышевых листков образуют желток и формируют желточный мешок – внезародышевый орган, обеспечивающий дальнейшее развитие зародыша. В эпидермальном слое желточного мешка возникают клетки, расщепляющие желток на составные элементы, которые идут на питание зародыша. В мезодермальном слое появляются кровеносные сосуды, по которым питательные вещества поступают к зародышу. Наружный эктодермальный слой клеток желточного мешка осуществляет газообмен. Желточный мешок соединяется с полостью средней кишки при помощи желточного стебелька. По мере расходования питательного материала желтка мешок втягивается в тело зародыша, энтодерма становится частью кишечной стенки, эктодерма входит в состав кожных покровов, а сосудистая система желточного мешка редуцируется. Таким образом, желточный мешок является провизорным (временным) органом, появляющимся в эволюции позвоночных.
Развитие птиц. К началу гаструляции зародыш птицы представляет собой диск, который лежит на поверхности желтка. Ранняя гаструляция происходит путем деламинации с образованием двух листков – эпибласта, лежащего на поверхности и гипобласта, прилегающего к желтку. Между ними формируется щелевидная бластоцель, заполненная жидкостью.
Основным механизмом поздней гаструляции является миграция клеток центральной части эпибласта – зародышевого щитка (рис. 10). Бластомеры располагающиеся на периферии зародышевого щитка, двумя потоками перемещаются на один из полюсов, который в ходе дальнейшего развития будет задним концом зародыша. Встретившись у полюса, клеточные потоки движутся по средней линии к центру и формируют утолщение – первичную полоску, которая намечает кранеокаудальную ось зародыша. На конце полости формируется первичный узелок – аналог дорсальной губы бластопора. Далее идет миграция клеток эпибласта в бластоцель через первичную полоску и первичный узелок. Первыми мигрируют клетки через первичный узелок вниз к гипобласту, замещают его клетки и образуют прехордальную пластинку или переднюю кишку. Вторам потоком клетки двигаются через первичный узелок в бластоцель, располагаются под эпибластом и формируют хорду. Следующим этапом в области первичной полоски клетки проваливаются под эпибласт и, формируя первичную бороздку, делятся на два потока: одни замещают клетки гипобласта и формируют зародышевую энтодерму, другие располагаются между эпибластом и гипобластом и образуют третий зародышевый листок – мезодерму. Оставшиеся клетки эпибласта формируют зародышевую эктодерму и нервную пластинку. Затем зародышевый щиток приподнимается над массой желтка, его края подгибаются, клетки эктодермы замыкаются по средней линии и формируют кишечную трубку.

Рис. 10. Гаструляция у птиц [7] и млекопитающих [1].
Развитие млекопитающих
У высших (плацентарных) млекопитающих яйцеклетка содержит малое количество желтка и является вторичной олигоизолецитальной. Дробление полное, неравномерное, асинхронное. В ходе дробления наблюдается ранняя компактизация бластомеров, в результате чего образуется плотный клеточный шар – морула (рис. 11). В последующем возникает полость наполненная жидкостью – бластоцель и формируется бластоциста. В процессе образования бластоцисты бластомеры наружной клеточной массы формируют трофобласт, а бластомеры внутренней клеточной массы – эмбриобласт.
У плацентарных млекопитающих по сравнению с беспозвоночными и низшими позвоночными очень рано начинает функционировать геном зародыша. Трансляция белков с генома зародыша наблюдается со стадии восьми бластомеров.
В период дробления у млекопитающих происходит ранее обособление клеток, которые образуют внезародышевые органы, что связано с внутриутробным способом развития.

Рис. 11. Схема дробления и образования бластоцисты млекопитающих [7]: А – полное асинхронное дробление зиготы, стадии 2 и 4 бластомеров; Б – обрастание светлыми клетками темных (морула); В – образование бластоцисты, формирование полости, обособление темных клеток внутренней клеточной массы от светлых - наружной клеточной массы - трофобласта; 1 – внутренняя клеточная масса, 2 – трофобласт
Особенности развития млекопитающих:
1) в ходе развития у млекопитающих появляется необходимость в создании таких внезародышевых органов как амнион и плацента, которые защищают зародыш и обеспечивают его метаболитами матери;
2) развитие зародыша происходит в матке, что освобождает его от необходимости развиваться быстро;
3) внезародышевые органы, в частности плацента, обеспечивают зародыша питательными веществами материнского организма, поэтому яйцеклетка не нуждается в большом количестве желтка и дробление протекает с той же скоростью что и деление соматической клетки.
Гаструляция млекопитающих, как и у всех позвоночных, протекает в две фазы и заканчивается образованием трех зародышевых листков, комплекса осевых зачатков органов и внезародышевых органов.
Первая фаза гаструляции осуществляется путем деламинации и протекает одновременно с имплантацией. При этом эмбриобласт бластоцисты расщепляется на эпи- и гипобласт (рис. 10). Эпибласт является источником развития зародыша и внезародышевой амниотической эктодермы. Клетки гипобласта перемещаются по внутренней поверхности трофобласта и участвуют в образовании желточного мешка, который плотно прилегает к трофобласту. Одновременно в конце ранней гаструляции происходит формирование таких внезародышевых органов, как желточный мешок, хорион, аллантоис.
Первыми из внезародышевых органов формируются желточный мешок и аллантоис. Сначала под эпибласт выселяются клетки, которые заполняют всю полость бластоцисты и образуют первичную внезародышевую мезенхиму. В ней формируются две полости – одна над эпибластом, вторая под гипобластом. Сформировавшиеся полости обрастают клетками за счет размножения клеток обоих клеточных пластов – эпи- и гипобласта. При этом образуются два пузыря – амниотический, расположенный над будущим зародышем, и желточный – под гипобластом; материалом для пузырей станут это будущие амнион и желточный мешок. Часть мезенхимы подходит близко к трофобласту и вместе с ним формирует хорион.
К началу ранней гаструляции зародыш имеет следующее строение: снаружи находится хорион, состоящий из двух слоев – трофобласта и внезародышевой мезенхимы. Сам трофобласт разделен на два листка – симпластотрофобласт, располагающийся снаружи, и внутренний цитотрофобласт. Полость зародыша заполнена внезародышевой мезенхимой. В ней находится два пузырька – амниотический пузырек, состоящий из внезародышевой мезенхимы и внезародышевой эктодермы; и пузырек желточного мешка, сформированный внезародышевой мезенхимой и внезародышевой энтодермой. Пузырьки прилегают друг к другу и прикреплены к хориону с помощью амниотической ножки, которая образована внезародышевой мезенхимой.
Тело зародыша, образованное клетками дна амниона и крышей желточного мешка, называется зародышевым диском и состоит из эпи- и гипобласта.
Поздняя гаструляция у млекопитающих осуществляется путем миграции и частичной инвагинации (рис. 10). Основные процессы протекают в эпибласте и напоминают таковые при гаструляции у птиц с образованием первичной полоски и первичного узелка. Первичная полоска содержит материал для формирования зародышевой эктодермы, и мезодермы. Первичный узелок является источником хордального отростка. Клетки первичной полоски прорывают эпибласт, мигрируют между эпибластом и гипобластом и образуют мезодерму. С этого момента зародыш становится трехслойным.
Клетки первичного узелка мигрируют под эпибласт и образуют хордальный отросток. Часть клеток первичной полоски перемещаются по направлению к гипобласту, встраиваются в него и занимают центральное положение. Из них формируется кишечная энтодерма.
Из всех зародышевых листков (но в основном из мезодермы) выселяются клетки, которые заполняют все пространство между зародышевым листком и формируют вторичную, или зародышевую, мезенхиму.
Таким образом, ранняя гаструляция у млекопитающих, как и у всех позвоночных, заканчивается расщеплением эмбриобласта на два листка, а поздняя – образованием третьего зародышевого листка – мезодермы, комплекса осевых зачатков (хорды, нервной и кишечной трубки) и внезародышевых органов (амниона, хориона, желточного мешка).
Внезародышевые органы – временные, или провизорные, – образуются у позвоночных в процессе гаструляции и имеют характерные особенности:
- формируются вне тела зародыша;
- принимают активное участие в процессе роста и развития;
- перестают функционировать при рождении.
Впервые внезародышевые органы в процессе эволюции появились у рыб в виде желточного мешка, который выполняет трофическую функцию у всех позвоночных, кроме млекопитающих. В стенке желточного мешка формируются кровеносные сосуды, которые связывают его с сосудистой системой зародыша: по ним желток, расщепленный ферментами, поступает в организм зародыша.

Рис. 12. Схема развития внезародышевых органов у птиц [7]: 1 – скорлупа; 2 – воздушная камера; 3 – белочная оболочка (белок); 4 – амниотическая складка; 5 – зародыш; 6 – кишечная трубка; 7 – аллантоис; 8 – формирование серозной оболочки (хориона); 9 – обрастание желтка и образование желточного мешка
С выходом на сушу у позвоночных появляется необходимость еще в одном внезародышевом органе – амнионе, который дает возможность зародышу развиваться в водной среде. Необходимость водной среды для развития зародыша обусловлена тем, что она удерживает постоянную температуру окружающей среды, предохраняет зародыш от высыхания, механических повреждений, обеспечивает процессы выделения и поглощения.
Амнион (амниотическая оболочка) формируется двумя способами: при замыкании надзародышевой складки у птиц и рептилий (рис. 12) или путем раздвижения экто- и энтодермы у млекопитающих и человека (рис. 13). Независимо от способа образования зародыш оказывается окруженным амниотическими оболочками со всех сторон. Сами оболочки продуцируют жидкость, которая заполняет амниотическую полость. Химический состав амниотической жидкости изменяется по мере развития зародыша.
Функции амниотической жидкости:
- механическая защита (от повреждения, высыхания, срастания);
- выбор пути дифференцировки клеток наружной поверхности зародыша и создания необходимого давления для развития полостей – носовой, ротовой, ткани для развития легкого.
В ходе эволюции формируется орган или группа вспомогательных органов, которые обеспечивают дыхание и выделение продуктов метаболизма. Это серозная оболочка у низших и хорион, плацента и аллантоис у млекопитающих.
Серозная оболочка является самой наружной внезародышевой оболочкой, которая примыкает к скорлупе у птиц или стенке матки у млекопитающего (хорион). Эта оболочка осуществляет обмен веществ между зародышем и окружающей средой.
Аллантоис – развивается в результате выпячивания стенки задней кишки. У птиц и рептилий он выполняет дыхательную и выделительную функции. У млекопитающих проводит сосуды к зародышу и плаценте.
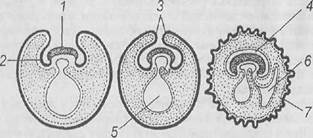
Рис. 13. Схема последовательных стадий образования внезародышевых органов у млекопитающих [7]: 1 – зародыш; 2 – туловищная складка; 3 – амниотические складки; 4 – амнион; 5 – желточный мешок; 6 – аллантоис; 7 – хорион
Хорион формируется с момента исчезновения оболочки оплодотворения и внедрения трофобласта в стенку матки, то есть с момента имплантации. В том месте, где зародыш прикрепился к слизистой оболочке матки, ворсины хориона разрастаются и вместе с децидуальной оболочкой матки формируют плаценту. На остальном протяжении они редуцируют, образуя гладкий хорион.
Первоначально ворсины представляют собой выросты трофобласта и называются первичными ворсинами. Ворсины, сформировавшиеся из трофобласта и подлежащей внезародышевой мезодерме, называются вторичными. Третичные, или истинные, ворсины состоят из эпителия трофобласта, соединительной ткани и сосудов зародыша, которые прорастают вдоль аллантоиса и разветвляются в ворсины.
Сосуды ворсин связаны с сосудами зародыша через аллантоисные сосуды, которые входят в состав пупочного канатика.
Обмен метаболитами в плаценте у млекопитающих осуществляется следующим образом: метаболиты зародыша по пупочным артериям попадают в сосуды ворсин и проникают последовательно через стенку капилляра ворсины, соединительную ткань ворсины, эпителий трофобласта и поступают в материнский организм. От матери все необходимые метаболиты проходят тот же плацентарный барьер, но уже в обратном порядке и по пупочной вене поступают к зародышу.
В зависимости от типа питания зародыша у млекопитающих выделяют следующие типы плацент (рис. 14).
1. Эпителиохориальная – ворсины хориона врастают в ткани матки и эпителий ворсин контактирует с эпителием матки.
2. Десмохориальная – ворсины хориона разрушают эпителий маточных желез и контактируют с соединительной тканью матки.
3. Эндотелиохориальная – ворсины хориона разрушают эпителий и соединительную ткань и контактируют с эндометрием сосудов.
4. Гемохориальная – ворсины хориона разрушают стенку кровеносного сосуда матки и контактируют непосредственно с кровью материнского организма.

Рис. 14. Типы плацент [2].
Животные с первым и вторым типом плацент после рождения быстро приспосабливаются к самостоятельной жизни, так как хорион только поглощает из крови матери белки и расщепляет их, а синтез собственных белков происходит в печени зародыша.
При третьем и четвертом типах плацент хорион сам синтезирует эмбриоспецифические белки из аминокислот материнского организма. Вот почему зародыши животных и человека с таким типом плацент длительное время не способны к внеутробному существованию.






